红松胚性愈伤组织增殖的激素配比、糖源类型和增殖周期效应研究
2022-08-04任毓辉彭春雪沈海龙
任毓辉 聂 帅 彭春雪 杨 玲 沈海龙
(东北林业大学林学院,林木遗传育种国家重点实验室,哈尔滨 150040)
红松()为松科(Pinaceae)松属()高大乔木树种,是我国长白山和小兴安岭林区红松阔叶林的建群种,既是珍贵用材树种,也是优质食用坚果生产树种,是具有很高经济价值和生态价值的珍贵乡土树种。红松良种选育工作已经取得一些成绩,获得了一些优良种质材料,包括杂交育种材料。这些少量的优良种质材料需要经过规模化扩繁,才能满足造林需求,而体细胞胚胎发生技术可以利用少量繁殖材料短时间内扩繁大量苗木,是实现这个目的的理想途径。红松组织培养最早的报道是1962年Lee等的胚培养、1986 年Kim 等获得愈伤组织、1978 年黑龙江省林业科学研究所获得不定芽,而最早的胚状体(体胚)是1992年东北林业大学所获得,但只到球形的原始阶段,未获得进一步发育。2005年申晓辉等报道获得红松胚性愈伤组织,2017 年高芳等对红松胚性愈伤组织诱导的培养基类型、糖源浓度和植物生长调节剂组合及外植体取材部位等进行了筛选,随后科研人员对红松体胚发生的外植体及其预处理技术,胚性愈伤组织诱导、增殖和超低温保存技术、体胚成熟促进和体胚的生理生化特性等进行了系统研究。总体来看,目前红松体胚发生已经取得重大进步,但是在胚性愈伤组织增殖过程中的植物生长调节剂组合、碳源种类和浓度尚有很大改进空间,合适的增殖周期尚未确立,生理生化特性尚不明晰。因此,本研究对红松胚性愈伤组织增殖培养基中的碳源类型、生长调节剂浓度和种类、增殖培养周期以及增殖中的胚性愈伤组织生理变化进行进一步研究,以期为优化红松体胚发生体系提供参考。
1 材料与方法
1.1胚性愈伤组织材料及增殖保存方法
以3 月龄的红松1-100 号细胞系为试材。增殖基本培养基在方案上微调:mLV 添加2 mg·L2,4-D,0.5 mg·L6-BA,25 g·L蔗糖,0.5 g·L酸水解络蛋白,0.5 g·L-谷氨酰胺,4 g·L结冷胶。所有培养基在灭菌前调节pH 为5.8,121 ℃高压灭菌30 min。在(25±2)℃条件下暗培养,2周继代1次。
1.2增殖培养的激素浓度效应研究
取胚性愈伤组织试材接种到含以下不同激素浓度的增殖培养基中。处理1,2 mg·L2,4-D+0.5 mg·L6-BA;处理2,1 mg·L2,4-D+0.25 mg·L6-BA;处理3,2 mg·LNAA+0.5 mg·L6-BA;处理4,1 mg·LNAA+0.25 mg·L6-BA;处理5,无激素。每个处理重复5 次。激素外增殖基本培养基其他成分和培养条件同1.1。在增殖培养14 d 后收集材料进行体胚分化培养和生理指标测定。
1.3增殖培养的碳源效应研究
取胚性愈伤组织试材接种到以下4 种不同碳源的增殖培养基上。处理6,25 g·L蔗糖;处理7,25 g·L麦芽糖;处理8,25 g·L果糖;处理9,25 g·L葡萄糖。每个处理重复5 次。碳源外增殖基本培养基其他成分和培养条件同1.1。在增殖培养14 d后收集材料进行体胚分化培养和生理指标测定。
1.4增殖培养的增殖周期效应研究
取红松胚性愈伤组织试材接种至增殖培养基中,设定增殖培养周期为以下2 种:7 d 和14 d。每个处理重复5 次。增殖基本培养基和培养条件同1.1。在增殖培养第7和第14天收集材料进行体胚分化培养和生理指标测定。
1.5体胚分化培养
将经过不同增殖培养处理的胚性愈伤组织接种到体胚分化培养基上诱导体胚分化。分化培养基基于Klimaszewska 等的方案:mLV+80 μmol·LABA+1.2%结冷胶+0.2 mol·L蔗糖,pH=5.8。黑暗中培养8 周后统计发生的体胚数量。培养60 d后统计每克胚性愈伤组织鲜质量的体胚发生数量,计算公式如下:

1.6生理指标测定
将经过不同增殖培养处理的胚性愈伤组织接种到相应类型的增殖培养处理组合的培养基上培养,取材进行生理生化指标的测定。采用考马斯亮蓝G-250法测定可溶性蛋白的质量分数(按样本鲜质量计算),采用蒽酮比色法测定可溶性糖及淀粉质量分数(按照蛋白浓度计算),采用氯化硝基四氮唑蓝法测定SOD 活性(按照蛋白浓度计算),愈创木酚法测定POD 活性(按照蛋白浓度计算),过氧化氢法测定CAT 活性(按照蛋白浓度计算)。每个处理3 次重复,每次重复分别收集0.5 g的材料。
1.7统计分析
试验数据用Excel 2003 进行统计处理,利用SPSS19.0 软件进行多重比较(Duncan’s)和方差分析。显著性差异水平=0.05。最后用SigmaPlot 12.5软件进行绘图制作。
2 结果与分析
2.1增殖培养的激素浓度对体胚产量的影响
增殖培养基中不同激素浓度处理下的红松胚性愈伤组织经过分化培养均有体胚发生,但不同浓度的生长调节剂组合显著影响红松体胚产量(<0.05)。其中,在1 mg·LNAA+0.25 mg·L6-BA浓度处理下体胚产量最高,为166 个·g,在2 mg·L2,4-D+0.5 mg·L6-BA 浓度浓度下体胚为127 个·g。而激素浓度为0的处理组合中体胚产量最低,为13个·g(见图1)。
2.2增殖培养的碳源效应
增殖培养基中不同碳源处理下红松胚性愈伤组织经过分化培养后,碳源为蔗糖的处理分别与碳源为麦芽糖、果糖、葡萄糖的处理体胚产量之间差异显著(<0.05),但是碳源为麦芽糖、果糖和葡萄糖的处理体胚产量之间差异相互不显著。其中碳源为蔗糖的处理体胚产量最高为127 个·g,其次是碳源为麦芽糖的处理(80 个·g)。碳源为葡萄糖的处理体胚产量最低为53 个·g。碳源为蔗糖的处理相较于碳源为葡萄糖的处理其体胚产量增加了75 个·g(见图1)。
2.3增殖培养的增殖培养周期效应
增殖培养中不同增殖培养周期处理下红松胚性愈伤组织经过分化培养后,增殖培养14 d 的处理和增殖培养7 d 的处理其体胚产量之间差异显著(<0.05)。增殖培养7 d 的处理体胚产量最低为47 个·g。增殖培养14 d 处理的体胚产量远高于增殖培养7 d 的处理,约为增殖培养7 d 处理体胚产量的2.7倍(见图1)。
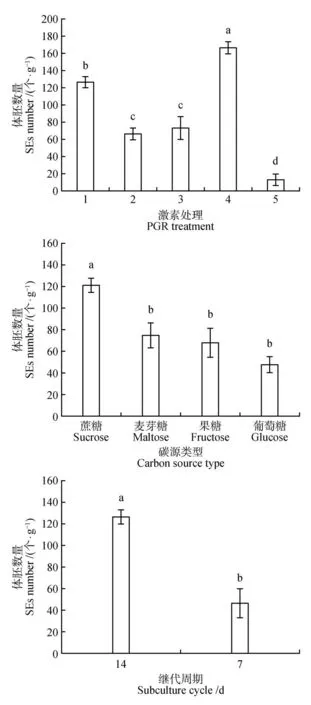
图1 激素、碳源类型和增殖培养周期对红松体胚产量的影响处理1. 2 mg·L-1 2,4-D+0.5 mg·L-1 6-BA、蔗糖、继代14 d;处理2.1 mg·L-1 2,4-D+0.25 mg·L-1 6-BA、处理3. 2 mg·L-1 NAA+0.5 mg·L-1 6-BA;处理4.1 mg·L-1 NAA+0.25 mg·L-1 6-BA;处理5.无激素;同一测定指标上不同字母代表P<0.05差异显著;下同Fig.1 Effects of hormones,carbon source types and proliferation culture cycle on somatic embryo yield of P.koraiensisTreatment 1. 2 mg·L-1 2,4-D+0.5 mg·L-1 6-BA,sucrose,subculture for 14 d;Treatment 2. 1 mg·L-1 2,4-D+0.25 mg·L-1 6-BA;Treat‑ment 3. 2 mg·L-1 NAA+0.5 mg·L-1 6-BA;Treatment 4. 1 mg·L-1 NAA+0.25 mg·L-1 6-BA;Treatment 5. No hormone;Different letters on the same measurement index represent P<0.05,the same as below
2.4不同增殖培养处理方式的生理特性
在红松愈伤组织增殖培养过程中,不同增殖培养方式显著影响红松胚性愈伤组织细胞内的可溶性糖和淀粉质量分数(<0.05)。在不同激素处理中,无激素处理可溶性糖质量分数最低(1.54 g·g),1 mg·L2,4-D+0.25 mg·L6-BA 处理可溶性糖质量分数最高(2.50 g·g)。在不同的碳源类型中,碳源为蔗糖的处理与其他碳源处理可溶性糖质量分数差异显著(<0.05)。碳源为葡萄糖的处理可溶性糖质量分数最低为0.77 g·g。在不同增殖培养周期中,增殖培养7 d的处理可溶性糖质量分数最高(2.75 g·g)为增殖培养14 d处理的1.14倍。在所有增殖培养类型中,对照处理淀粉质量分数最高(6.53 g·g)。在不同激素处理中,1 mg·L2,4-D+0.25 mg·L6-BA,2 mg·LNAA+0.5 mg·L6-BA,1 mg·LNAA+0.25 mg·L6-BA 处理的淀粉质量分数之间差异不显著。在不同的碳源类型中,碳源为葡萄糖的处理中淀粉质量分数最低(2.05 g·g),约为对照(碳源为蔗糖)处理淀粉质量分数的1/3(图2~3)。
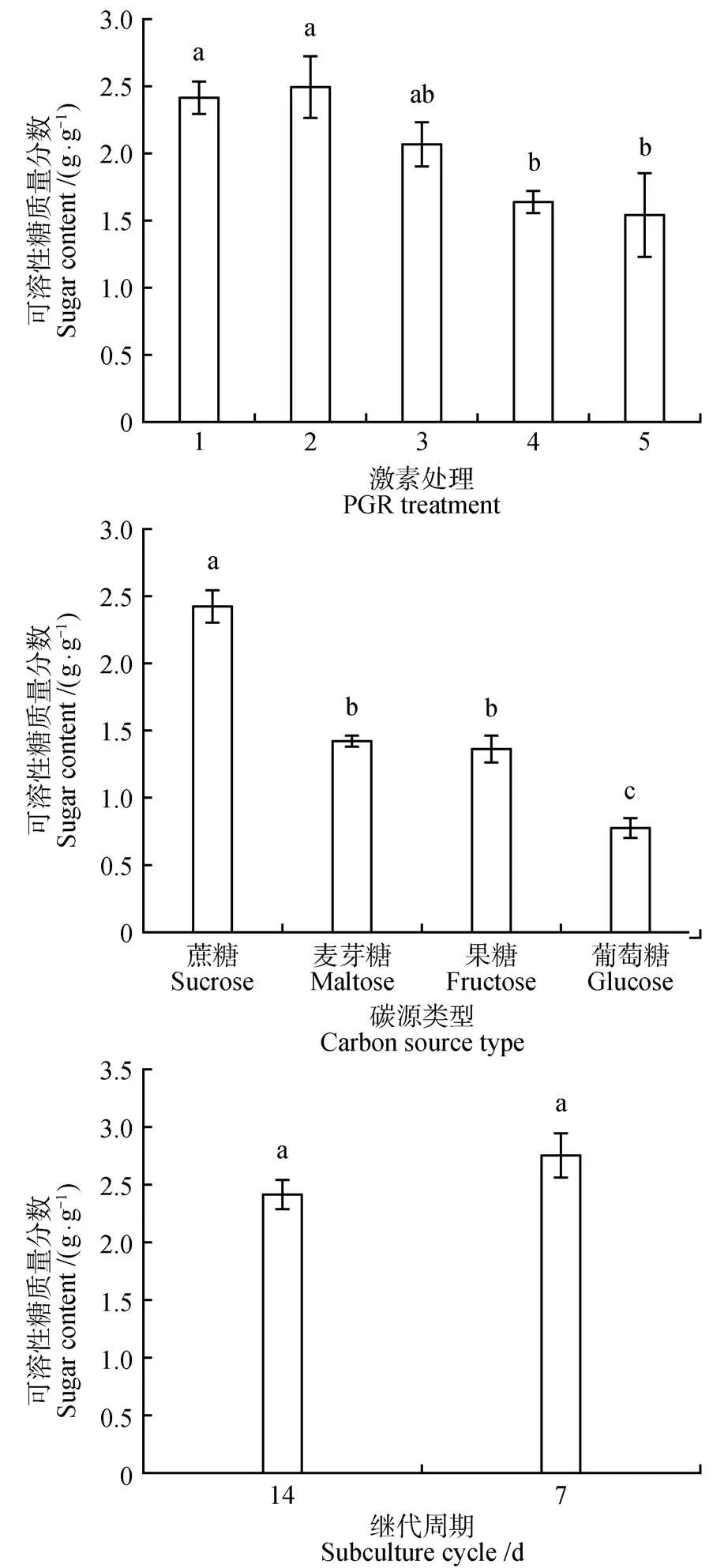
图2 激素、碳源类型和增殖培养周期对红松胚性愈伤组织可溶性糖质量分数的影响Fig.2 Effects of hormones,carbon source types and proliferation culture cycle on soluble sugar mass fraction in embryogenic callus of P.koraiensis
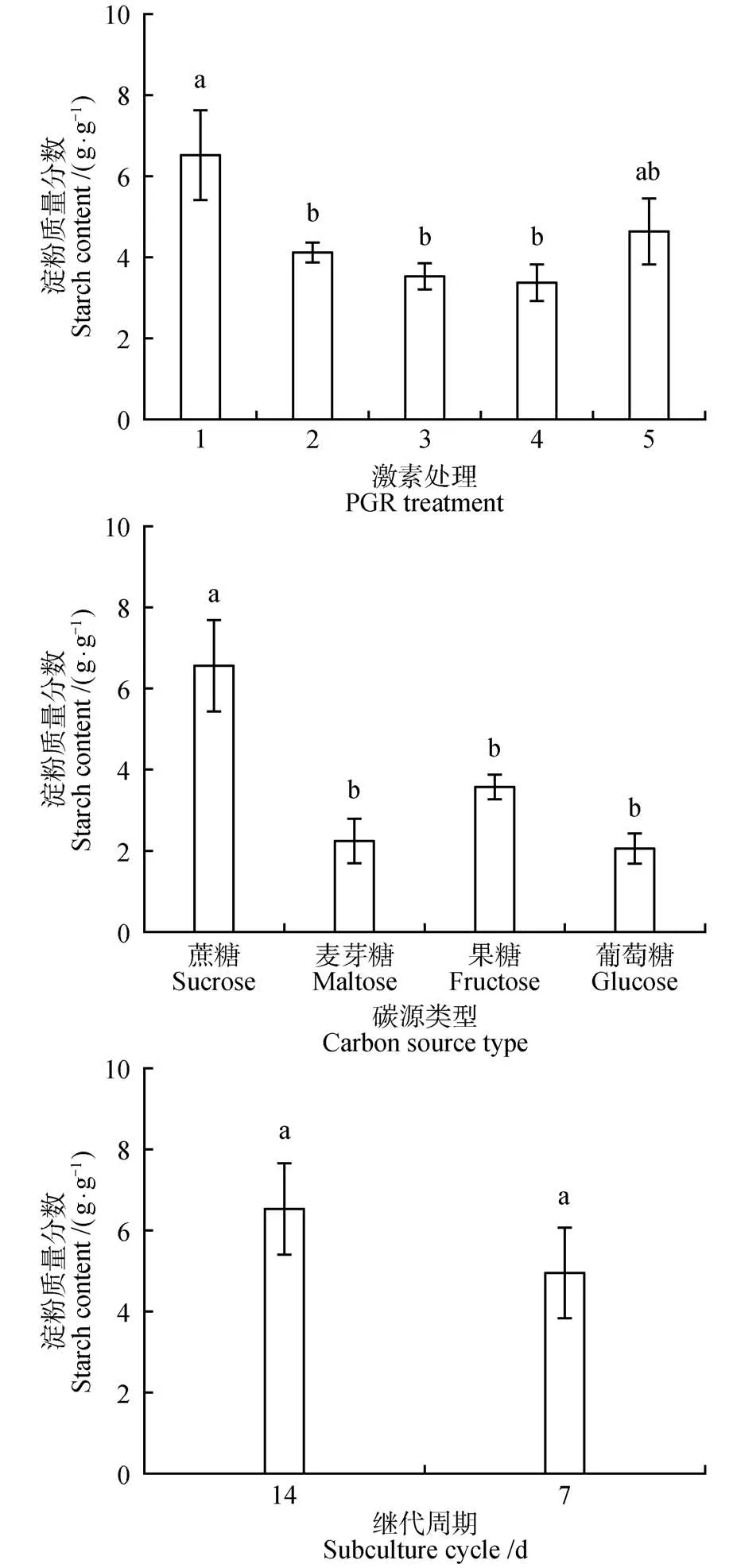
图3 激素、碳源类型和增殖培养周期对红松胚性愈伤组织可溶性淀粉质量分数的影响Fig.3 Effects of hormones,carbon source types and proliferation culture cycle on soluble starch mass fraction in embryogenic callus of P.koraiensis
2.4.2 可溶性蛋白分析
不同的激素处理的红松胚性愈伤组织可溶性蛋白质量分数差异不显著(=0.199)。其质量分数保持在0.81 mg·g左右。1 mg·L2,4-D+0.25 mg·L6-BA处理可溶性蛋白质量分数最低,为0.67 mg·g左右,与对照的蛋白质质量分数相似。在不同的碳源类型中,碳源为麦芽糖的处理分别与碳源为蔗糖和果糖的处理可溶性蛋白质量分数差异显著,与碳源为葡萄糖的处理差异则不显著。碳源为麦芽糖的处理可溶性蛋白质量分数最高为0.96 mg·g(见图4)。
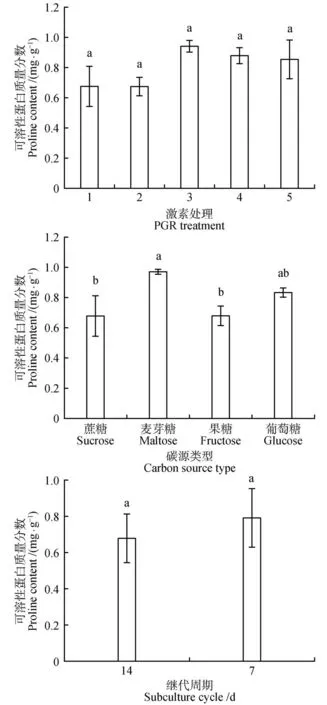
图4 激素、碳源类型和增殖培养周期对红松胚性愈伤组织可溶性蛋白质量分数的影响Fig.4 Effects of hormones,carbon source types and proliferation culture cycle on soluble protein mass fraction in embryogenic callus of P.koraiensis
在红松愈伤组织增殖过程中,不同增殖培养方式对红松愈伤组织CAT、POD、SOD 活性差异显著(<0.05)。在不同激素类型中,2 mg·LNAA+0.5 mg·L6-BA 处理CAT 活性最高(129.70 μmol·min·g),相较于对照增加了约3 倍(见图5)。1 mg·L2,4-D+0.25 mg·L6-BA处理POD活性最高(2 157.60 U·mg),其次是2 mg·LNAA+0.5 mg·L6-BA 处理POD 活性(1 968.93 U·mg)(见图6)。SOD 活性与POD 的活性趋势相似,2 mg·LNAA+0.5 mg·L6-BA 处理SOD 活性最高(612.93 U·mg),其次是2 mg·LNAA+0.5 mg·L6-BA 处理SOD活性(601.99 U·mg)(见图7)。在不同的碳源类型中,碳源为蔗糖的处理CAT 活性与其他碳源类型处理差异显著(<0.05),POD 和SOD 活性与碳源为麦芽糖的处理差异不显著,与碳源为果糖的处理差异显著(<0.05)。在不同增殖培养周期中,增殖培养7 d 的处理CAT 活性最高(229.79 μmol·min·g)相较于对照处理增加了6倍。
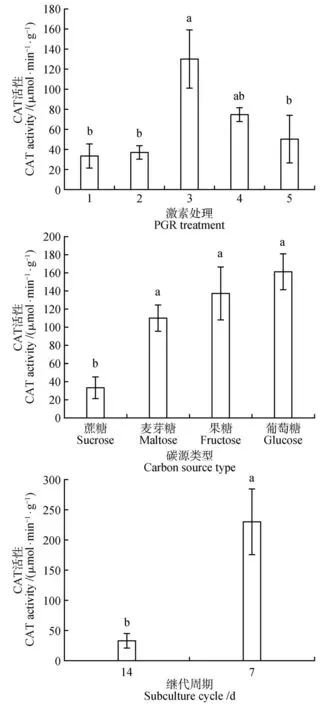
图5 激素、碳源类型和增殖培养周期对红松胚性愈伤组织中CAT活性的影响Fig.5 Effects of hormones,carbon source types and proliferation culture cycle on CAT activity in embryogenic callus of P.koraiensis
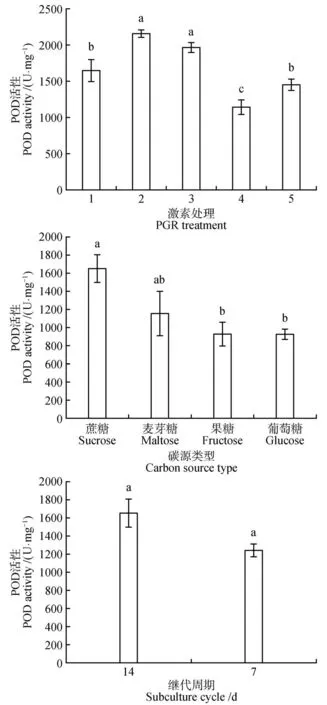
图6 激素、碳源类型和增殖培养周期对红松胚性愈伤组织中POD活性的影响Fig.6 Effects of hormones,carbon source types and proliferation culture cycle on POD activity in embryogenic callus of P.koraiensis
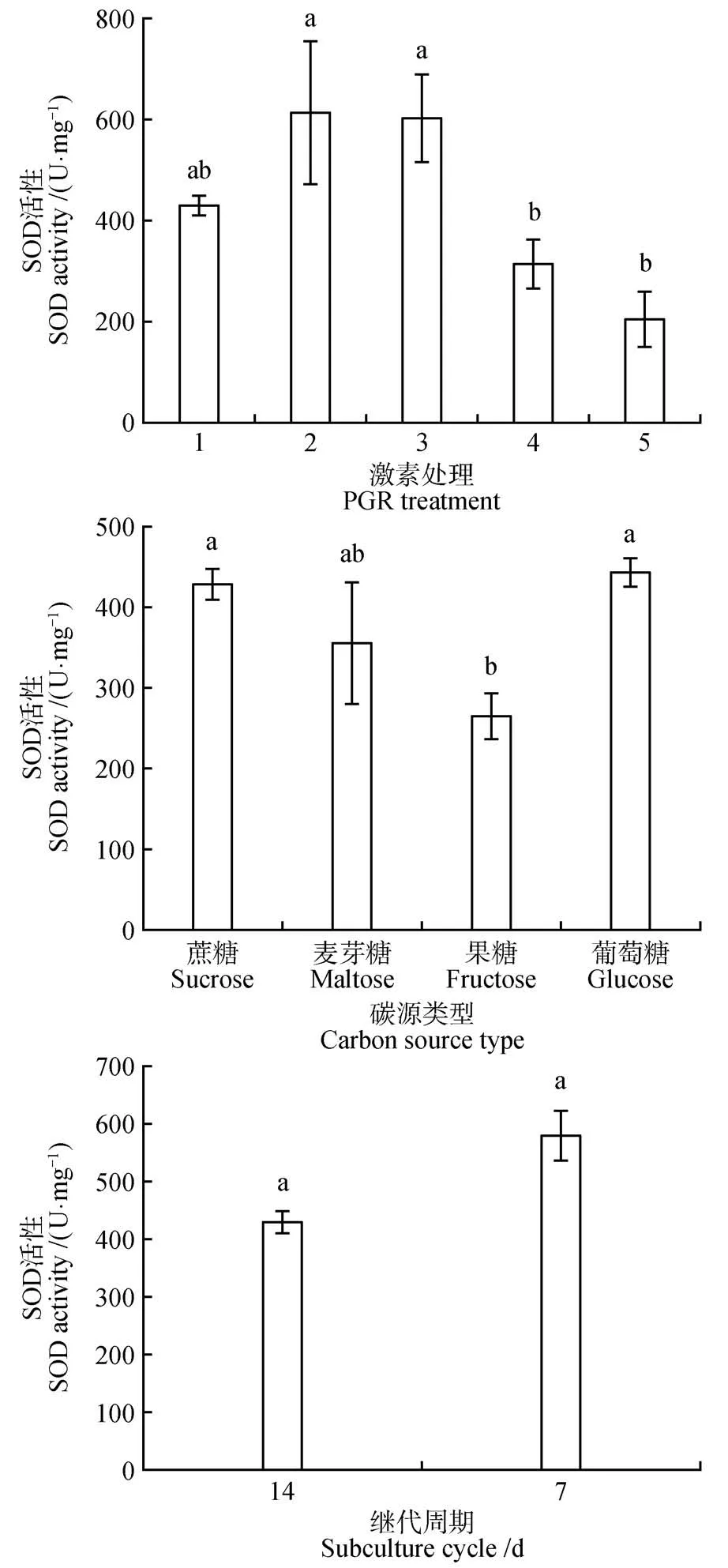
图7 激素、碳源类型和增殖培养周期对红松胚性愈伤组织中SOD活性的影响Fig.7 Effects of hormones,carbon source types and proliferation culture cycle on SOD activity in embryogenic callus of P.koraiensis
3 讨论
3.1增殖培养基中的激素浓度对体胚发生的影响
在植物组织培养过程中,植物生长素和细胞分裂素是植物离体培养所必须的激素,生长素和细胞分裂素的不同比值和类型不仅能诱导细胞分裂和生长,而且能控制细胞分化和形态发生。在长白落叶松()中研究发现胚性愈伤组织在含有2,4-D的培养基上培养时间过长时,会造成无法正常增殖的现象。在增殖阶段应该及时去掉或降低2,4-D浓度,这样既有利于胚性愈伤组织的增殖,也有利于后期更多的胚性细胞团转化为胚性胚柄细胞团。此外,有研究表明,在愈伤组织增殖阶段,用NAA 代替2,4-D 更有利于以后体胚的发生,长时间培养在含2,4-D的培养基上易造成体细胞胚成熟能力的丧失。本研究在培养基中添加1 mg·LNAA+0.25 mg·L6-BA 处理比添加2 mg·LNAA+0.5 mg·L6-BA 处理效果好,同时添加NAA 效果比添加2,4-D 的效果更好,分别印证了这些观点。我们的研究表明,在1 mg·LNAA+0.25 mg·L6-BA 处理组合下获得最多的体胚产量(166 个·g),同时该处理下愈伤组织中CAT 活性较低,POD 活性较高。有研究指出,在细胞脱分化和胚的早期发育过程中,CAT 活性低,而POD 活性高,这可能是不同时期的过氧化氢由不同酶来催化清除,使生物抗氧化系统处在一个动态平衡中。此外,Marco的研究指出,POD活性与细胞壁的松动、延伸、和交联有关,它们调控细胞壁的可塑性,影响细胞的伸长生长和细胞分裂速度。说明1 mg·LNAA+0.25 mg·L6-BA 处理POD 活性促进细胞分裂有助于体胚发生。这与詹园凤等在大蒜()体胚发生研究中发现POD 对体胚诱导起主导作用的报道相似。
3.2增殖培养基中的碳源对体胚发生的影响
碳源不仅为培养物的生长和发育代谢提供所需的能量和底物,而且调节培养基的渗透势。在植物组织培养中,蔗糖是最常用的碳源和渗透调节剂。在培养基中添加蔗糖有利于体细胞胚干物质的积累,对促进体胚发生尤其是子叶胚的形成发挥了重要的作用。在本研究中,碳源为蔗糖的处理体胚产量最高(127 个·g),同时该处理下糖和淀粉质量分数显著高于碳源为葡萄糖、麦芽糖和果糖的处理。说明碳源为蔗糖的处理中积累了体胚发生所需要的充足能量,为进一步的体胚发生奠定物质基础。与此同时,碳源为蔗糖处理中CAT 活性最低相较于其他碳源处理,而POD 活性最高。有报道指出,CAT 活性与植物抗逆性有关,与植物老化也有一定的关系,在老化的组织中其活性最强。一般认为,CAT 活性与器官代谢强弱有关,代谢强,产生的过氧化氢副产物就多,CAT 活性相对较高。而本研究中CAT 活性低说明愈伤组织还尚未老化,生长状态良好。此外,在愈伤组织细胞存在活性氧和自由基时,SOD活性会迅速增加消除此类物质避免细胞受到伤害,培养物细胞内SOD 活性在体胚发生早期迅速增加,说明SOD 可能对体胚发生有促进作用。而碳源为蔗糖的处理中SOD 活性相对较高,说明有利于细胞正常分裂和分化从而促进体胚发生。综上所述,蔗糖为红松愈伤组织体胚发生的最适碳源。
3.3增殖培养周期对体胚发生的影响
适宜的增殖培养周期,对于保持愈伤组织良好的生长状态和促进其代谢十分有利。细胞增殖及代谢物的合成都需要大量能量和营养成分,在培养基中的营养成分没有完全耗尽的情况下,及时将细胞转接到新鲜培养基中,为细胞迅速增殖创 造 条 件。秦 彩 云 等在 青 海 云 杉()的研究中发现,增殖培养周期为14 d 的愈伤组织既能保持胚性,又能获得较大的增殖量,而在海岸松()的研究中发现每周1次的继代培养,使老化效应最小化,有利于海岸松体细胞胚的早期发育。在本研究中,增殖培养周期为14 d获得的体胚产量最多(127个·g)。增殖培养周期为7 d 的处理和增殖培养周期为14 d 的处理中淀粉、可溶性糖和蛋白质质量分数差异不显著,说明其都具有体胚发生所需要的能量物质。Peng 等的研究说明丧失胚性愈伤组织细胞内可能积累过高的HO从而丧失胚性。此外,Cui 等对宁夏枸杞()体胚发生的研究中也认为,适量的H0浓度对宁夏枸杞体胚发生有促进作用,过高的HO浓度对宁夏枸杞体胚发生则有抑制作用。本研究中增殖培养7 d 的处理CAT 活性显著高于增殖培养14 d 的处理中CAT 活性,可能是HO等代谢产物增多,产生毒害作用,从而丧失胚性。由此可以看出增殖培养周期过短,不利于后期体胚的发育和成熟。与此同时,增殖培养周期过短,提高成本加大了工作量,随着增殖培养次数的增加,变异率也会提高,且体胚发生潜力逐渐下降甚至消失。
4 结论
在1 mg·LNAA 和0.25 mg·L6-BA 组合下,碳源为蔗糖,增殖培养周期为14 d 所获得的红松体胚产量最多(166个·g)。在胚性愈伤组织增殖的过程中,储藏物质为红松的体胚发生提供充足的能量来源,为其体胚发生奠定物质基础;CAT 活性过高不利于愈伤组织体胚发生,可能是HO等代谢产物增多,产生毒害作用。
