假单胞菌的筛选及其发酵液控制赤潮甲藻的研究
2022-09-14郑璐薇苏玉萍BalajiPrasathBARATHAN
郑璐薇,林 洪,苏玉萍,2,Balaji Prasath BARATHAN ,郑 毅,郑 怡
(1.福建师范大学环境与资源学院、碳中和现代产业学院,福建 福州 350007;2.福建省河湖健康研究中心(福建师范大学),福建 福州 350007;3.福建省特色海洋生物资源可持续利用重点实验室,福建 福州 350117)
有害藻华(harmful algal blooms,HABs)是指水体中一些有毒有害的微藻、大型藻类等生物大量暴发,对生态环境和人类健康造成危害的现象[1].在海洋环境中,常见的有害藻华主要有赤潮、褐潮、绿潮等,在赤潮生物暴发性增殖的过程中,会消耗大量的CO2,导致海水的pH值不断上升,对海洋生物的生长和繁衍产生不利的影响[2].Soto等[3]研究了几乎每年都会出现在佛罗里达沿岸海湾的由短凯伦藻(Kareniabrevis)引起的赤潮,其在2017年持续了15个月,导致大面积缺氧事件,严重危害了当地石蟹的生存.一些赤潮生物分泌的粘液会附在海洋动物的鳃上,使其无法呼吸,最后导致它们窒息死亡,造成渔业损失,如夜光藻(Noctilucascintillans)、血红哈卡藻(Akashiwosanguinea)等[4].部分有毒赤潮藻暴发时会产生毒素,引起鱼类死亡,如链状裸甲藻(Gymnodiniumcatenatum)、米氏凯伦藻(Kareniamikimotoi)、球形棕囊藻(PhaeocystisglobosaScherffel)等[5].更为严重的是,藻毒素会通过食物链在海鱼、牡蛎贝类等海产品的体内富集,居民误食了受污染的海产品会导致腹泻、中毒,甚至死亡.有害赤潮消亡时,死鱼的大量分解会释放二氧化硫等有害物质,改变水体的理化性质,对海洋生态系统、渔业养殖生产、旅游业以及人类健康产生严重威胁[6].我国每年4-6月频发的米氏凯伦藻和链状裸甲藻等有毒甲藻赤潮,严重影响居民健康和沿海经济发展.在近年的海洋观测中发现,福建沿海的米氏凯伦藻的藻密度居高不下[7-8],链状裸甲藻孢囊分布密集,为赤潮的暴发埋下了一定的隐患[9].
迄今为止,控制有害赤潮的方法主要可分为物理、化学和生物3类方法[10].生物法具有特异性较强、低成本、低毒、环境友好等特点,细菌在赤潮发生和消亡过程中起到了重要的作用,因此也是生物法控制HABs的研究热点[11-12],目前分离鉴定出了数百种能够溶解藻细胞或抑制藻类生长的溶藻菌(Algicidal bacteria),主要有芽孢杆菌属(Bacillus)、假单胞菌属(Pseudomonas)、短杆菌属(Brevibacterium)等[13-15].Skerratt等[16]发现交替假单胞菌对链状裸甲藻有积极的杀藻作用,Zheng等[17]研究则发现黄杆菌属和交替假单胞菌属的细菌培养滤液对米氏凯伦藻的溶藻效果明显.以上研究说明了溶藻菌在控制赤潮领域具有广泛的应用潜力.
目前,国内外对链状裸甲藻的研究仍主要停留在底泥孢囊的监测、藻类生长以及藻毒素处理,针对其采用溶藻菌进行调控的研究较少[18-19].本研究筛选出一株特异性溶藻菌Ps3,探究其发酵液不同组分的溶藻效果.由于一些溶藻菌可分泌多种特定的溶藻物质,宿主范围较广[20],而链状裸甲藻、米氏凯伦藻均为福建沿海常见的有毒甲藻,具有广温广盐的特性,已成为我国近海海域典型的海洋生态灾害的原因种[8,21].因此利用菌Ps3对米氏凯伦藻和链状裸甲藻进行溶藻效果探究,对海洋甲藻赤潮的控制具有现实意义.
1 材料与方法
1.1 实验藻种的培养
链状裸甲藻、米氏凯伦藻均购自厦门大学近海海洋环境科学国家重点实验室.选用L1培养基[22],培养条件设置为温度(20±1)℃,光照强度3 000 lx,光暗比12 h∶12 h.每日手动摇2次,防止赤潮藻贴壁生长,根据其生长状态,取一定体积处于对数生长期状态的赤潮藻接种到新鲜营养液中,进行传代培养[23].
1.2 实验菌种的培养
本实验使用的5株溶藻菌(Ps1-Ps5)是由项目组福建师范大学生命科学学院合作团队从海洋虾池中分离保存的菌种.将初代保存于4 ℃的菌Ps1-Ps5微生物斜面取出,在经紫外灭菌的超净台上各挑取一满环接种于50 mL牛肉膏蛋白胨液体培养基中,置于恒温摇床中30 ℃、130 r·min-1条件下培养24 h,即为复壮种子液.从复壮种子液中再重新取一满环接种于50 mL培养基,在30 ℃、130 r·min-1培养24 h.同时对菌株种子液进行平板划线,检验无染菌情况[24].
1.3 菌-藻共培养实验
将保存于-80 ℃的甘油(质量分数50.0%)待试菌置于4 ℃下解冻2 h.在紫外灭菌后的超净台中用接种针在固体培养基上进行平板划线.于恒温振荡培养箱中30 ℃、180 r·min-1条件下培养24 h,检查菌落的均一性.从平板中挑取一满环菌株接种到含50 mL培养液的100 mL锥形瓶中,将锥形瓶置于恒温培养箱中,条件设置为30 ℃、180 r·min-1.将5株已复苏的菌株培养48 h后的菌液按体积分数1.0%分别加入处于指数生长期的链状裸甲藻.以加入等体积的培养基为对照,分别于0、4、8、12、18、24、36、48、72 h取样测定藻细胞密度,每组样本设置3个平行,计算溶藻率,见公式(1).
(1)
式中R为溶藻率;ρ0为对照组藻细胞密度,L-1;ρ1为处理组藻细胞密度,L-1.
1.4 溶藻显微观察
每次取0.1 mL已固定的藻样品于0.1 mL浮游植物计数框,置于江南生物显微镜BM2000上观察藻细胞结构变化并拍照.根据Lovejoy等[25]设定的细胞死亡特征(MSE)方法,用显微镜观察判断藻细胞是否死亡.MSE主要为:运动性丧失(细胞无运动性)、崩解(细胞膜破裂,内部结构紊乱)、裂解(细胞迅速渗漏,细胞器破坏)或蜕皮(细胞膜脱落).
1.5 分子生物学鉴定
将该菌株用液体牛肉膏培养基培养至生长期,在无菌条件下,移取2 mL至无菌离心管,冷藏寄送至北京奥维森基因有限公司进行16S rDNA鉴定,包括DNA提取、PCR扩增、序列测定等工作.
PCR扩增的上游引物为27F(5′-AGAGTTTGATCCTGGCTCAG-3′),下游引物:1492R (5′-TACGGCTACCTTGTTACGACTT-3′).在美国国立生物技术信息中心(NCBI)的GenBank数据库中用局部相似性基本查询工具(BLAST)进行比较,初步确认细菌种属.在MEGA7.0软件中使用邻接法(Neighbor-Joining),用极大似然估计模型(Maximum Composite Likelihood),计算1 000次自展值,构建系统发育树.将测得的菌Ps3的16S rDNA序列信息提交到GenBank (http:∥www.ncbi.nlm.nih.gov).
1.6 发酵液组分分离
利用极性溶剂对细菌发酵液进行粗分离,观察不同组分的溶藻效果.以等量的培养基为对照,取100 mL培养至稳定期的菌Ps3发酵液,用0.22 μm孔径玻璃纤维滤膜重复过滤3次,得到Ps3无菌发酵液.将Ps3无菌发酵液与乙酸乙酯等体积混合后在180 r·min-1振荡20 min,在分液漏斗中静置分层,收集有机相溶液,重复萃取3次.在80 ℃下将萃取相进行减压蒸馏蒸干,最后加入5 mL纯乙酸乙酯溶解,定容至10 mL,为乙酸乙酯相提取溶液;剩余物质进行减压蒸馏蒸干,用5 mL水溶解并定容至10 mL,获得剩余相溶液,均在-4 ℃避光保存备用.
1.7 不同组分溶藻效果实验
以纯乙酸乙酯为对照,将乙酸乙酯相提取溶液、剩余相水溶液分别按体积分数0.1%、0.5%、1.0%、2.0%、4.0%的投加量加入链状裸甲藻和米氏凯伦藻中,培养温度为(20±1) ℃,光照强度为3 000 lx、光暗比为12 h∶12 h.定时取样,计算藻细胞数目,取3次平行样,在48 h取样并计算数量,观察溶藻效果,测定相应的溶藻率.
1.8 数据分析
实验数据使用Excel2019、Origin2021软件进行作图,SPSS24.0对溶藻数据进行分析(P<0.05为显著性差异),MEGA7.0构建系统发育树.
2 结果与讨论
2.1 溶藻细菌基本形貌特征
从海洋养虾池中分离得到5株菌,编号分别为Ps1-Ps5.在培养管中各菌株特征如图1所示,Ps1、Ps2的菌落表面较透明,边缘光滑整齐,可能为具有荚膜类的菌种;Ps3的菌落边缘不规则;Ps4的菌落表面干燥皱褶;Ps5单菌落为鲜明的黄色.各菌落在颜色、光泽及形状方面差异较为明显.

图1 菌Ps1-Ps5的基本形貌Fig.1 Basic morphology of bacteria Ps1-Ps5
2.2 共培养下5株细菌对链状裸甲藻生长的影响
经过一段时间的培养,在48 h内对照组链状裸甲藻细胞密度从9.5×104L-1增长到1.7×105L-1,表现出良好的生长活性.而在与细菌共同培养的过程中,各组表现出了不同的抑藻效果(图2).从图2a、2e可看出,0~18 h链状裸甲藻生长滞缓,18 h之后藻细胞密度稳步增长.这表明链状裸甲藻对菌Ps1、Ps5具有一定抵抗能力,能够适应菌Ps1、Ps5带来的藻际环境的变化.图2b中,前24 h链状裸甲藻的生长受到菌Ps2的抑制,但在24~72 h内藻密度变化并不明显,可能是菌与藻形成了拮抗作用.从图2c、2d可看出,菌Ps3、Ps4均表现出较好的溶藻能力.将5株细菌48 h的溶藻率进行对比,从高到低分别为Ps3>Ps4>Ps2>Ps1>Ps5(图2f).溶藻效果最好的是菌Ps3,48 h溶藻率达到87.4%;其次是Ps4,48 h的溶藻率为78.0%.
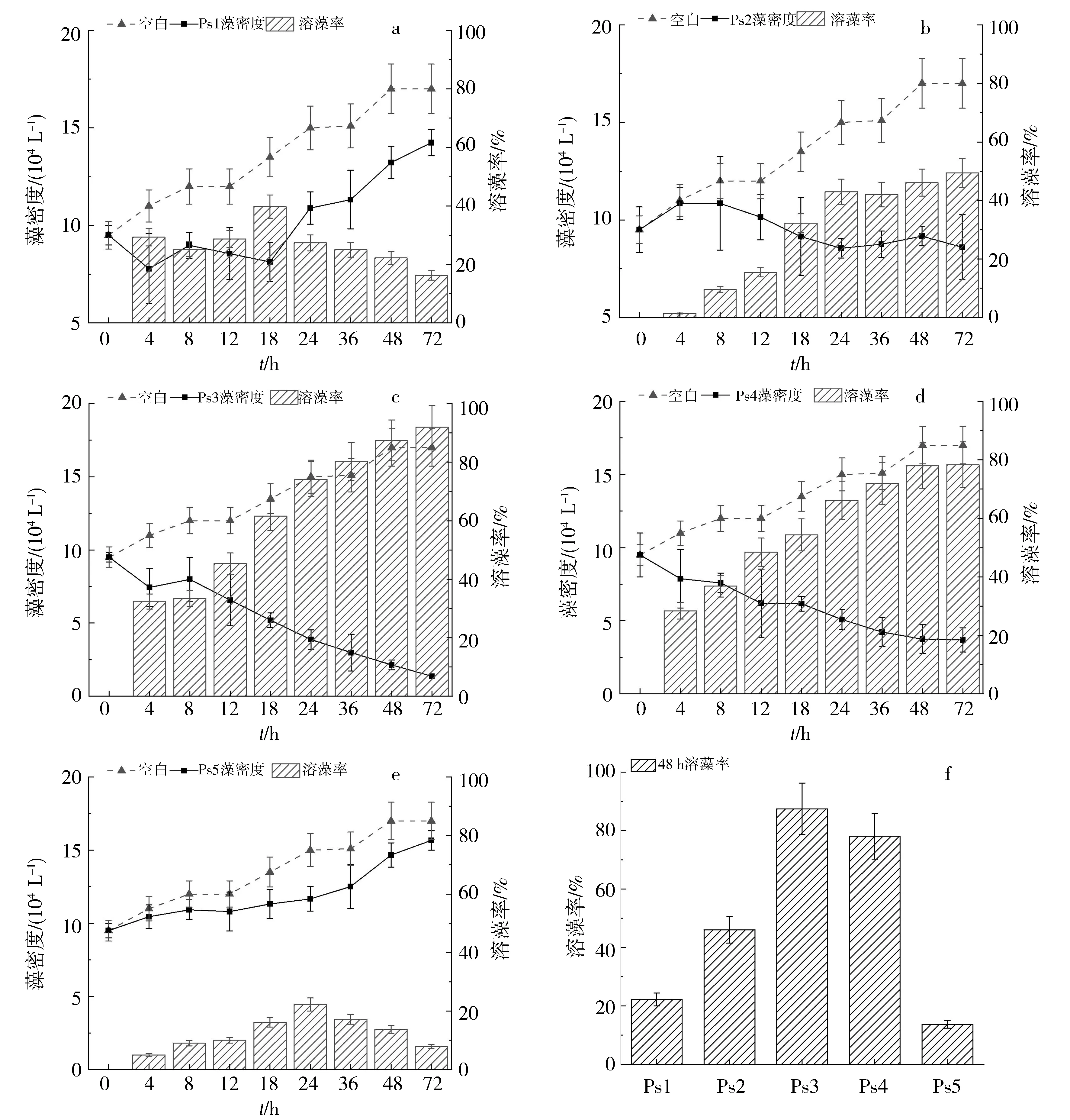
图2 共培养下菌Ps1-Ps5对链状裸甲藻的抑藻能力Fig.2 The algicidal activity of bacterial Ps1- Ps5 on Gymnodinium catenatum under co-culture
链状裸甲藻个体较大,可以在低倍的光学显微镜中观察到藻体形态变化.菌藻共培养后,从图3可观察到各实验组剩余可辨识藻体的主要形貌.在与菌Ps1、Ps5分别共同培养后的藻细胞内部结构保持较好,横沟清晰,具有活动性(图3a及图3e).与菌Ps2共培养的藻细胞膜结构完整,但内部结构较模糊(图3b);而菌Ps3作用后,可以看到藻体内部结构紊乱,横沟消失,细胞膨胀(图3c);图3d则观察到与菌Ps4共同培养的藻细胞膜稍有变形,无细胞质渗漏.
根据菌-藻共培养实验和镜检结果综合判断,菌Ps3的溶藻能力明显高于其它菌株,因此选择菌Ps3做进一步研究.
2.3 菌Ps3的菌落及菌体形态
从图4a可观察到Ps3的单菌落呈圆形颗粒状,稍有隆起,菌落直径在1~3 mm之间,表面光滑湿润,乳白色不透明.Ps3的革兰氏染色情况如图4b所示,染色结果为蓝色,判断为革兰氏阴性菌.使用扫描电镜进一步观察,单个Ps3菌体形态如图4c所示,为短杆状,大小为(1.5~1.7) μm×(1.1~ 1.3) μm,多个头尾相连,无鞭毛,未观察到芽孢存在.
2.4 菌Ps3的分子生物学鉴定
对菌Ps3的16S rDNA基因序列进行扩增,根据鉴定结果可知,该菌有碱基对1 437 bp.在NCBI上(http:∥www.ncbi.nlm.nih.gov)上传菌种的基本情况和基因序列,登录号为OK103600.1.将菌Ps3的基因序列进行BLAST比对发现,与菌Ps3的高相似菌种均为假单胞菌属(Pseudomonas),相似度皆达到99.0%以上(表1).

表1 菌Ps3在BLAST库比对结果Tab.1 Results of strain Ps3 in BLAST database comparison
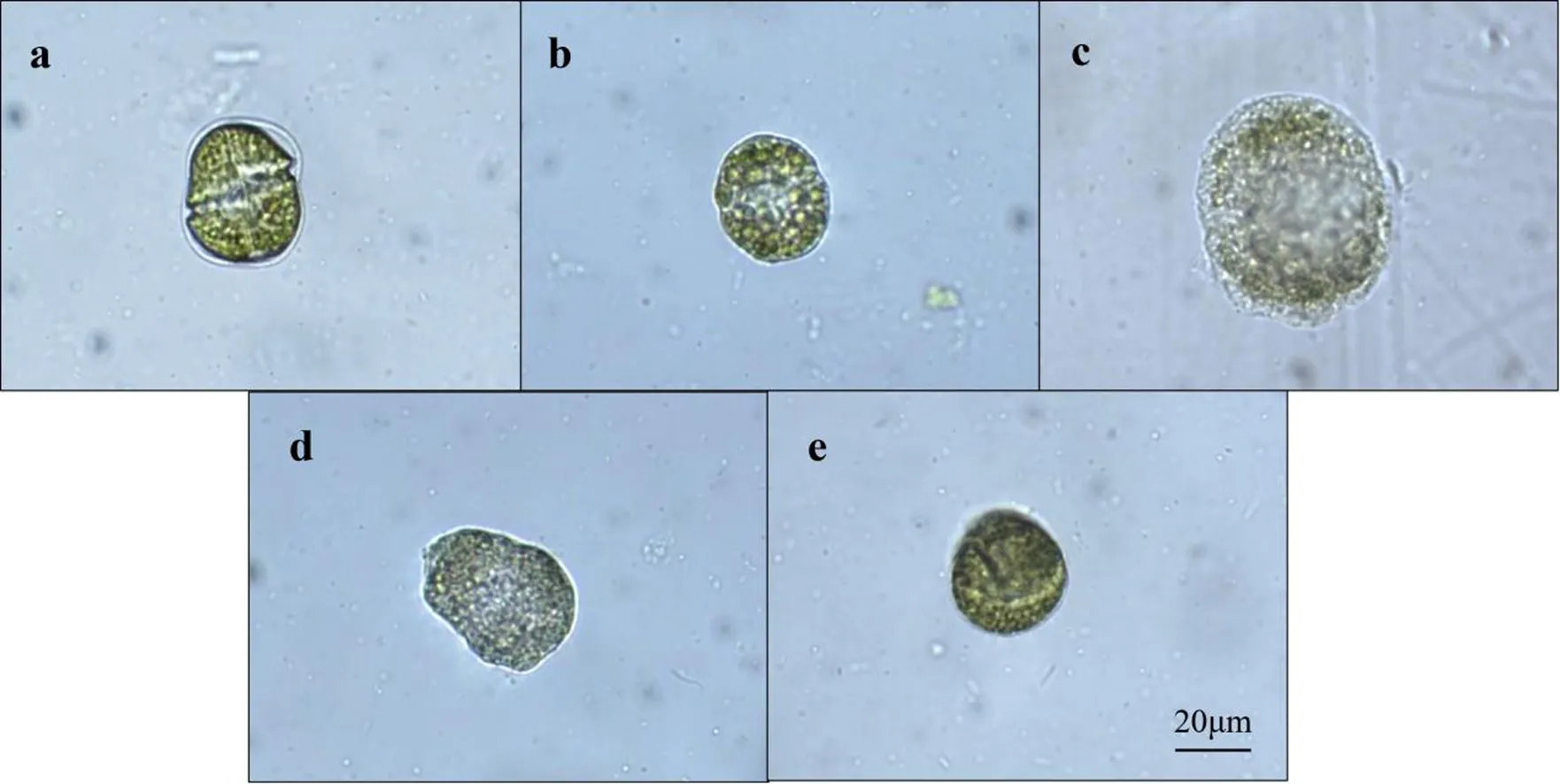
a-e分别表示与菌Ps1-Ps5共培养后的藻细胞图3 菌藻共培养48 h后可辨识的藻体主要形貌Fig.3 Identifiable algal cells after 48 h of co-culture
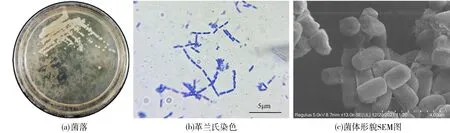
图4 Ps3菌落的基本形态Fig.4 Basic morphology of bacterial Ps3
本研究选取了合适的菌株基因序列,构建的系统发育树如图5所示.与Ps3同源性比较高的菌株为Pseudomonassp.JC5和Pseudomonasprotegensstrain CP-M2-5,三者位于同一分支上.菌Ps3也与Pseudomonasplecoglossicidastrain 2-3有较高的同源性,二者均处于一个较大的分支上的两个分支.
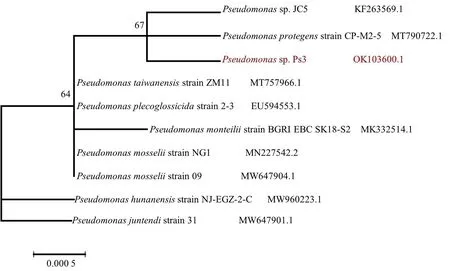
图5 假单胞菌Ps3的系统发育树Fig.5 Phylogenetic trees of Pseudomonas Ps3
2.5 菌发酵液分离组分的溶藻效果
在链状裸甲藻和米氏凯伦藻中分别投加不同体积分数的乙酸乙酯后,48 h内藻细胞密度基本持平,并无明显改变.因此投加乙酸乙酯对溶藻实验的影响可以忽略.
将不同体积分数的乙酸乙酯提取相溶液和剩余相溶液分别投加到链状裸甲藻后,实验结果如图6a所示,体积分数0.1%和0.5%乙酸乙酯提取相溶液对藻的溶解效果较差,分别为10.4%和24.6%;投加体积分数1.0%的乙酸乙酯提取相溶液后,48 h溶藻率明显增加,为80.5%;而投加体积分数4.0%的提取相溶液之后,溶藻率达到98.4%.相较之下,投加体积分数4.0%的剩余相溶液48 h之后溶藻率仅为24.5%,表明剩余相的溶藻能力较差.
在米氏凯伦藻中(图6b),投加体积分数0.1%和0.5%乙酸乙酯提取相溶液后,溶藻率分别为15.4%和54.7%,高于链状裸甲藻组,表现出较好的效果;而体积分数4.0%乙酸乙酯提取相溶液的溶藻率达到95.9%,相较其它组有显著提升(P<0.05).值得注意的是,不同于链状裸甲藻,体积分数4.0%的剩余相溶液对米氏凯伦藻的溶藻率也有45.0%,作用显著(P<0.05),可能是由于藻的种类以及溶藻机理的不同,存在一定差异[26].实验结果表明,假单胞菌Ps3的活性物质主要在乙酸乙酯提取相中.
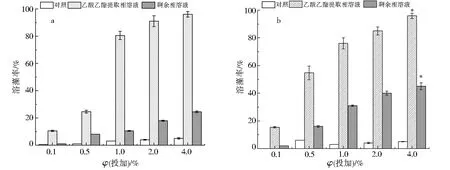
图6 假单胞菌Ps3发酵液中的乙酸乙酯提取相与剩余相对链状裸甲藻(a)和米氏凯伦藻(b)的溶藻作用Fig.6 The algal lysis of Gymnodinium catenatum (a) and Karenia mikimotoi (b) by ethyl acetate extraction phase and residual phase in the fermentation broth of Pseudomonas sp.Ps3
3 结论
(1)本实验采用菌藻共培养的方法,筛选出一株溶藻效果好的菌Ps3,革兰氏染色结果为蓝色,判断为革兰氏阴性菌;扫描电镜观察,单个Ps3菌体为短杆状,大小为(1.5~1.7) μm×(1.1~1.3) μm.结合形态学、生理生化和测序比对结果,鉴定菌Ps3为假单胞菌属(Pseudomonas).
(2)采用乙酸乙酯作为发酵液提取相,将不同体积分数投加量下乙酸乙酯提取相溶液和剩余相溶液对链状裸甲藻和米氏凯伦藻的溶藻率进行比较,剩余相溶液的溶藻能力较差,投加体积分数4.0%时对链状裸甲藻的溶藻率仅为24.5%,对米氏凯伦藻的溶藻率为45.0%;假单胞菌Ps3的活性物质主要在乙酸乙酯提取相中,4.0%乙酸乙酯相提取液对链状裸甲藻和米氏凯伦藻的溶藻效果是最好的,48 h溶藻率分别为98.4%和95.9%.
