宰后成熟期间结构蛋白对秦川牛肉嫩度的影响
2022-09-30马旭华李亚蕾罗瑞明张杏亚
马旭华,杨 波,李亚蕾,罗瑞明,张杏亚,张 萌
(宁夏大学食品与葡萄酒学院,宁夏 银川 750021)
作为一种低脂肪、高蛋白的食品,牛肉深受消费者喜爱。其中,嫩度是影响消费者满意度和接受度的主要因素,也是衡量牛肉食用品质的关键指标之一。因此,国内外对肉嫩度变化机理的研究一直没有停止。综合学者的研究成果,可总结出影响嫩度形成的4 个主要机理:一是肌原纤维蛋白的碎裂或蛋白质结构发生改变导致Z线弱化;二是通过蛋白酶(钙蛋白酶、组织蛋白酶)和蛋白酶体裂解蛋白质引发嫩度变化;三是糖酵解酶在宰后调节嫩度;四是细胞凋亡和热休克蛋白参与嫩度形成。
蛋白质组学是指研究一个组织或细胞的所有蛋白质。蛋白质组学起初是用于人体医学检测,随着技术的发展,逐渐应用到食品学科中,为研究食品深层变化机理提供了新的方法。此技术不用同位素标记,通过液相色谱-质谱联用,分析蛋白质酶解后肽段的信号强度,进而对蛋白质定性定量。在3D蛋白质组学技术中主要从3 个维度(离子强度、质荷比和保留时间)检测蛋白质,4D-非标记定量(4D-label free quantification,4D-LFQ)蛋白质组学技术则是在3D分离技术基础上增加了参数离子淌度,其主要根据离子的截面及形状进行分离,为测定秦川牛肉组织这种复杂体系样本带来了更多的可能。李升升利用同位素相对标记和绝对定量(isobaric tags for relative and absolute quantitation,iTRAQ)技术研究牦牛平滑肌贮藏过程嫩度的变化机理,结果表明ADP/ATP转位酶1、钙蛋白酶抑素、原肌球蛋白-3链、细胞色素b-c1复合物亚基1等蛋白可能是牦牛平滑肌贮藏期嫩度变化的指示蛋白。Sawdy等研究表明牛背长肌贮藏7 d后嫩度的变化与肌球蛋白重链断裂有关,结果显示,在肌肉嫩化过程中,肌球蛋白轻链1、2和重链1的表达量明显降低。Alessandro等研究发现在肉成熟过程中肌动蛋白显著降解,产生了31 kDa条带。
本研究主要采用4D-LFQ蛋白质组学技术筛选差异蛋白质,在贮藏0~8 d对秦川牛背最长肌的蛋白质组分进行检测,分析其涉及的信号通路,探索秦川牛宰后结构蛋白降解对嫩化机制的影响,以更加深入地研究剪切力、肌原纤维小片化指数(myofibrillar fragmentation index,MFI)、肌肉总可溶性蛋白、肌原纤维蛋白、肌浆蛋白的变化机理。实验选用秦川黄牛背最长肌作为研究对象,秦川黄牛作为陕甘宁地区优势特色畜种,肉质细腻、瘦肉比例高、大理石纹路明显,其背最长肌可做成高档牛排。利用4D-LFQ蛋白质组学技术研究其嫩度变化机理,有望为秦川牛肉工业加工中对嫩度的控制提供相应的理论支撑。
1 材料与方法
1.1 材料与试剂
25 月龄秦川公牛由甘肃省平凉市泾川县旭康食品有限责任公司提供。
三乙基碳酸氢铵、尿素、碘代乙酰胺、二硫苏糖醇美国Sigma公司;三氟乙酸 美国Sigma-Aldrich公司;胰酶 美国Promega公司;蛋白酶抑制剂 德国Calbiochem公司;乙腈 美国Fisher Chemical公司;甲酸 瑞士Fluka公司;聚氰基丙烯酸正丁酯(bicinchoninic acid,BCA)试剂盒 上海碧云天公司;2X还原性Losding Buffer、考马斯亮蓝G-250、考马斯亮蓝R-250、牛血清白蛋白 中国上海麦克林生化科技有限公司。
1.2 仪器与设备
NAI-CS150超声波细胞破碎机 上海那艾精密仪器有限公司;205便捷式pH计 德国德图集团;MiniVac Alpha冷冻离心机 美国Scan Speed公司;timsTOF Pro质谱仪 德国Bruker公司;Forma 994-80 ℃超低温冰箱 美国Thermo公司;TA-XT plus质构仪 英国Stable Micro Systems公司;Power Pac基础电泳仪 美国伯乐公司;c150凝胶成像仪 美国Azure Biosystems公司;JXFSTPRP-CL冷冻研磨仪 杭州优嘉科学仪器有限公司;UV-1200紫外分光光度计 上海美谱达仪器有限公司。
1.3 方法
1.3.1 样品采集
采样对象选用3 头自然放养、发育正常、健康无病、体质量相近、25 月龄左右的秦川公牛。用体积分数75%乙醇溶液对采样所需刀具、镊子等工具进行消杀处理。屠宰后的秦川公牛对胴体修整后,用去离子水冲洗,取3 头牛的左胴体背最长肌,每个背最长肌样品平均分割成3 份,共计9 个样品。样品分割成块后用透气性为23.5 g/(m·d)的聚乙烯薄膜密封,贮存于0~4 ℃的冰箱中。待贮存时间为0、2、4、6、8 d时立即置入液氮中2 h,然后放入-80 ℃低温冰箱中保存,待检测分析使用。
1.3.2 秦川牛背最长肌剪切力测定
参考李桂霞等的方法稍作修改。取规格约3 cm×6 cm×6 cm肉块除去表面的脂肪和筋膜,将肉样置于蒸煮袋内,抽去袋内空气,使肉块表面紧贴蒸煮袋,肉块中插入热电偶温度计,密封袋口,置于水浴锅在80 ℃下加热至中心温度为70 ℃后立即取出,冷却至室温,将肉样沿肌纤维方向用直径1.27 cm采样器平行取4 个肉柱,然后用质构仪V型活动剪切刀架垂直于肌纤维方向测定其剪切力,剪切速率1.5 mm/s,剪切距离40 mm,测定4 次取平均值。
1.3.3 秦川牛背最长肌蛋白质溶解度测定
选取宰后贮藏0、2、4、6、8 d的秦川牛背最长肌样品,参考Joo等的方法提取总可溶性蛋白和肌浆蛋白,参考李升升的方法提取肌原纤维蛋白,并采用考马斯亮蓝法测定蛋白质含量,即为溶解度。
参考焦洁、Bradford等的方法,以牛血清白蛋白作标准曲线,595 nm波长处测其吸光度,通过Origin软件线性拟合可得方程=5.767 6-0.018 2(=0.996 1)(为吸光度;为蛋白质含量/(mg/g))。
1.3.4 十二烷基硫酸钠-聚丙烯酰氨凝胶电泳分析
根据黄峰的方法稍作修改。制备5%(质量分数,下同)的浓缩胶和12%的分离胶,将1.3.3节所得总可溶性蛋白溶液统一调至质量浓度1.00 mg/mL,加入7.5 μL样品缓冲液(2×还原性Loading Buffer)和15 μL蛋白质溶液样品于小离心管中混匀,煮沸加热5 min,室温冷却后加入到上样孔中。加入10 μL蛋白Marker(10~250 kDa)。浓缩胶电流设置为15 mA,待溴酚蓝指示条带迁移至分离胶与浓缩胶分割线处调整电流为25 mA,蛋白Marker不同条带均分离后关闭电源,电泳完成。小心取出凝胶置于培养皿中,加入考马斯亮蓝R-250,50 r/min摇床染色1 h后脱色,脱色剂更换3~5 次,每30 min更换1 次。脱色完成后置于凝胶成像仪中成像。
1.3.5 MFI测定
参考Culler等的方法,采用考马斯亮蓝法测蛋白质量浓度。用MFI缓冲液将蛋白溶液稀释至质量浓度0.5 mg/mL,在595 nm波长处测定吸光度,每个样品重复3 次,取平均值带入下式计算MFI。
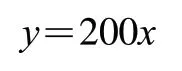
式中:为MFI;为。
1.3.6 4D-LFQ蛋白质组学技术检测蛋白质含量
参照张杏亚等的方法提取蛋白质并测定蛋白质含量,参照罗辉等的方法进行数据库搜索、生物信息学分析筛选差异蛋白质,利用京都基因与基因组百科全书(Kyoto encyclopedia of genes and genome,KEGG)数据库(http://www.genome.jp/kegg/)检索代谢途径,并利用String 10.0(http://string-db.org/)和Cytoscape 3.6.1软件绘制差异蛋相互作用网络图。
1.4 数据处理与分析
数据均以平均值±标准差表示,运用SPSS 24软件进行单因素方差分析和Duncan检验,确定数据间是否具有差异显著性,<0.05为差异显著,<0.01为差异极显著,利用Origin 8.0软件作图。
2 结果与分析
2.1 不同贮藏时间秦川牛背最长肌剪切力的变化
肉的嫩度直接影响其食用时的口感,故嫩度是牛肉的重要品质指标之一。在研究中常用剪切力来反映肉的嫩度情况。
由图1可知,0~8 d贮藏期剪切力总体呈先上升后下降的变化趋势,且上升的幅度小于下降的幅度。牛肉在贮藏0 d时的剪切力为(120.25±5.15)N,0~4 d时呈现上升趋势,4 d时最高,剪切力为(157.94±2.53)N;4 d后呈现下降趋势,8 d时达到整个贮藏期的最低点,剪切力为(93.41±5.08)N。马秀利在研究牛肉品质变化时剪切力也出现相似的变化趋势。
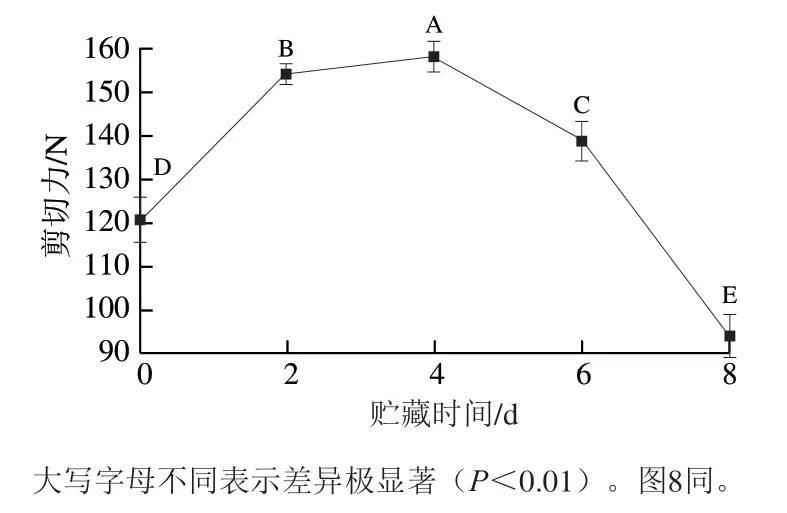
图1 秦川牛背最长肌不同贮藏时间剪切力变化Fig. 1 Changes in shear force of Qinchuan cattle Longissimus dorsi muscle at different storage times
2.2 不同贮藏时间秦川牛背最长肌总可溶性蛋白含量的变化
由图2可知,0~8 d贮藏期总可溶性蛋白含量呈显著下降趋势(<0.05)。贮藏0 d时含量为(196.44±2.10)mg/g,贮藏8 d时总可溶性蛋白含量达到整个过程的最低点((128.47±0.87)mg/g),总体下降了34.60%。这一结果表明秦川牛肉贮藏期蛋白质变化较为明显,宰后贮藏过程中肌肉组织进行代谢,氨基酸侧链基团的氧化修饰程度高导致蛋白质构象的变化加剧,从而使蛋白质分子内暴露更多的疏水基团,表面疏水性不断增加。同时由于游离巯基易氧化转化为二硫键,导致蛋白质大量聚集和沉淀,使肌肉组织蛋白发生降解。
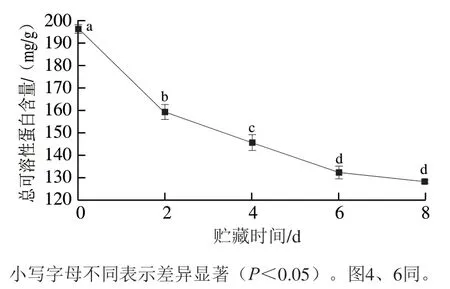
图2 秦川牛背最长肌贮藏过程中总可溶性蛋白的含量变化Fig. 2 Changes in total soluble protein content of Longissimus dorsi muscle of Qinchuan cattle during storage
根据秦川牛背最长肌蛋白分子质量选择蛋白Marker指示范围(10~250 kDa),共有10 条条带。电泳前在每个凹糟中加入等质量浓度的溶液,保证有且仅有蛋白降解变化影响条带的颜色和宽度。
由图3可知,秦川牛背最长肌条带众多,由此可判断蛋白种类较多。随贮藏时间延长,150~250 kDa内蛋白条带逐渐变窄;秦川牛背最长肌总可溶性蛋白100 kDa处I条带逐渐变浅;37~50 kDa内蛋白条带逐渐变浅且分散;25~37 kDa内原肌球蛋白条带逐渐变浅,但II条带显著变深;10~25 kDa内III、IV条带变深,IV条带明显变深。总体上,高分子质量蛋白条带逐渐变浅,低分子质量蛋白条带逐渐变深,这一结果与总可溶性蛋白分析结果结合,可以推测出秦川牛背最长肌总可溶性蛋白出现显著降解现象,其结构总可溶性蛋白变化可能会影响嫩度的形成。
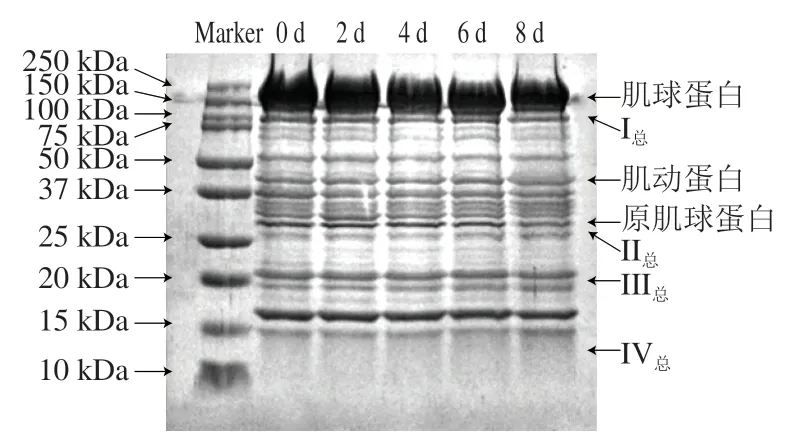
图3 秦川牛背最长肌贮藏过程总可溶性蛋白电泳图Fig. 3 Degradation of total soluble proteins in Qinchuan cattle Longissimus dorsi muscle during storage
2.3 不同贮藏时间秦川牛背最长肌肌浆蛋白含量的变化
利用肌浆蛋白在水溶液和低盐溶液中具有较高的溶解性提取肌浆蛋白,以此判断肌浆蛋白含量的变化。由图4可知,0~8 d贮藏期内肌浆蛋白含量呈上升趋势,由(43.48±0.43)mg/g增加至(52.97±0.35)mg/g,总体增加了21.83%。
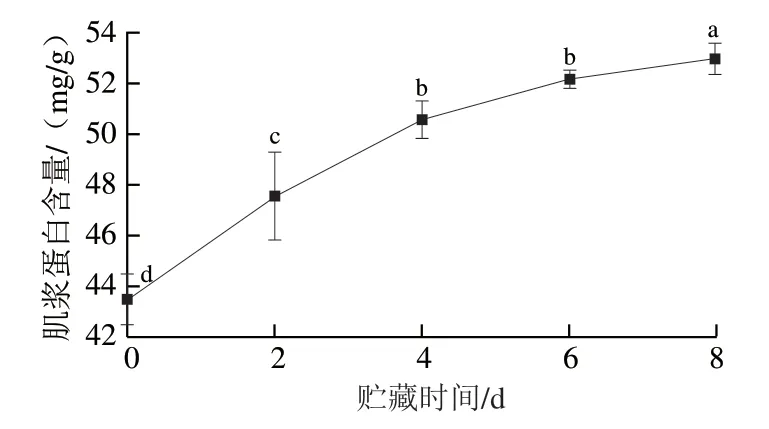
图4 秦川牛背最长肌贮藏过程中肌浆蛋白含量的变化Fig. 4 Changes in sarcoplasmic protein content in Qinchuan cattle Longissimus dorsi muscle during storage
由图5可知,0~8 d贮藏期I(150~250 kDa范围内两条蛋白条带)逐渐变浅;II(100 kDa蛋白条带)逐渐变浅变窄;III(37 kDa蛋白条带)由3 条蛋白条带逐渐变成两条,中间条带消失;IV(25 kDa蛋白条带)在前2 d逐渐明显加深,2~8 d贮藏期内变化不明显;V(15 kDa蛋白条带)在2 d后逐渐变宽,可能是大分子蛋白分解的结果;其他条带无显著变化。通过以上5 处条带的变化也可归纳出,高分子质量蛋白条带逐渐变浅,低分子质量蛋白条带逐渐变深变宽。可以推测出,贮藏过程中秦川牛背最长肌肌浆蛋白中高分子质量蛋白出现分解,含量变少,低分子质量蛋白种类逐渐增多,浓度不断增大。
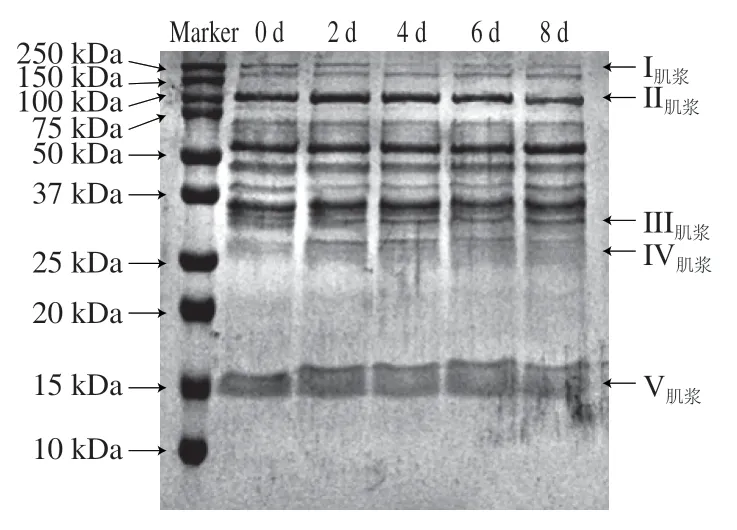
图5 秦川牛背最长肌贮藏过程中肌浆蛋白降解电泳图Fig. 5 Degradation of sarcoplasmic protein in Qinchuan cattle Longissimus dorsi muscle during storage
2.4 不同贮藏时间秦川牛背最长肌肌原纤维蛋白含量的变化
利用肌原纤维蛋白具有高盐溶解性提取肌原纤维蛋白,观察肌原纤维蛋白的变化情况。由图6可知,0~8 d贮藏期肌原纤维蛋白含量呈下降趋势,由(136.72±3.06)mg/g持续下降至(67.60±0.84)mg/g。前期下降速率较快,后期下降速率有所放缓,总体下降了50.56%。
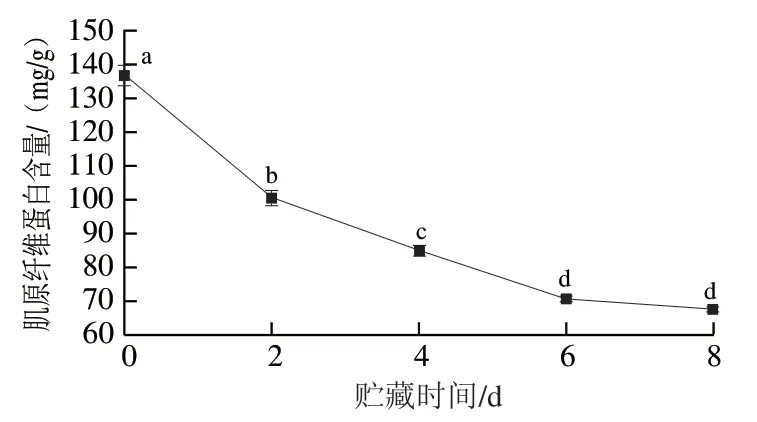
图6 秦川牛背最长肌贮藏过程中肌原纤维蛋白含量的变化Fig. 6 Changes in myofibrillar protein content in Qinchuan cattle Longissimus doris muscle during storage
由图7可知,随贮藏时间延长,分子质量在100~250 kDa范围内蛋白条带逐渐变深,蛋白含量逐渐增多;分子质量在37~50 kDa内的蛋白条带变浅;分子质量在25~37 kDa内出现相对高分子质量蛋白条带变深、相对低分子质量蛋白条带逐渐消失或变淡;分子质量在15~20 kDa内的蛋白条带逐渐变深;10~15 kDa处随着贮藏期延长逐渐出现渐变蛋白条带。由以上蛋白条带变化可知,其发生降解的肌原纤维蛋白出现在37~75 kDa范围内,而其含量增加的肌原纤维蛋白出现在10~20、100 kDa以上范围内,总体变化较为显著,其变化可能会影响蛋白嫩度的形成。贮藏过程中秦川牛背最长肌总可溶性蛋白在内源酶的作用下被降解,而导致总可溶性蛋白含量下降,肌原纤维蛋白含量减少。这与李婷婷报道大黄鱼中不同溶解性蛋白冷藏期间的浓度变化规律一致。
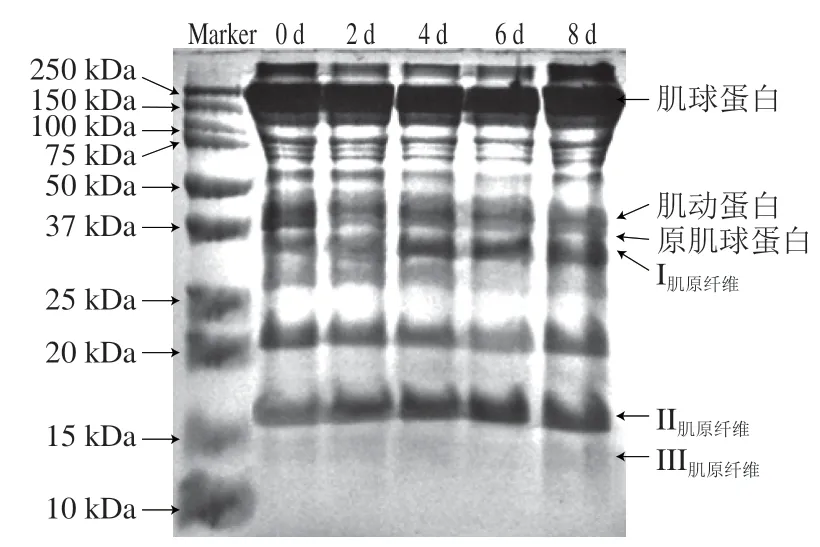
图7 秦川牛背最长肌贮藏过程中肌原纤维蛋白降解电泳图Fig. 7 Degradation of myofibril protein in Qinchuan cattle Longissimus dorsi muscle during storage
2.5 不同贮藏时间秦川牛背最长肌MFI分析
肌纤维降解程度常用MFI来反映。由图8可知,秦川牛背最长肌在整个贮藏过程中MFI呈上升趋势,0~2 d和6~8 d贮藏期上升较缓,2~6 d时上升较快。贮藏0 d时MFI为28.50±5.16,之后MFI一直增加,贮藏8 d时达到整个贮藏过程的最高点(99.98±1.20),增长了250.81%。这一结果与李升升对牦牛平滑肌嫩度形成机理的研究中MFI的变化趋势相一致。
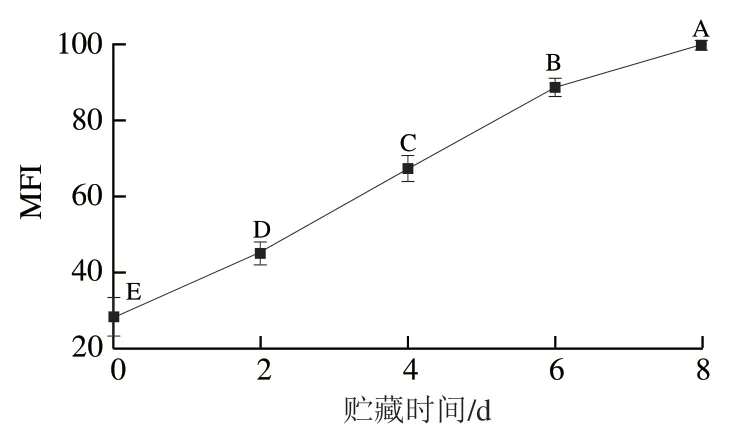
图8 秦川牛背最长肌不同贮藏时间MFI的变化Fig. 8 Changes in MFI of Qinchuan cattle Longissimus dorsi muscle at different storage times
2.6 结构蛋白对宰后秦川牛背最长肌嫩化影响的机制分析
由表1可知,0~8 d贮藏期表征肌纤维降解程度的MFI与剪切力并无显著相关性(=-0.285),D’Alessandro等在牛肉嫩度研究中也出现相同结果。结合大量学者提出的嫩度形成与肌原纤维降解有密切关系,可得贮藏前期肌原纤维小片化程度增大,主要是能量与pH值的变化引起肌肉收缩、组织僵化,进而影响嫩度的降低,对剪切力影响较小。贮藏期内MFI逐渐升高,中后期肌原纤维小片化累积到一定量时对嫩度的形成有贡献。
此外,MFI与总可溶性蛋白、肌原纤维蛋白含量呈极显著负相关(<0.01),与肌浆蛋白含量呈极显著正相关(<0.01)。这说明MFI变化与肌肉蛋白组成变化有密切的关系,蛋白降解会增大MFI。
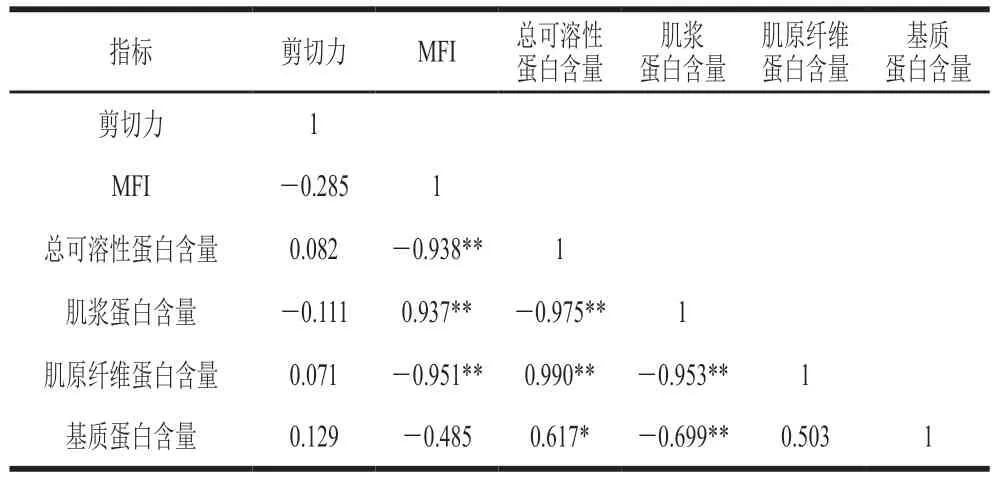
表1 秦川牛背最长肌MFI、总可溶性蛋白、肌浆蛋白、肌原纤维及基质蛋白含量和剪切力之间的相关性Table 1 Correlation between MFI and total protein, sarcoplasmic protein, myofibrillar protein and matrix protein contents and shear force in Qinchuan cattle Longissimus dorsi muscle
肌肉所含蛋白质是由肌浆蛋白、基质蛋白和作为结构蛋白的肌原纤维蛋白组成。由图9可知,贮藏0 d时测得秦川牛背最长肌肌肉中肌原纤维占比69.60%,肌浆蛋白占比22.13%,基质蛋白占比6.60%,符合肌肉组织中蛋白结构组成比例。此外,贮藏0~8 d肌原纤维蛋白所占比例呈持续减少趋势,由0 d时的69.60%逐渐降至8 d 时的52.61%;肌浆蛋白所占比例呈持续上升趋势,由0 d时的22.13%逐渐增加至8 d时的41.23%,但基质蛋白含量仅有少量变化。由此可得肌肉所含蛋白质中主要是肌原纤维蛋白的降解。而肌原纤维蛋白主要由肌动蛋白(45 kDa)、肌球蛋白重链(200 kDa)和原肌球蛋白蛋白(35 kDa)组成,由图7可知,0~8 d贮藏期肌动蛋白与原肌球蛋白所属条带呈显明显变浅趋势,可得肌动蛋白与原肌球蛋白发生降解,在31 kDa附近出现新条带有可能是肌动蛋白的分解产物,作为肌动蛋白降解标志物。对于肌球蛋白重链的降解情况,在总可溶性蛋白凝胶电泳图中发现在200 kDa附近条带变浅,但在肌原纤维蛋白电泳图中未发现有明显变浅的情况,由此无法判断肌球蛋白重链是否出现降解。肌原纤维蛋白降解可能是因为贮藏过程中受到内源酶(钙蛋白酶、组织蛋白酶、细胞凋亡酶)作用,同时微生物对肌肉污染和对蛋白质分解利用也有加速肌动蛋白降解的作用。研究表明肉中肌原纤维蛋白降解会导致其在空间结构、溶解度、疏水性等方面发生改变。肌原纤维是肌肉结构骨架中主要的蛋白质,是肌纤维的物质基础,其出现降解后,MFI逐渐增加,表明肌原纤维内部结构完整性遭到破坏,结合剪切力4~8 d呈下降趋势,可推测出肌肉中结构蛋白发生降解,对嫩度形成可能会有一定影响。

图9 秦川牛背最长肌贮藏过程中蛋白组成占比Fig. 9 Change in protein composition of Qinchuan cattle Longissimus dorsi muscle during storage
2.7 差异蛋白质筛选
实验中采用4D-LFQ检测了秦川牛宰后蛋白质组的变化,对所得的蛋白质二级结构进行质谱检索,共检索确定蛋白1 149 个,设置1.2 倍为标准差异倍数,共筛选出120 个符合标准的差异蛋白质,利用KEGG数据库对这120 种差异蛋白进行生物信息分析,所得的数据利用SPSS 25软件中Pearson相关性分析,共确定11 种差异蛋白质与嫩度形成机理有关,如表2所示。
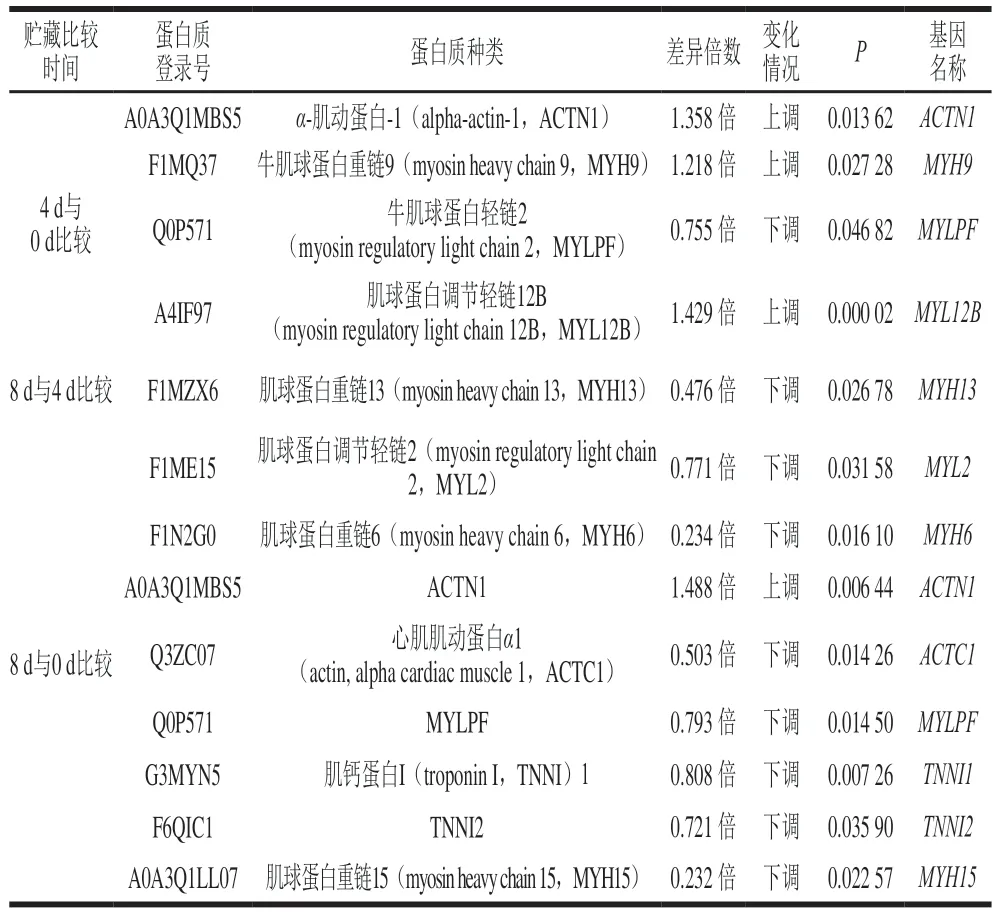
表2 秦川牛背最长肌贮藏0、4、8 d嫩度相关差异蛋白对比Table 2 Comparison of tenderness-related differential proteins in Longissimus dorsi muscle of Qinchuan cattle at days 0 versus 4 versus 8 of storage
2.8 生物信息学分析
图10是利用Cytoscape 3.6.1和String 10.0软件制作筛选出的与嫩度相关的蛋白质互作网络图。此网络图聚类系数为0.759,蛋白质相互作用(proteinprotein interaction,PPI)富集值为1.0×10。ACTN1、MYH9、MYLPF、MYL12B、MYH13、MYL2、MYH6、ACTC1、TNNI1、TNNI2、MYH15的相互作用强烈。以上11 种蛋白在肌球蛋白结合、钙离子结合、细胞骨架蛋白结合的肌原纤维组装、骨骼肌组织发育过程等途径调控细胞的生理状态。
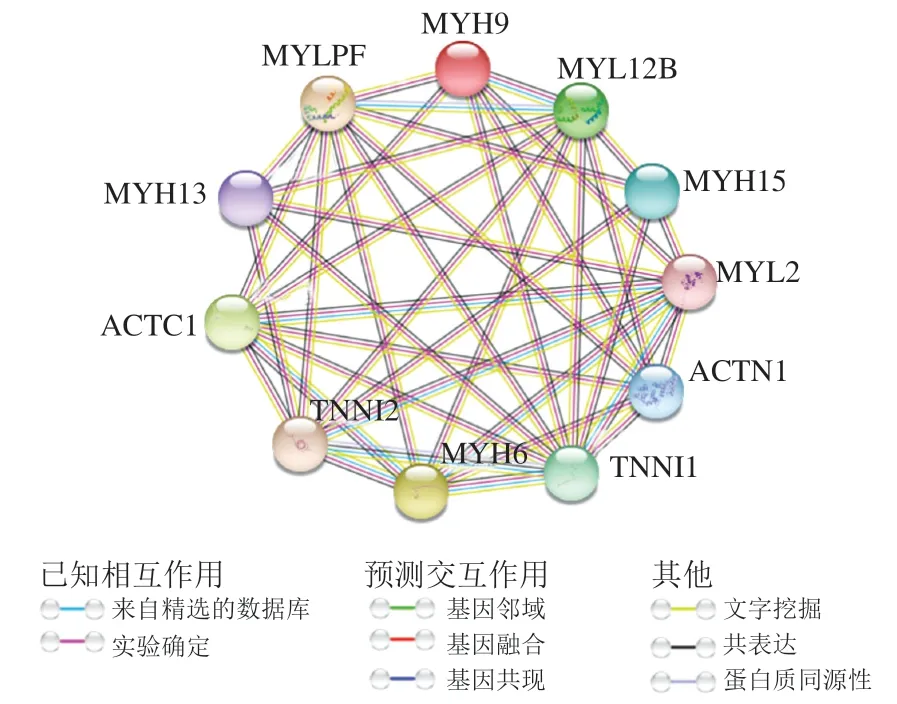
图10 蛋白质互作网络图Fig. 10 Protein-protein interaction network diagram
2.9 结构蛋白变化对秦川牛背最长肌嫩度形成的影响
利用蛋白质组学技术筛选出11 种肌肉结构蛋白(ACTN1、ACTC1、TNNI1、TNNI2、MYH9、MYH13、MYH6、MYH15、MYL2、MYL2B、MYLPF)出现异常表达,丰富度发生了不同程度变化。
实验中发现异常表达的ACTN1与ACTC1均属于肌动蛋白家族。ACTN在细胞骨架蛋白家族广泛表达,通常是以反向平行二聚体形式存在。据报道,ACTN有4 种亚基,ACTN2和ACTN3在肌肉细胞中表达,ACTN1和ACTN4与细胞膜上肌动蛋白偶联。ACTN1被认为能将肌动蛋白锚定在多种细胞内结构上,是一种捆绑蛋白,0~8 d贮藏期内呈现显著上调趋势,推测是由于在肌原纤维束过于松散时的应激反应试图重新固定松散的纤维束,增强骨架结构;ACTC1也属于肌动蛋白家族,与肌原纤维细丝的聚集和形成有关,谢珊珊利用多组学技术整合分析梅山猪,发现ACTC1与肌原纤维形成有关。实验中0~8 d贮藏期内ACTC1表达量显著下调,其低表达可能会影响肌纤维聚集度和结构,进而导致肌原纤维骨架网抗剪切能力降低。
TNNI1与TNNI2均属于肌钙蛋白家族,分别为肌钙蛋白的1型和2型,是肌钙蛋白的重要组成部分。有研究表明TNNI1与TNNI2在肌肉放松和收缩时发挥重要作用,同时影响肌原纤维的直径、类型、形态,肌原纤维这些特征会直接影响肉质,尤其是肌纤维的类型与直径会直接影响牛肉的嫩度。由表2可知,TNNI1与TNNI2表达量在0~8 d贮藏期内均出现显著下调,呈现低表达现象。有研究证明,TNNI1在脊椎动物排酸成熟过程中比较容易降解,且TNNI1发生降解后会伴随着肉品嫩度的增加。综上可得秦川牛宰后肌肉组织通过TNNI1与TNNI2低表达,调节肌肉纤维的直径与形体,进而参与嫩度的形成。
MYH6、MYH9、MYH13、MYH15、MYL2、MYL12B与MYLPF 7 种蛋白均属于肌球蛋白家族。MYH6、MYH9、MYH13与MYH15均属于肌球蛋白激酶ATPase超家族。Fang Xinhui等应用对比实验将敲除,发现在缺氧情况下未敲除组基因的表达量明显上调,与敲除组相比细胞葡萄糖消耗量与乳酸产生量明显增多,这一结果表明MYH9可能会参与糖酵解过程。在本研究中MYH9表达量在0~4 d显著上调,可以看出,秦川牛宰后缺氧情况下,会通过调节MYH9的表达量,增强无氧呼吸的强度,增加产能。MYH6属于肌球蛋白重链家族。付睿琦利用蛋白质组学技术对北京油鸡肌肉基因筛选,结果表明MYH6可能是肌肉细胞骨架结构组成成分,穆琳等研究也表明MYH6可能参与肌纤维发育过程,当肌肉组织的生长速度加快时,MYH6表达水平也会提高。有研究表明,MYH6的低表达会影响心肌的收缩,造成心肌衰竭,由此可判断MYH6还可能与纤维收缩有关。本实验发现MYH6表达量在0~8 d显著下调,其低表达可能会影响秦川牛背最长肌肌肉纤维的再生速度,减少生成量,较难维持肌肉纤维骨架原有状态,并影响了肌肉的收缩功能,增加肌肉的松弛度。查阅文献发现,对MYH13的研究较少,主要集中在眼外肌和喉肌上,MYH13的表达可以使肌肉具有更强的收缩能力和较小的张力。本实验发现MYH13表达量在4~8 d显著下调,可推断出,其低表达可能会使肌肉收缩能力减弱、张力发生变化、肌肉松弛、质地变软、弹性减小、剪切力变小和嫩度变大。MYH15被认为是一种慢性肌球蛋白,参与肌肉的收缩和细胞骨架的重塑,是哺乳动物横纹肌纤维收缩的动力分子。MYH15表达量在0~8 d内显著下调,其低表达可能会降低秦川牛背最长肌横纹肌纤维的收缩能力,使得肌肉长期处于舒张状态。
MYL2、MYL12B和MYLPF为肌球蛋白轻链家族。MYL2参与调解肌原纤维的活性,对肌肉纤维的生长发育有重要的调节作用,通过增加肌球蛋白杠杆臂刚度和促进肌球蛋白头部扩散,从而增加最大收缩力和收缩力的钙敏感性。MYL2表达量在0~8 d显著下调,低表达可能会影响新纤维的生成及损伤纤维的修复过程。MYLPF可磷酸化,属于快速骨骼肌。刘瑞莉等在对布莱凯特黑牛与鲁西黄牛背最长肌中基因表达量的变化规律进行研究时发现,MYLPF可能起调节骨骼肌生长发育的作用。实验中MYLPF表达量在0~4 d显著下调。MYL12B通过磷酸化在调节平滑肌和非肌肉细胞收缩活动中发挥重要作用,还与胞质分裂、受体封顶和细胞运动有关,Park等通过基因敲除实验发现缺失后会导致细胞结构和形态发生显著变化,肌动蛋白纤维的形成会被严重破坏,纤维突起和长度明显增加,MYL12B对于维持MYH9、MYH10和MYL6的稳定性也至关重要,进而对细胞肌动球蛋白的正常功能也具有一定影响。本实验中MYL12B表达量在0~4 d显著上调。通过基因本体注释(gene ontology,GO)分析可以看出MYL2、MYL12B和MYLPF 3 种蛋白共同参与了细胞组分的肌动蛋白细胞骨架和肌球蛋白复合体条目,生物过程中的钙离子结合条目。以上3 种蛋白在贮藏0~8 d出现不同程度调控,对受损纤维修复、纤维再生、肌动蛋白细胞骨架产生不同程度影响,改变肌纤维原有的稳定状态,最终影响纤维骨架强度,使剪切力下降,嫩度上升。
3 结 论
4D-LFQ蛋白质组学分析结果表明,贮藏0~8 d秦川牛背最长肌所含结构蛋白溶解度降低,蛋白质碎片从膜和肌纤维网络如肌纤维网络中释放出来,其中ACTN1、MYH9、MYLPF、MYL12B、MYH13、MYL2、MYH6、ACTC1、TNNI1、TNNI2、MYH15丰富度发生变化,相互作用强烈,在肌球蛋白结合、钙离子结合、细胞骨架蛋白结合的肌原纤维组装、骨骼肌组织发育、肌肉器官发育、横纹肌组织发育过程等途径调控细胞的生理状态,进一步说明贮藏期间,构成肌肉骨架的结构蛋白发生降解,影响肌肉收缩,引起肌肉僵直,从而造成肌肉剪切力增大、MFI增加,对秦川牛肉贮藏中后期嫩度的提高具有一定影响。
