毛地黄鼠尾草在不同授粉条件下的繁殖成功与花性状选择
2022-07-08张勃孙淑范
张勃,孙淑范
(甘肃农业大学草业学院,草业生态系统教育部重点实验室,甘肃 兰州 730070)
普遍认为,被子植物花的多样性进化在很大程度上是由传粉者介导的自然选择所驱动[1-3]。近年来,大量的进化生态学研究显示,植物花性状选择在自然种群普遍发生[4],传粉者类群的转变在植物物种或种群分化中发挥着关键作用[5-6]。但是,目前有关植物传粉者介导选择的定量估测研究相对较少[7-8],从而限制了对传粉者作为选择媒介(selective agent)促进花表型进化过程的理解。
花表型选择发生,需要同时满足两个基本条件:首先,植物的花性状表达(trait expression)在自然种群中存在个体间的变异性[9];其次,不同的花性状能通过与选择媒介(如传粉昆虫)的互作引起个体间繁殖适合度(reproductive fitness)的变异[10]。也就是说,当种群内不同个体因为花性状变异而获得不同的传粉机会(或效率),从而导致个体间繁殖适合度存在差异时,传粉者介导的选择才能够发生[5]。植物通常有两类花性状会通过影响昆虫传粉而受到传粉者选择[11-12]。一类是花吸引(传粉昆虫)相关性状,如花大小、数量[13]和花报酬[14]等,这类性状可通过影响昆虫访花频率影响植物个体的传粉机会,进而影响其繁殖成功。另一类是植物的花部结构性状[15],特别是花药和柱头的相对位置以及花冠结构等,这类性状可通过与传粉者的机械互作影响植物的传粉效率和交配成功[16],从而影响其繁殖适合度。
鼠尾草(Salvia)是唇形科(Lamiaceae)最大的一个属,被认为是研究植物花适应性进化的理想类群[17]。该属绝大多数物种具有高度特化的杠杆状雄蕊结构(lever-like stamens)和背部传粉机制[18]。有学者认为,这种特殊花结构及其与传粉者形成的传粉互作系统是激发该属物种适应性分化的“关键性状”(a key innovation)[19];因此,该植物的花性状进化和选择机制成为学者普遍关注的科学问题。针对该属植物的花表型进化,前人已在其系统发育[20]、传粉生态[21-22]、花结构功能与适应性[23-24]以及性系统的多样性分化[18,25]等方面展开了深入研究。其中,Zhang 等[17]通过对毛地黄鼠尾草(Salvia digitaloides)的花适应性研究发现,该物种的花产量及雄蕊杠杆相关花性状受到强烈的自然选择,但是其选择机理(包括选择媒介)仍不清楚。基于此,本研究通过人工辅助授粉创造不同的传粉环境,比较了该植物在不同花粉供给条件下的繁殖成功和花性状选择,探讨了花性状在当前自然种群的适应性及其选择机理。具体关注如下两个问题:1)在自然种群,毛地黄鼠尾草的繁殖是否受到花粉限制?2)该植物的哪些花性状受传粉者介导的选择,其选择强度和选择模式是什么?
1 材料与方法
1.1 试验地点和材料
本试验于 2018年 5-6月进行,试验地位于云南省丽江市甘海子(27°0′54″N,100°14′41″E),海拔 2419 m;毛地黄鼠尾草是该地区常见的优势草本植物。该物种属于唇形科弧隔鼠尾草亚属(subgenusSalvia),主要分布于我国云南西北部和四川西南部。本研究种群特征为:花乳白色,花冠口及唇瓣有红色或紫红色斑点;花期为4月底到5月中旬,单花有4个胚珠,小坚果呈灰黑色。
1.2 试验设计与方法
初花期,在毛地黄鼠尾草种群随机选择150个开花状态相同的单株,每株同时标记大小基本一致且尚未开放的2个花序。其中,一个花序作对照,使其自然授粉;另一个花序作处理,进行人工辅助授粉。待所标记的花序开花后,对处理花序上每朵新开放的小花进行1~2 次人工辅助授粉,直至所有小花开败萎蔫。授粉时,用柔软毛刷蘸取采自周围未标记花序的混合花粉,轻触花柱头即可。
1.2.1 花性状测量 待标记植株处于盛花期时,在每个个体上选择4~5 朵花,用电子游标卡尺(精度为0.01 mm)测量其花冠长、花冠口高度和宽度、柱头高度以及柱头探出花冠距离(即柱头探出距离)5个花部结构性状。具体测量方法参照 Zhang 等[17,25]的方法。
1.2.2 适合度估测 待毛地黄鼠尾草种子成熟后(花后15 d 左右),采集所有标记花序,统计其坐果数和结籽数;同时,根据花序上留存的花柄数量统计每个花序的花数量,并在花序基部测量每个花序的茎粗,以此作为个体大小的测度。用每个处理花序的坐果率(坐果数/花数量)和结籽率(种子数/花数量)估测个体的繁殖效率,用结籽数(种子数)估测其繁殖适合度。
1.2.3 花粉限制程度估测个体的花粉限制程度用个体在花粉限制下的繁殖效率折损来估测。坐果率(或结籽率)折损的计算公式如下,以此表示该个体在坐果和结籽水平所受的花粉限制程度。

1.2.4 选择梯度的估测 选择梯度用来度量性状的直接选择作用强度[26]。参照Lande 等[27]的方法,通过建立目标性状与个体适合度的回归模型估测花性状的选择梯度,其完整的估测模型[4]如下:

1.3 数据统计分析
人工辅助授粉(处理)和自然授粉(对照)个体的坐果率、单花结籽率以及种子数的差异性均采用广义线性模型(generalized linear model,GLM)进行分析。采用具有拟二项(quasi-binomial)分布族的广义线性模型(logistic链接函数)分析坐果率与单花结籽率的差异显著性;采用具有负二项式分布族的广义线性模型(log 链接函数)分析个体适合度(种子数)的差异显著性。
个体花性状与其坐果率和结籽率的关系用广义线性模型(二项分布,logistic 链接函数)进行多元回归分析。模型中,因变量为个体的坐果率或结籽率,自变量为各花性状值,同时加入“茎粗”作为协变量,以消除母株影响。花性状与个体坐果率和结籽率折损的线性关系,采用一般线性模型进行分析。分析前,对各模型解释变量进行标准化处理:(个体性状值-种群平均值)/标准差,使其平均数为0,方差为1。
性状选择梯度用一般线性模型(linear model,LM)通过多元回归分析进行估测,因变量为个体的相对适合度(个体适合度/种群平均适合度),自变量为标准化的性状值。根据Lande 等[27]的方法,线性(定向)选择梯度通过建立简单线性模型进行估测,即模型中解释变量只包括各性状的一次项;非线性选择和相关选择梯度通过建立全模型进行分析,模型的解释变量包括各性状的一次项、二次项和互作项。本研究中,所有选择模型均未检测到显著的非线性选择,因此,在结果部分未展示相关数据。最后,对模型中显著性状的选择梯度运用变量添加图(the added-variable plots)进行回归诊断。以上所有数据统计均在R 软件(R 3.5.1)中运行完成,采用Sigmaplot 和Origin 软件作图。
2 结果与分析
2.1 毛地黄鼠尾草在不同授粉条件下的繁殖成功
毛地黄鼠尾草个体的坐果率,单花结籽率以及种子数在自然授粉和人工辅助授粉条件下皆存在显著差异(图1)。在人工辅助授粉条件下,个体的坐果率与单花结籽率分别为38.61%和0.837,在自然授粉条件下分别为26.20%和0.535,二者在处理与对照间均存在显著差异(P<0.001)。在自然授粉条件下,个体的平均种子数为20.48,显著低于人工辅助授粉条件(32.20,P<0.001)。

图1 毛地黄鼠尾草在不同授粉条件下的坐果率、单花结籽率和种子数Fig.1 Fruit set,seed set per flower and seed number of S. digitaloides individuals under different conditions of pollination
2.2 毛地黄鼠尾草个体的繁殖效率及其在花粉限制条件下的折损与花性状的关系
在不同授粉条件下,毛地黄鼠尾草个体的坐果率和结籽率与花性状的关系存在明显的差异(表1)。自然授粉条件下,个体的坐果率和结籽率均与花柱头高度存在显著的负相关关系,其偏回归系数分别为-0.173(P=0.017)与-0.158(P=0.029);花冠口大小与个体的坐果率显著正相关(P=0.026),而与结籽率的关系不显著(P=0.088)。在人工辅助授粉条件下,个体的坐果率和结籽率均与花数量存在显著的负相关关系,偏回归系数分别为-0.171(P=0.024)和-0.203(P=0.006);另外,个体的结籽率与花柱头探出花冠长度也存在显著的负相关关系,其偏回归系数为-0.227(P=0.031)。在自然授粉条件下,该植物因花粉限制导致的结籽率折损与花柱头探出花冠长度和花数量均存在显著的负相关关系,其偏回归系数分别为-0.091(P=0.041)和-0.119(P=0.003);个体的坐果率折损与花数量之间存在显著的负相关性,其回归系数为-0.040(P=0.018)。

表1 毛地黄鼠尾草的坐果率和结籽率及其在花粉限制条件下的折损与花性状的回归关系Table 1 The regression relationship of S. digitaloides floral traits with fruit set,seed set,and their discounts resulting from pollen limitation
2.3 不同授粉条件下毛地黄鼠尾草花性状的选择
毛地黄鼠尾草的花性状在自然授粉与人工辅助授粉条件下的选择存在显著差异(表2 和图2)。花数量在自然授粉与人工辅助授粉条件下皆受到显著的正定向选择,选择梯度分别为0.254(图2B,P<0.001)和0.120(图2F,P=0.040)。柱头高度在自然授粉条件下受到了显著的定向选择,其选择梯度为-0.166(图2A,P=0.014),而在人工辅助授粉条件下,其选择梯度不显著(P=0.093)。相反,柱头探出花冠长度在人工辅助授粉条件下受到了显著的负定向选择,选择梯度为-0.176(图2E,P=0.028),而在自然授粉条件下其选择梯度不显著。自然授粉条件下,该植物的花冠长和柱头高度以及冠口大小和柱头探出花冠长度两个组合性状受到了显著的相关选择,其选择强度分别为0.115(图2C,P=0.025)和-0.119(图2D,P=0.025);在人工辅助授粉条件下,对花性状组合未检测到显著的相关选择。
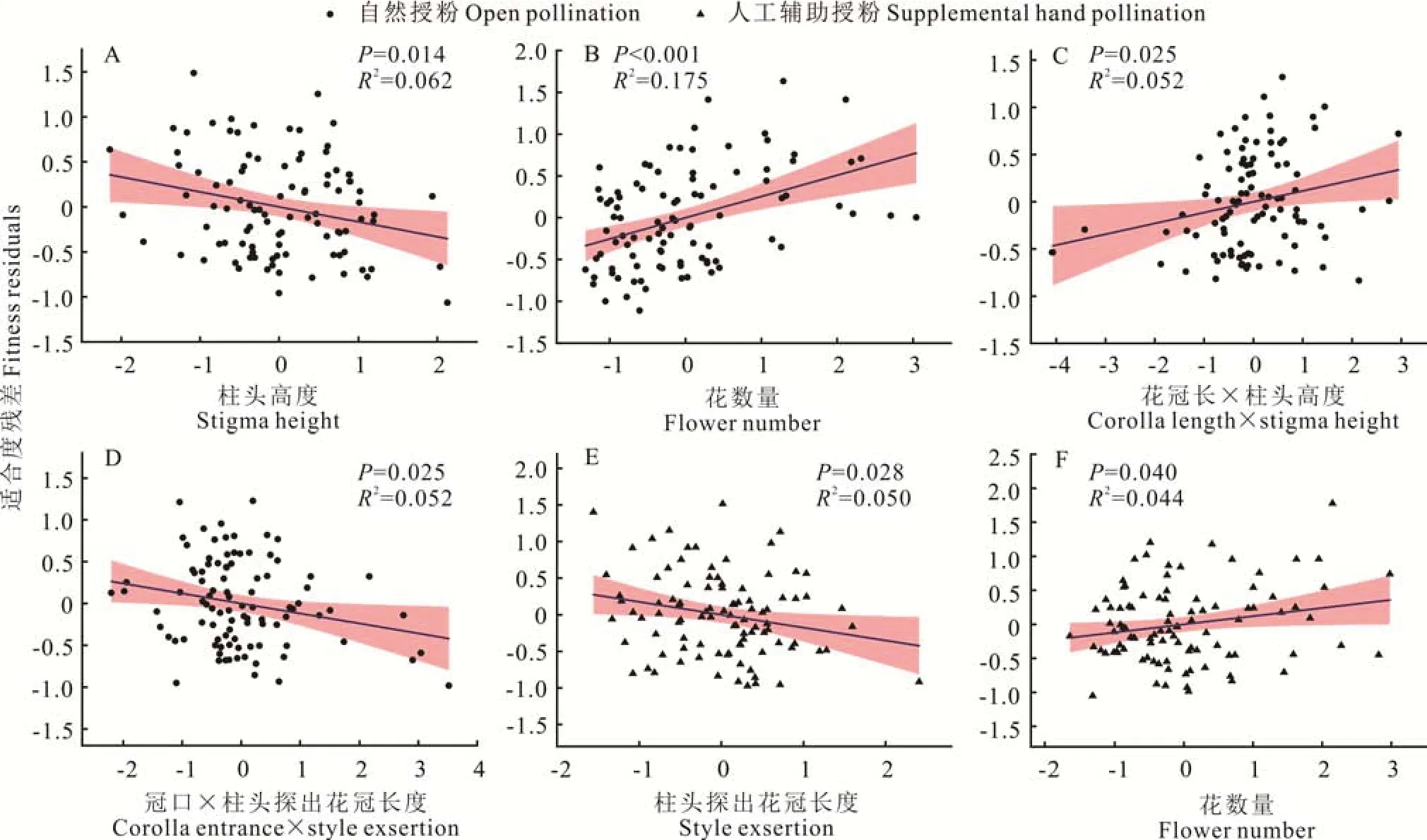
图2 毛地黄鼠尾草花性状在不同授粉条件下显著的选择梯度Fig.2 Significant selection gradients for floral traits of S. digitaloides under different pollination conditions
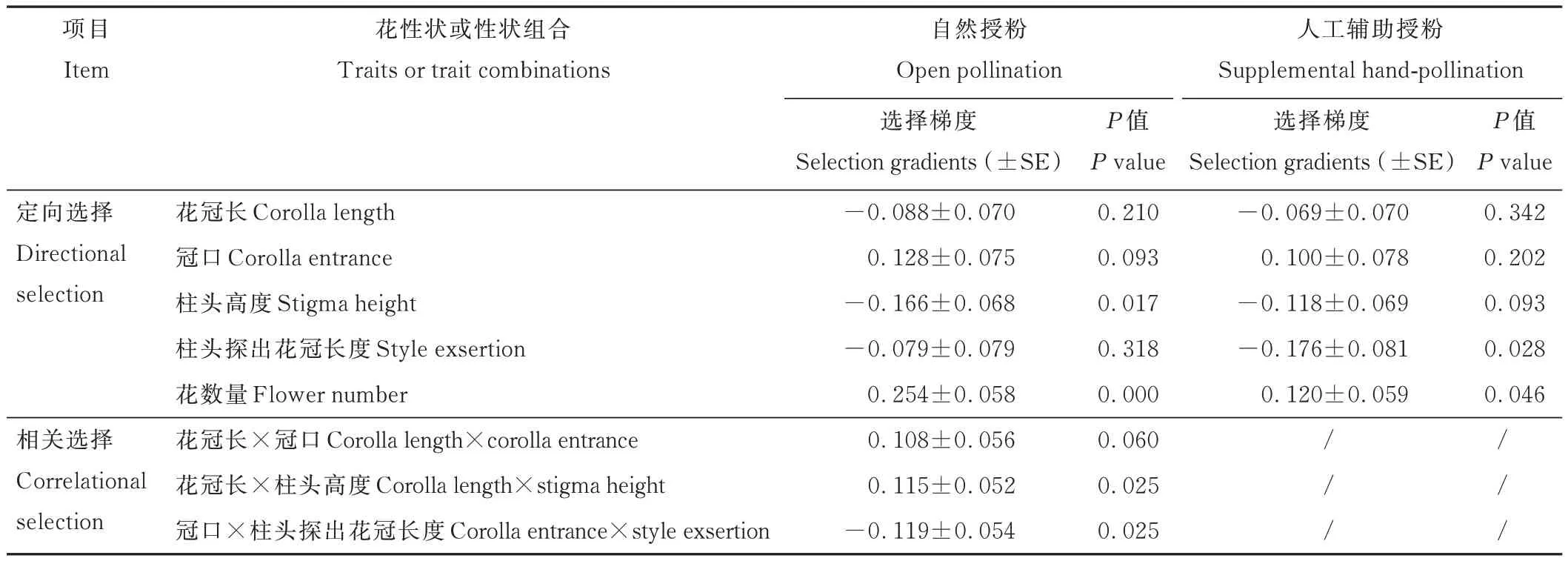
表2 毛地黄鼠尾草种群在不同授粉条件下的花性状选择梯度Table 2 Selection gradients for floral traits of S.digitaloides under different pollination conditions
3 讨论
在自然种群中,植物的繁殖成功普遍受到花粉限制或资源供给不足的影响[28-29]。表型选择理论认为,当植物的繁殖存在花粉限制,且种群内不同个体的花粉限制程度存在差异时,传粉者介导的选择才能够发生[5]。本研究结果显示,人工辅助授粉能显著提高毛地黄鼠尾草个体的雌性适合度,表明该植物的繁殖存在明显的花粉限制。另外,在自然授粉条件下,该植物不同个体因花粉限制导致的繁殖效率(即坐果率和结籽率)折损与个体的花数量之间存在显著的负相关性,说明花数量越大的个体,其花粉限制程度相对越低。相应地,该植物的花数量在自然授粉条件下受到了强烈的定向选择,通过人工补充授粉后,其选择强度显著降低。这一结果表明,在花粉限制条件下,该植物的花产量性状受到了强烈的传粉者介导的选择。许多研究表明,植物的花数量不仅会通过传粉者受到间接选择,而且在很大程度上会因为决定植物个体的繁殖交配机会(reproductive capacity for mating opportunities)而受到直接选择[30-31]。本研究中,该植物的花产量性状在补充授粉条件下仍检测到显著的选择梯度,在一定程度上反映了花数量通过影响个体的繁殖交配机会所受到的选择作用。根据Bateman 法则,植物在授粉充足时,其雌性适合度(种子生产)主要受资源限制的影响[32]。本研究中,毛地黄鼠尾草在辅助授粉条件下的坐果率和结籽率与个体的花数量之间皆存在显著的负相关关系,表明在授粉充足时该植物个体的花产量与种子生产之间存在明显的资源权衡(trade-off)。因此,其花产量性状选择随传粉环境变化会受到不同程度的资源限制影响[29]。
在开花植物与昆虫的传粉互作系统中,花部结构性状会通过与传粉者的机械互作(mechanical interaction)影响植物的传粉效率和繁殖成功,因而会受到传粉者介导的自然选择[33-34]。有学者认为,鼠尾草属植物精巧的杠杆式背部传粉机制,其花部结构(或昆虫体型)的任何微小改变会对其传粉效率产生显著影响,因此花性状会受到强烈的传粉者选择[35]。本研究结果显示,在自然授粉条件下,毛地黄鼠尾草不同个体的繁殖效率与其花柱头高度显著负相关,即柱头位置较低的个体具有相对的适合度(或选择)优势;同时,该性状受到了显著的负定向选择作用。然而,在人工补充授粉条件下,该植物的繁殖效率与花柱头高度之间不存在显著的相关关系,同时也未检测到显著的选择梯度。这一结果表明,在自然授粉条件下,该植物的花柱头高度会显著影响个体的传粉效率和繁殖成功,从而受到传粉者介导的选择。Zhang 等[17]通过对该物种另一种群的花表型选择研究发现,该植物的花柱头高度未受到显著的选择作用,这可能是由传粉者类群及其与花结构互作在不同种群的变异性所引起[11]。
在鼠尾草属植物的花部结构性状中,花柱探出花冠长度决定着柱头的相对位置,是易受传粉者选择的另一关键性状[17]。本研究中,该植物的柱头探出花冠长度在自然授粉(花粉限制)条件下未受到显著的选择作用;相反,在补充授粉条件下,该性状受到了显著的负定向选择作用。换言之,在传粉不足的条件下,该植物柱头探出花冠长度的变异性并未导致不同个体间显著的雌性适合度差异;但是,在花粉供给充足时,该性状变异会显著影响其雌性适合度,柱头探出花冠外越长越不利于个体的繁殖成功。这一结果说明,该性状在当前种群并未通过与传粉者互作而受到显著选择。考虑到高海拔地区植物繁殖环境(如气候条件)的多变性,该植物在不存在花粉限制时,短花柱个体较长花柱个体具有适合度优势,可能是因为长花柱延长了花粉管生长发育时间,提高了植株个体授粉到受精的繁殖风险,因而会影响其繁殖成功;相反,较短的花柱能大大缩短授粉至受精的时间,有利于提高个体的繁殖成功率。因此,在自然授粉(花粉限制)条件下,即便长花柱个体具有相对的传粉优势,其传粉者对该性状的选择作用也会因为长花柱带来的负效应而抵消。因此,在该植物种群,花柱探出花冠长度可能同时受传粉者和气候因子选择的影响[2,36]。
当植物的两个性状通过互作影响个体的繁殖适合度时,该性状组合(trait combination)将受到相关选择作用(correlational selection)[26]。开花植物花性状的相关选择可导致花表型及其功能的整合[37]。本研究中,尽管毛地黄鼠尾草的花冠长与柱头高度的组合性状检测到了显著的相关选择梯度,但是其变量添加结果显示,该性状组合未表现出明显的相关选择趋势。另外,花冠口大小与柱头探出花冠长度受到了显著的负相关选择作用,表明花冠口较小、柱头探出花冠外较长的个体以及花冠口较大、柱头探出花冠外较短的个体均具有适合度优势。这种相关选择作用,很可能反映了该植物的花对不同大小传粉者类群的分化适应。因为小体型传粉者容易为花冠口较小、柱头探出花冠较长的花进行传粉;然而,大体型传粉者适宜为花冠口较大、柱头探出花冠较短的花进行传粉。同样,Zhang 等[17]在该物种的另一种群发现,该植物的花冠口高度与柱头探出花冠长度受到了显著的相关选择,而且该性状组合为了适应不同大小传粉者类群的年度变异表现出不同的选择模式。类似地,Zhang 等[25]研究发现,同属的雌-全花异株(gynodioecious)植物草地鼠尾草(Salvia pratensis)的花性状对种群内不同大小的传粉者类群表现出性别特异的适应性。
4 结论
毛地黄鼠尾草的繁殖成功在该自然种群存在明显的花粉限制,其花产量性状既可通过传粉者介导受到定向选择,也可直接影响个体的繁殖交配机会而受到选择。花部结构性状中,花柱头高度对个体的繁殖效率及适合度有显著影响,其选择作用主要通过传粉者介导而发生;花柱头探出花冠长度会影响个体的受精成功率,其选择作用主要受花粉萌发和受精环境(如气候因子)的影响;花冠大小与柱头位置相关性状的组合受到显著的相关选择作用。总体表明,毛地黄鼠尾草的花性状受到显著的传粉者介导选择;其中,花部结构性状为提高传粉效率表现出明显的功能整合趋向。
