si-CECR7联合紫杉醇对肝癌耐药细胞Huh-7/ADM恶性行为的影响和机制
2022-07-25王菁王振杰伍淑娟
王菁 王振杰 伍淑娟
(1濮阳医学高等专科学校基础医学部,河南 濮阳 457000;2濮阳市人民医院风湿内科)
肝癌发病率不断增长,其治疗以手术、化疗、放疗等为主〔1〕。紫杉醇是肝癌化疗的常用药物,但肝癌细胞对紫杉醇耐药性的产生常导致肝癌化疗失败〔2〕。因此,预防和逆转肝癌耐药十分必要。猫眼综合征临界区基因(CECR)7是一种在肝癌中表达升高且与患者肿瘤等级、TNM分期及总生存率密切相关的长链非编码RNA(lncRNA)〔3〕,但其对肝癌细胞耐药性的影响及还未知。生物信息学软件预测显示,CECR7与miR-197可能存在靶向调控关系。miR-197是肝癌组织中表达降低的一个微小RNA(miRNA),其高表达的患者整体生存率高于低表达患者,miR-197可作为肝癌早期诊断的生物标志物〔4〕;同时,miR-197可通过靶向抑制锌指蛋白(ZIK1)的表达抑制肝癌细胞侵袭〔5〕。但是,miR-197对肝癌细胞耐药性的影响也还未知。本研究主要探究了CECR7小干扰RNA(si-CECR7)联合紫杉醇对肝癌紫杉醇耐药Huh-7细胞(Huh-7/ADM)恶性行为的影响及其能否通过调控miR-197发挥作用。
1 材料与方法
1.1细胞和试剂 肝癌细胞Huh-7及紫杉醇耐药细胞Huh-7/ADM(中国科学院上海细胞库);紫杉醇(北京协和药厂)每支30 mg/ml;胎牛血清(浙江天杭);双荧光素酶活性检测试剂盒、四甲基噻唑蓝(MTT)、二喹啉甲酸(BCA)蛋白检测试剂盒和膜联蛋白(Annexin)V-异硫氰酸荧光(FITC)/碘化丙啶(PI)试剂盒(北京索莱宝);PCR实验相关试剂盒(大连宝生物);引物序列、miR-144-3p模拟物(mimics)及模拟对照序列(miR-NC)、si-CECR7及小干扰RNA阴性序列(si-NC)、miR-197抑制剂(anti-miR-197)及阴性序列(anti-miR-NC)、野生型(WT)和突变型(MUT)的CECR7荧光素酶报告载体(上海吉凯基因公司);LipofectamineTM2000试剂盒(美国Invitrogen公司);蛋白质印迹实验相关抗体(中国Abcam公司)。
1.2临床资料 56例肝癌选自2016年3月至2018年11月于濮阳市人民医院确诊并行手术治疗的患者,男39例,女17例,平均年龄(62.31±6.49)岁。术中留取患者癌组织和癌旁组织,液氮保存。纳入标准:术前均未接受治疗。研究符合《赫尔辛基宣言》原则。
1.3方法
1.3.1细胞培养 Huh-7和Huh-7/ADM细胞均用完全培养基(含10%胎牛血清的RPMI1640培养基)培养。
1.3.2MTT实验 于96孔板中接种Huh-7和Huh-7/ADM细胞,接种个数均为5.0×104个/孔。培养12 h后,均用含12.5、25.0、50.0、100.0 μg/ml〔6〕紫杉醇的培养基干预48 h。然后加5 g/L的MTT(20 μl/孔),孵育4 h。弃培养基,加二甲基亚砜(150 μL/孔),振荡均匀。将96孔板放置在酶标仪卡槽中,490 nm测吸光度(A)值。细胞抑制率(%)=(A对照组-A实验组)/A对照组×100%。计算紫杉醇对每组细胞的半数抑制浓度(IC50)。
1.3.3实时荧光定量PCR(qRT-PCR)检测CECR7和miR-197表达 ①在液氮保护下,用Trizol试剂提取组织样本中总RNA。②于6孔板中接种Huh-7和Huh-7/ADM细胞,接种个数均为1.0×105个/孔,培养24 h后,收集细胞,用Trizol试剂提取组织样本中总RNA。将组织和细胞中RNA均逆转录为cDNA,进行扩增。引物序列:CECR7上游5′-GAGATGGATAGCGCTCGA-3′,下游5′-AACCGTGACGAAC GTTCC-3′;miR-197上游5′-CCGTACGGGCAATGCAAC-3′,下游5′-GTGCCAACGTACGGTCGAG-3′;GAPDH上游5′-GACCCCCATGACCTCAAC-3′,下游5′-CTCCTCAGTGTGGTGAAGA-3′;U6上游5′-GAGTGCGACTGACGTAGTACCG-3′,下游5′-CGTGTCCAAGGCCTGTTCACGCG-3′。2-△△Ct法计算CECR7相对GAPDH、miR-197相对U6的表达。
1.3.4细胞转染 接种Huh-7/ADM细胞至6孔板中,接种个数为1.0×105个/孔。培养24 h后,将LipofectamineTM2000试剂分别与si-CECR7、si-NC、si-CECR7和anti-miR-NC、si-CECR7和anti-miR-197混合均匀,加至6孔板中(100 μl/孔),孵育细胞12 h。qRT-PCR检测CECR7或miR-197表达,验证转染效果后,用于后续实验。
1.3.5细胞存活率检测 MTT检测细胞存活率。将转染si-CECR7或si-NC、共转染si-CECR7和anti-miR-NC、si-CECR7和anti-miR-197的Huh-7/ADM细胞均接种至96孔板中,接种个数均为5.0×104个/孔。培养12 h后,均用含12.5 μg/ml紫杉醇的培养基干预48 h,并分别记为紫杉醇+si-CECR7组、紫杉醇+si-NC组、紫杉醇+si-CECR7+anti-miR-NC组、紫杉醇+si-CECR7+anti-miR-197组。同时接种未转染的Huh-7/ADM细胞,分别用含0.0(对照组)、12.5 μg/ml(紫杉醇组)紫杉醇的培养基干预48 h。然后按照上述1.3.2测定各孔A值。存活率(%)=A实验组/A对照组×100%。
1.3.6检测细胞克隆形成能力检测 克隆实验检测细胞克隆形成能力。将未转染或转染后的Huh-7/ADM细胞均接种至6孔板中,接种个数均为5.0×104个/孔。培养12 h后,均按照上述1.3.5分组处理。培养14 d后,弃培养基。显微镜观察,统计超过50个细胞的克隆数。
1.3.7细胞凋亡检测 流式细胞仪检测细胞凋亡。将未转染或转染后Huh-7/ADM细胞,均接种至6孔板中,接种个数均为5.0×104个/孔。培养12 h后,均按照上述1.3.5分组处理。培养48 h后,收集各组细胞,利用Annexin V-FITC/PI试剂盒,上流式细胞仪检测细胞凋亡。
1.3.8细胞迁移和侵袭检测 Transwell检测细胞迁移和侵袭。将未转染或转染后Huh-7/ADM细胞,均接种至6孔板中,接种个数均为5.0×104个/孔。培养12 h后,均按照上述1.3.5分组处理。培养48 h后,收集各组细胞。迁移:将收集的各组细胞均接种至Transwell上室,接种个数均为2.5×104个/孔。另取500 μl完全培养基加至下室。培养24 h后,弃培养基。将下室细胞进行固定和染色。显微镜观察并计数。侵袭:将基质胶铺于Transwell上室,干燥后,再接种细胞,后续操作同迁移实验。
1.3.9细胞周期蛋白(Cyclin)D1、p21、B淋巴细胞瘤(Bcl)-2、Bcl-2相关X蛋白(Bax)、上皮型钙黏蛋白(E-cadherin)和基质金属蛋白酶(MMP)-2蛋白表达检测 Western印迹法检测CyclinD1、p21、Bcl-2、Bax、E-cadherin和MMP-2蛋白表达。用RIPA试剂提取各组细胞中总蛋白,BCA法测定蛋白含量。利用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)实验分离总蛋白,并转至聚偏氟乙烯(PVDF)膜,于5 %脱脂牛奶中封闭1 h。然后将PVDF膜分别置于CyclinD1、p21、Bcl-2、Bax、E-cadherin和MMP-2一抗孵育液中,放置于4℃冰箱中孵育过夜。洗膜后,再置于山羊抗兔二抗中,置于37℃摇床中孵育2 h。滴加显影液显影,曝光拍照。
1.3.10CECR7与miR-197靶向关系验证 双荧光素酶报告基因实验验证CECR7与miR-197靶向关系。接种Huh-7/ADM细胞至6孔板中,接种个数为1.0×105个/孔。培养24 h后,将LipofectamineTM2000试剂分别与miR-NC(或miR-197mimics)和WT-CECR7、miR-NC(或miR-197mimics)和MUT-CECR7混合均匀,加至6孔板中(100 μl/孔),孵育细胞12 h。收集细胞并裂解,将裂解液3500 r/min离心10 min。取100 μl 1×荧光素酶反应工作液,加20 μl离心后的上清液,混合均匀,检测萤火虫荧光强度。再加100 μl 1×海肾荧光素酶反应工作液,混合均匀,检测海肾荧光强度。以萤火虫荧光强度与海肾荧光强度的比值表示细胞荧光素酶活性。
1.4统计学分析 采用SPSS22.0软件进行独立样本t检验、单因素方差分析、SNK-q检验。
2 结 果
2.1紫杉醇对Huh-7/ADM细胞抑制率的影响 与对照组比较,紫杉醇组Huh-7、Huh-7/ADM细胞抑制率显著升高,且呈浓度依赖性(P<0.05)。紫杉醇对Huh-7、Huh-7/ADM细胞的IC50值分别为24.10 μg/ml、161.00 μg/ml,后者约是前者的6.7倍。见表1。

表1 不同浓度的紫杉醇对Huh-7/ADM细胞抑制率的影响
2.2肝癌组织和细胞系中CECR7表达水平 与癌旁组织(1.01±0.10)比较,肝癌组织中CECR7表达水平显著升高(1.56±0.16,P<0.05)。与Huh-7细胞(1.01±0.10)比较,Huh-7/ADM细胞中CECR7表达水平显著升高(2.09±0.21,P<0.05)。
2.3si-CECR7联合紫杉醇对Huh-7/ADM细胞增殖和凋亡的影响 CECR7在转染si-CECR7的细胞中的表达量显著低于转染si-NC的细胞(0.25±0.04 vs 1.03±0.10,t=21.726,P<0.05)。对照组与紫杉醇组各检测指标比较差异均无统计学意义(P>0.05)。紫杉醇+si-CECR7组Huh-7/ADM细胞存活率、克隆数、CyclinD1和Bcl-2蛋白表达均显著低于紫杉醇+si-NC组(P<0.05),而凋亡率、P21和Bax蛋白表达均显著高于紫杉醇+si-NC组(P<0.05)。见图1、图2、表2、图3。
2.4si-CECR7联合紫杉醇对Huh-7/ADM细胞迁移和侵袭的影响 对照组与紫杉醇组各检测指标比较差异均无统计学意义(P>0.05)。紫杉醇+si-CECR7组Huh-7/ADM细胞迁移数、侵袭数和MMP-2蛋白表达均显著低于紫杉醇+si-NC组(P<0.05),E-cadherin蛋白表达显著高于紫杉醇+si-NC组(P<0.05)。见图4、表3、图5。

1~4:对照组,紫杉醇组,紫杉醇+si-NC组,紫杉醇+si-CECR7组图1 si-CECR7联合紫杉醇对Huh-7/ADM细胞增殖和凋亡相关蛋白表达的影响

图2 si-CECR7联合紫杉醇对Huh-7/ADM细胞凋亡的影响
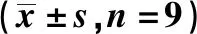
表2 si-CECR7联合紫杉醇对Huh-7/ADM细胞增殖和凋亡的影响
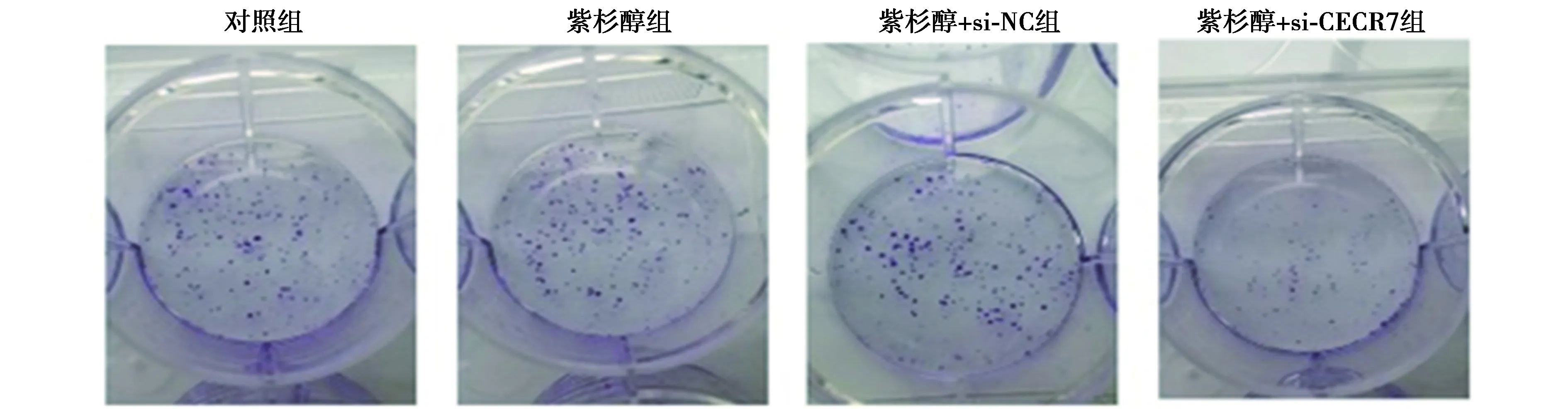
图3 si-CECR7联合紫杉醇对Huh-7/ADM细胞克隆形成的影响(结晶紫染色,×50)
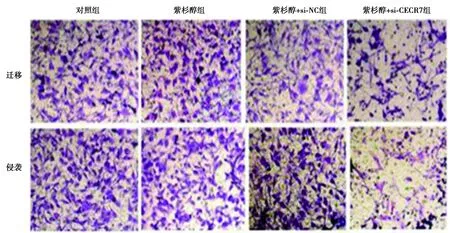
图4 si-CECR7联合紫杉醇对Huh-7/ADM细胞迁移和侵袭的影响(结晶紫染色,×200)
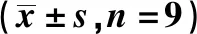
表3 si-CECR7联合紫杉醇对Huh-7/ADM细胞迁移和侵袭的影响

1~4:对照组,紫杉醇组,紫杉醇+si-NC组,紫杉醇+si-CECR7组图5 si-CECR7联合紫杉醇对Huh-7/ADM细胞迁移和侵袭蛋白表达的影响
2.5CECR7靶向调控miR-197表达 CECR7与miR-197的结合位点见图6。共转染miR-NC与WT-CECR7、miR-197与WT-CECR7的Huh-7/ADM细胞荧光素酶活性分别为0.93±0.09、0.20±0.02,两组间比较差异有统计学意义(t=23.754,P<0.05);共转染miR-NC与MUT-CECR7、miR-197与MUT-CECR7的Huh-7/ADM细胞荧光素酶活性分别为0.96±0.09、1.01±0.10,两组间比较差异无统计学意义(t=1.115,P=0.533),说明CECR7靶向结合miR-197。此外,miR-197在转染si-CECR7的Huh-7/ADM细胞中的表达量为2.46±0.15,显著高于转染si-NC的Huh-7/ADM细胞中的表达量(1.01±0.10,t=24.129,P<0.05),说明CECR7负调控miR-197表达。
2.6anti-miR-197逆转si-CECR7联合紫杉醇对Huh-7/ADM细胞增殖、凋亡、迁移和侵袭的影响 miR-197在共转染si-CECR7与anti-miR-197的细胞中的表达量显著低于共转染si-CECR7与anti-miR-NC的细胞(0.36±0.04 vs 1.00±0.10,t=17.827,P<0.05)。紫杉醇+si-CECR7+anti-miR-197组Huh-7/ADM细胞存活率、克隆数、迁移数、侵袭数及CyclinD1、Bcl-2和MMP-2蛋白表达均显著高于紫杉醇+si-CECR7+anti-miR-NC组(P<0.05),而凋亡率、P21、E-cadherin和Bax蛋白表达均显著低于紫杉醇+si-CECR7+anti-miR-NC组(P<0.05)。见图7~9、表4和表5、图10。

图6 CECR7与miR-197的结合位点
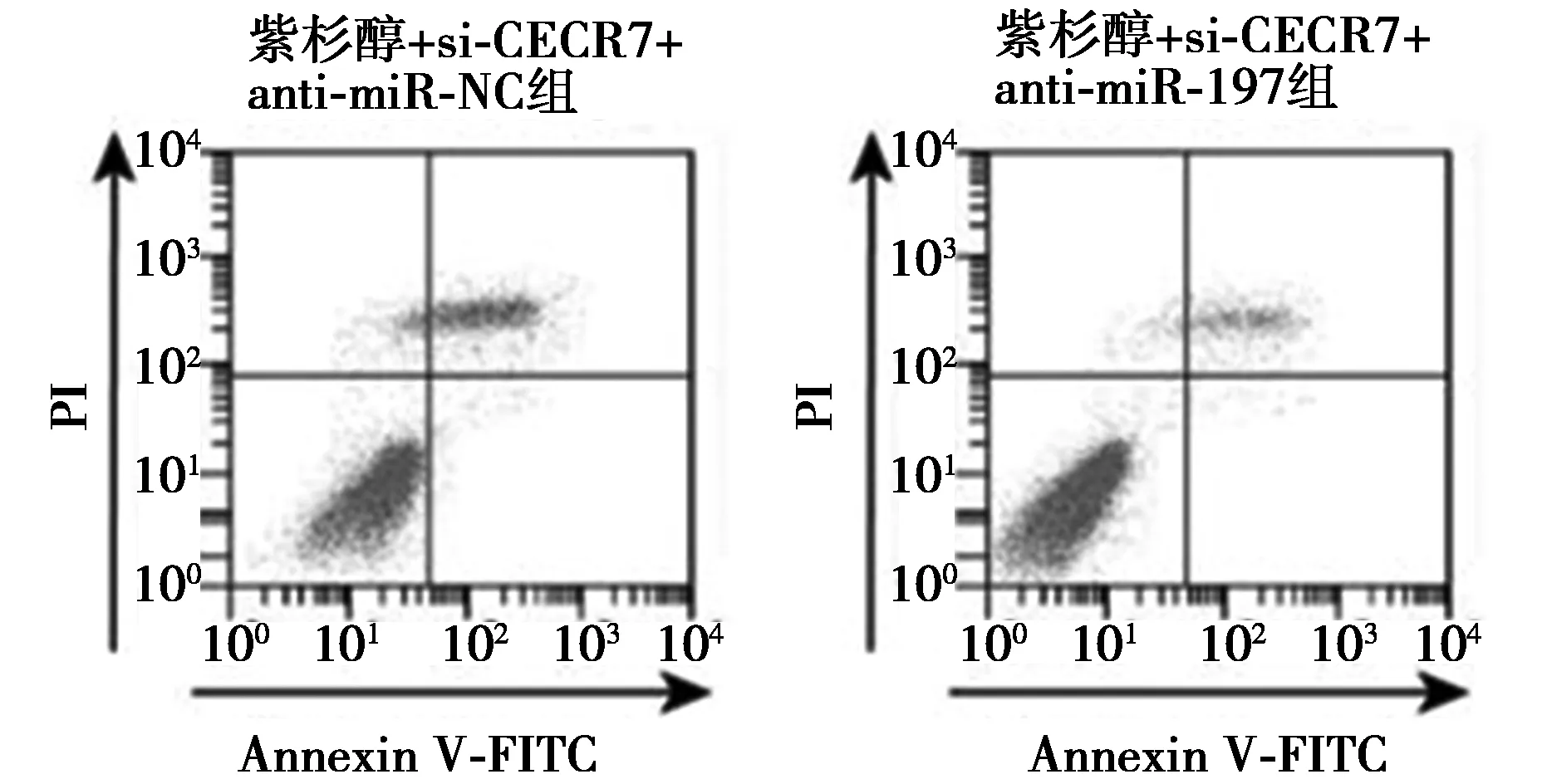
图7 anti-miR-197逆转si-CECR7联合紫杉醇对Huh-7/ADM细胞凋亡的影响

图8 anti-miR-197逆转si-CECR7联合紫杉醇对Huh-7/ADM细胞克隆形成的影响(结晶紫染色,×50)

图9 anti-miR-197逆转si-CECR7联合紫杉醇对Huh-7/ADM细胞迁移、侵袭的影响(结晶紫染色,×200)
表4 anti-miR-197逆转si-CECR7联合紫杉醇对Huh-7/ADM细胞增殖、迁移、侵袭的影响
表5 anti-miR-197逆转si-CECR7联合紫杉醇对Huh-7/ADM细胞凋亡的影响
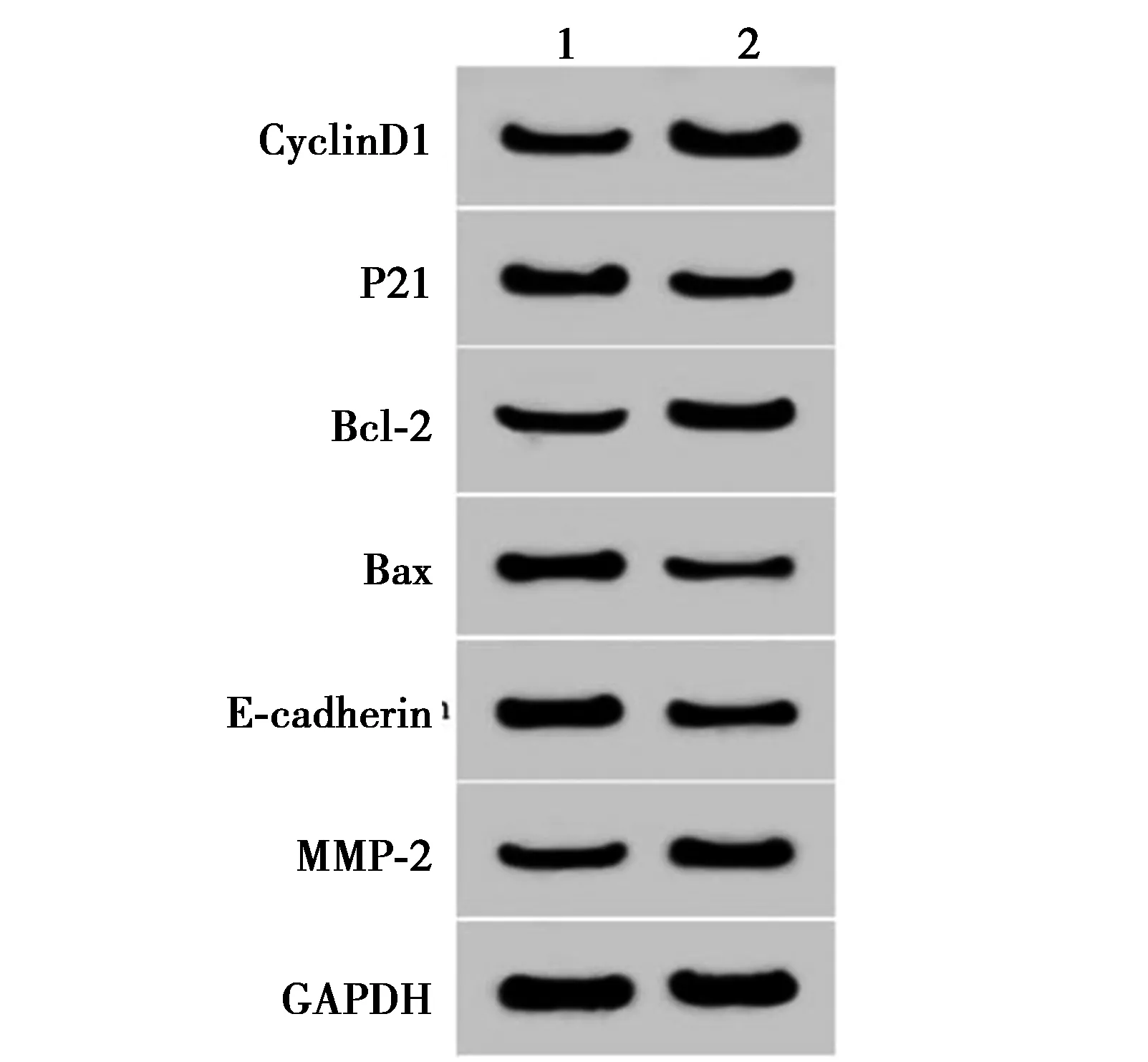
1,2:紫杉醇+si-CECR7+anti-miR-NC组,紫杉醇+si-CECR7+anti-miR-197组图10 anti-miR-197逆转si-CECR7联合紫杉醇对Huh-7/ADM细胞增殖、凋亡、迁移、侵袭蛋白表达的影响
3 讨 论
化疗是肝癌常用的治疗方法,但肝癌细胞易对紫杉醇等化疗药物产生耐药性,导致化疗失败〔7,8〕。因此,降低肝癌细胞耐药性对于提高肝癌化疗疗效具有积极意义。研究显示,lncRNA参与调控肿瘤细胞的耐药性〔9〕,探讨影响肿瘤细胞耐药性的lncRNA及其作用机制,可为降低肿瘤耐药性提供分子靶点。例如,敲减lncRNA SNHG7降低卵巢癌细胞对紫杉醇的耐药性〔10〕;lncRNA OTU结构域蛋白酶6B反义RNA1(OTUD6B-AS1)通过调节miR-26a-5p/异黏蛋白(MTDH)通路促进三阴性乳腺癌细胞对紫杉醇的耐药性,通过敲减lncRNA OTUD6B-AS1可起到改善三阴性乳腺癌紫杉醇耐药的作用〔11〕;lncRNA尿路上皮癌相关基因(AUCA)1在结肠癌中表达上调,敲减lncRNAUCA1可通过下调糖酵解酶己糖激酶2和乳酸脱氢酶A的表达增强结肠癌细胞对紫杉醇的敏感性〔12〕。
作为一种lncRNA,CECR7参与多种肿瘤的发展进程。王向辉等〔13〕研究显示,食管癌细胞中CECR7表达升高,沉默CECR7可能通过抑制细胞周期蛋白依赖激酶4、CyclinD1和p21活化蛋白激酶2的表达抑制食管癌细胞的恶性行为,CECR7是食管癌治疗的潜在新靶点。本研究主要探究了CECR7对肝癌细胞紫杉醇脑药性的影响,结果显示,CECR7在肝癌耐药细胞株Huh-7/ADM中的表达水平高于肝癌细胞Huh-7,转染si-CECR7联合紫杉醇降低了Huh-7/ADM细胞的增殖、迁移及侵袭性,促进了细胞凋亡,同时降低了细胞中与增殖、凋亡、迁移和侵袭相关的蛋白CyclinD1、Bcl-2、E-cadherin和MMP-2的表达,而促进了P21和Bax蛋白表达,与相关报道结果一致〔14,15〕,说明si-CECR7可降低Huh-7/ADM细胞对紫杉醇的耐药性,CECR7有可能成为改善肝癌细胞对紫杉醇耐药的分子靶点。
miR-197是胃癌中下调的miRNA,其可通过靶向抑制CXCR6的表达阻碍胃癌的发展进程〔16〕;子宫内膜癌中miR-197表达下调,上调miR-197可通过靶向抑制CTNND1/Wnt/β-catenin轴降低子宫内膜癌细胞的恶性行为〔17〕。Wang等〔18〕研究显示,miR-197可在体外和体内显著抑制肝细胞癌细胞的生长,可能是干扰肝细胞癌中白细胞介素6/信号转导及转录激活因子3炎症途径的潜在治疗靶标。本研究利用恢复实验发现,转染anti-miR-197逆转了si-CECR7联合紫杉醇对紫杉醇耐药肝癌细胞恶性行为的影响,这提示CECR7可能通过靶向负调控miR-197来降低紫杉醇耐药肝癌细胞对紫杉醇的耐药性。
综上所述,CECR7在肝癌耐药细胞Huh-7/ADM中呈高表达,si-CECR7联合紫杉醇抑制紫杉醇耐药肝癌细胞的恶性行为,这可能与转染si-CECR7引起细胞中miR-197的表达上调有关,但其具体作用的miR-197的靶基因还有待进一步探究,CECR7有可能成为改善紫杉醇耐药肝癌细胞对紫杉醇耐药性的分子靶点。
