新疆天山北坡荒漠草原碳通量特征及其对环境因子的响应
2022-07-08郭文章井长青邓小进陈宸赵苇康侯志雄王公鑫
郭文章,井长青,邓小进,陈宸,赵苇康,侯志雄,王公鑫
(新疆农业大学草业与环境科学学院,新疆草地资源与生态重点实验室,新疆 乌鲁木齐 830052)
陆地生态系统碳循环在全球碳循环研究中有重要意义[1],草地生态系统是陆地生态系统中最重要、分布最广的生态系统类型之一[2]。草地碳储量在全球陆地生态系统中仅次于森林[3],其碳储量高达308 Pg C,在陆地生态系统碳循环中承担着重要角色,在维持生态系统结构稳定方面具有关键作用[4]。目前,草地生态系统碳通量各组分季节性变化特征及其与环境因子间关系的研究已受到广泛关注[5]。涡度相关法可以长期测算生态系统碳通量[6],是目前最常用的碳通量观测方法,但是与样地实测数据相比较,使用涡度相关法观测的夜间碳通量总体是偏低的[7],由此导致对生态系统呼吸速率(ecosystem respiration,ER)的估算可能偏低[8],而箱式法可以直接测量生态系统呼吸速率,很好的避免这一问题。目前,我国碳通量研究主要集中于农田和森林生态系统[9],对草地生态系统碳通量的研究大多集中在典型草原、高寒草原等[10]。然而,由于天山北坡缺乏长期的草地碳通量观测站,目前为止在该区域荒漠草原碳通量的准确估算还存在较大的不确定性。
中国草地资源丰富,拥有各类天然草地394.93×104km2,约占陆地国土面积的41%,是全球草地生态系统的重要组成部分[11]。新疆天然草地资源丰富,面积辽阔,天山山区草地植被具有明显的垂直和水平地带性分布规律[12],天山北坡分布有山地荒漠、山地荒漠草原、山地草原、山地草甸草原、山地草甸及高寒草甸草地等多种草地类型,各具代表性的草地承担着维护生态和保护生物多样性的功能,是荒漠绿洲持续利用的天然绿色屏障[13]。随着全球气候变化和区域人类活动、土地利用等因素的干扰,对天山北坡草地生态系统碳循环过程产生了深远的影响。荒漠草原占新疆可利用草地面积的33.5%[14],是新疆重要的春秋牧场,但由于近年来超载放牧、持续放牧,导致荒漠草原退化严重,生态系统稳定性和恢复能力降低[15],生态环境十分脆弱[16],对人类活动和气候变化反应敏感[17]。目前,对于天山北坡荒漠草原生态系统碳循环的研究主要集中于对草地土壤碳储量的研究[18-19],而对天山北坡荒漠草原生态系统碳通量的变化特征及环境因素控制机制尚缺乏研究。选取新疆天山北坡山地荒漠草原为研究对象,采用静态箱式法对该地区的碳通量进行观测,旨在探明该地区荒漠草原碳通量的日变化和季节变化规律,分析荒漠草原碳吸收/排放变化特征及其环境驱动机制,为进一步准确评估新疆荒漠草原的碳汇功能区分布提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于天山北坡山地荒漠草原,隶属于新疆乌鲁木齐县南山谢家沟(87°17′44″ E,43°31′11″ N),海拔1742 m。属典型中温带大陆性气候,年平均气温2~5 ℃,年平均降水332 mm,降水主要集中在5-10月,年均蒸发量1008 mm。该研究区植被主要以镰芒针茅(Stipa caucasica)、羊茅(Festuca ovina)为优势种,以博洛塔绢蒿(Seriphidium borotalense)、针茅(Stipa capillata)、草原苔草(Carex liparocarpos)等为伴生种,灌木以刺叶锦鸡儿(Caragana acanthophylla)为主,植被覆盖度30%,6-8月平均地上生物量73.6 g·m-2、地下生物量1332.3 g·m-2。
1.2 研究方法
1.2.1 微气象因子观测 研究区架设有HOBO U30 微气象观测系统,主要监测指标包括风向、风速、光合有效辐射(photosynthetic active radiation,PAR)、太阳辐射、气压、降水量(precipitation,P)、土壤温度(soil temperature,Ts)、土壤含水量(soil water content,SWC)、空气温度(air temperature,Ta)、空气湿度(air humidity,RH)。通过数据采集器,每30 min 自动记录数据平均值。测量气象因子的传感器设备信息见表1。

表1 观测站点仪器装置信息(微气象观测系统)Table 1 Instrument and device information of observation station(Micro meteorological observation system)
1.2.2 试验设计 选取天山北坡山地荒漠草原为研究对象,在研究区选择微生境差异较小、有代表性的区域,设置3个10 m×10 m 的样地,3个样地间距5 m 以上。在每个10 m×10 m 的样地内,选取3个1 m×1 m 的小样方,并在每个样方内安装同化箱基座(内径50 cm×50 cm)1个,共9个同化箱基座,同化箱底座高8 cm,入地下5 cm,地上部3 cm,整个观测周期同化箱基座位置保持不变。采用CO2/H2O 非色散红外气体分析仪(LI-840A,LICOR,USA)测定净生态系统碳交换速率(net ecosystem carbon exchange,NEE)、生态系统呼吸速率。静态箱(0.5 m×0.5 m× 0.5 m)由透明有机玻璃构成,箱体内配有2个风扇,可使箱内气体充分混匀。测定前,先抬起静态箱,使箱内空气与周围气体混匀,测量时,将静态箱放置在同化箱基座上,保持整个箱子密闭状态,开始测定箱内气体浓度变化,设置每秒自动记录一次CO2浓度值,记录120 s 左右。净生态系统碳交换速率测量结束后,抬起箱子使箱内气体与外界气体充分混匀,将静态箱放置在同化箱基座上,在箱子外盖上黑色不透光的布罩,开始测定箱内气体浓度变化,记录120 s,用于测定生态系统呼吸速率。每个样方重复测定3 次,在测定净生态系统碳交换速率和生态系统呼吸速率的同时,用手持气象站(Kestrel)记录对应时段内风速、气压、空气温度、空气湿度。
2020年生长季 6-10月每月中旬(6月 16 日、7月 14 日、8月 15 日、9月 16 日、10月 15 日)测定 1 次,每次监测时间为1个完整昼夜24 h,于当日10:00 开始,至次日10:00 结束,日间测定间隔为2 h,夜间测定间隔为3 h。
1.2.3 数据处理 采用公式(1)计算生态系统CO2交换量:

式中:FC是 CO2通量(μmol·m-2·s-1);V是箱体的体积(m3);Pav是测量期间箱体内的平均大气压强(kPa);Wav是测量期间箱体内水汽分压(mmol·mol-1);R是大气常数(8.314 J·mol-1·K-1);S是同化箱的面积(m2);Tav是测量期间箱体内平均温度(℃);dc/dt是箱体内气体浓度变化率。
采用公式(2)直角双曲线模型[20]模拟白天净生态系统碳交换速率与光合有效辐射的关系:

式中:NEEd为白天净生态系统碳交换速率(μmol·m-2·s-1);α为表观量子效率(μmol·mol-1);Amax为最大光合速率(μmol·m-2·s-1);Rd为日间生态系统呼吸速率(μmol·m-2·s-1);PAR为日间光合有效辐射(μmol·m-2·s-1)。
采用公式(3)Van’t Hoff 模型[21]模拟温度与生态系统呼吸速率的关系:

式中:ER为生态系统呼吸速率(μmol·m-2·s-1);T为温度(℃);α为反映生态系统基础呼吸速率的参数;β为生态系统呼吸速率温度敏感性的参数。
采用公式(4)计算生态系统呼吸速率对温度变化敏感程度Q10[22]:

式中:Q10为生态系统呼吸速率温度敏感系数;β为生态系统呼吸温度敏感性参数。
采用公式(5)计算饱和水汽压差(vapor pressure deficit,VPD)[23]:

式中:VPD为饱和水汽压差(kPa),Ta为空气温度(℃),RH为空气湿度(%)。
采用SPSS 22.0 对试验数据进行统计分析,用Canoco 5.0 进行冗余分析,Origin 2018 作图。
2 结果与分析
2.1 荒漠草原环境因子季节变化特征
由图1 可以看出,2020年生长季6-10月荒漠草原环境因子具有明显的季节变化特征,观测期内日均光合有效辐射在 62.35~663.59 μmol·m-2·s-1,月均光合有效辐射在 6月最高,达到 466.86 μmol·m-2·s-1,10月最低为278.22 μmol·m-2·s-1,受天气变化影响,光合有效辐射波动剧烈。日均饱和水汽压差在0.36~1.52 kPa 之间波动,呈先升高再降低趋势,8月份饱和水汽压差高于其余月份。生长季总降水量为192 mm,主要集中在7月(70 mm),降水频率(58 次)。土壤含水量在0.04~0.17 m3·m-3之间波动,降水与5 cm 土壤含水量有很强的响应关系。空气温度与5 cm 土壤温度季节变化趋势相似,二者日均值变化范围分别为-1.40~23.34 ℃和0.91~25.53 ℃,土壤温度明显高于空气温度,但升温时间滞后于空气温度。

图1 荒漠草原环境因子季节变化Fig.1 Season variations of environmental factors in desert steppe
2.2 荒漠草原净生态系统碳交换速率日变化特征
荒漠草原6-9月净生态系统碳交换速率日变化均呈倒“U”型曲线(图2),且有明显的日变化规律。净生态系统碳交换速率在白天8:00 左右开始变为负值,此时生态系统由夜间的碳源转变为白天的碳汇,随着温度升高,光合有效辐射增强,10:00-12:00 碳吸收达到峰值,直至19:00 左右净生态系统碳交换速率转变为正值,生态系统开始向大气中释放CO2,荒漠草原从碳汇转变为碳源。荒漠草原10月进入生长季末期,随着气温下降、光合有效辐射降低,植物的光合作用和呼吸作用都很微弱,净生态系统碳交换速率日变化趋势变得十分平缓。

图2 荒漠草原净生态系统碳交换速率日变化Fig. 2 Diurnal variation of net ecosystem carbon exchange in desert steppe
荒漠草原生态系统每日吸收CO2开始时间以8月(8:00)最早,以 9月(9:30)最晚,每日吸收CO2结束时间以 9月(14:00)最早,以 8月(20:00)最晚。在整个生长季日变化过程中,净生态系统碳交换速率最低为-2.16 μmol·m-2·s-1,出现在 8月(10:00);净生态系统碳交换速率的最高为 3.01 μmol·m-2·s-1,出现在7月(22:00)。
2.3 荒漠草原生态系统呼吸速率日变化特征
荒漠草原生长季生态系统呼吸速率日变化均呈“单峰”曲线,具有明显的季节变化规律。由图3 可以看出,7月生态系统呼吸速率日变化波动最大,最大值为 6.92 μmol·m-2·s-1,最 小 值 为 1.71 μmol·m-2·s-1,日均值为 4.12 μmol·m-2·s-1;10月生态系统呼吸速率日变化波动最小,最大值为 0.64 μmol·m-2·s-1,最小值 为 0.02 μmol·m-2·s-1,日 均 值 为 0.25 μmol·m-2·s-1。不同月份生态系统呼吸速率峰值出现时间不同:7月在16:00 到达峰值,其他月份出现时间主要集中在14:00 左右。生态系统呼吸速率日变化随着时间推移,在 12:00-16:00 之间到达峰值之后开始下降,在6:00-8:00 之间生态系统呼吸速率最小。荒漠草原生态系统呼吸速率排序大小为7>9>8>6>10月。

图3 荒漠草原生态系统呼吸速率日变化Fig. 3 Diurnal variation of ecosystem respiration in desert steppe
2.4 荒漠草原生态系统碳通量变化特征
如图4 所示,6-8月荒漠草原生态系统均表现为碳吸收,月累积净固碳量分别为4.15、4.75、23.03 g C·m-2,而 9、10月表现为碳排放,累积碳释放量为13.46、2.97 g C·m-2;荒漠草原生态系统 6-10月表现为微弱的碳汇,总固碳量为15.50 g C·m-2。生态系统呼吸碳排放量在 7.92~128.42 g C·m-2之间,7月生态系统碳排放量最大,为 128.42 g C·m-2;10月碳排放量最少,为7.92 g C·m-2。生长季生态系统呼吸碳排放总体呈先增加后减小的趋势;总初级生产力(gross primary productivity,GPP)的 变 化 范 围 在4.95~133.17 g C·m-2之间,均值为 63.50 g C·m-2。

图4 荒漠草原碳通量的季节变化Fig.4 Seasonal changes of carbon fluxes in desert steppe
2.5 净生态系统碳交换速率与光合有效辐射的关系
由图5 可以看出,荒漠草原生长季NEE 与PAR关系符合直角双曲线模型[14]。拟合方程为NEE=R2=0.72(P<0.01)。白天NEE 随着PAR 的增大而逐渐减小,生态系统对CO2的吸收量逐渐增大,碳汇能力也逐渐增强,但当PAR 增加到光饱和点(PAR=1200 μmol·m-2·s-1),光合速率达到最大。生长季荒漠草原生态系统白天最大光合速率为 3.63 μmol·m-2·s-1,白天平均生态系统呼吸速率为 2.26 μmol·m-2·s-1。

图5 荒漠草原净生态系统碳交换与光合有效辐射的关系Fig. 5 Relationship between net ecosystem carbon exchange and photosynthetic active radiation in desert steppe
2.6 生态系统呼吸速率与土壤温度、空气温度的关系
通过Van’t Hoff 指数方程分别拟合生长季各月生态系统呼吸速率与土壤温度、空气温度的回归关系。由表2 可得,Van’t Hoff 指数方程能很好地描述生态系统6-9月的呼吸速率对土壤温度和空气温度的响应,土壤温度可以解释生态系统呼吸速率日变化的55.2%~86.3%,土壤温度敏感性Q10范围为 1.85~2.71,对不同月份Q10进行比较,8月(1.85)<9月(1.94)<7月(2.09)<6月(2.71)。空气温度可以解释生态系统呼吸速率日变化的70.1%~97.1%,空气温度敏感性Q10范围为2.31~2.80;不同月份生态系统呼吸速率日变化温度敏感性有差异,对不同月份Q10进行比较,8月(2.31)<9月(2.33)<6月(2.53)<7月(2.80);10月生态系统呼吸速率与土壤温度、空气温度无明显相关性。

表2 生态系统呼吸速率与温度指数回归关系Table 2 Exponential regression between ecosystem respiration and temperature
2.7 荒漠草原生态系统呼吸速率与环境因子之间的关系
如图6 所示,生长季荒漠草原生态系统呼吸速率与空气温度、土壤温度呈显著正相关关系(P<0.01),随着空气温度和土壤温度的增加,生态系统呼吸速率逐渐增加。生态系统呼吸速率与土壤含水量符合二次函数关系,相关性显著(P<0.01),土壤含水量在0.162 m3·m-3时,生态系统呼吸速率最高,土壤含水量过低或过高都会抑制生态系统呼吸速率。生态系统呼吸速率与空气湿度呈线性负相关关系(P<0.01),说明随着空气湿度的增加,生态系统呼吸速率逐渐减小。

图6 荒漠草原生态系统呼吸速率与环境因子之间的关系Fig.6 Relationships between respiration and environmental factors in desert steppe ecosystem
2.8 不同月份NEE 和ER 与环境因子的相关关系
NEE、ER 与环境因子的相关性分析结果见(表 3)。结果表明:生长季 6-8月,NEE 与 PAR、VPD、Ta、Ts、SWC 均呈负相关关系(P<0.05);生长季 6-8月,NEE 与 RH 呈正相关关系;10月份 NEE 与 PAR、VPD、Ta、RH、Ts、SWC 无明显相关性。生长季 7-9月,ER 与 VPD、Ta、Ts、SWC 存在显著正相关关系(P<0.01),ER 与RH 呈显著负相关关系(P<0.01);10月 ER 与 PAR、VPD、Ta、RH、Ts、SWC 无明显相关性。
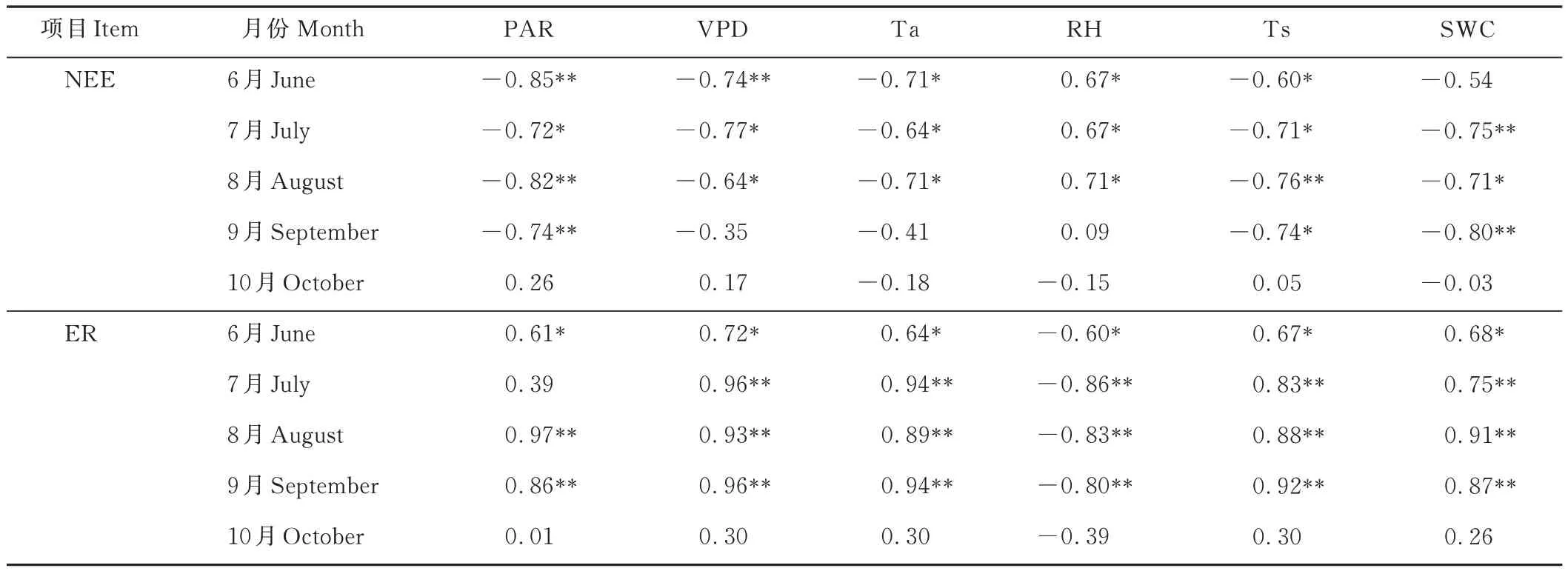
表3 不同月份NEE、ER 与环境因子的相关系数Table 3 Correlation coefficients between NEE,ER and environmental factors
2.9 生长季荒漠草原NEE 和ER 与环境因子的相关性分析
分析不同环境因子[PAR、VPD、Ta、RH、Ts(5、20、40 cm)、SWC(5、20、40 cm)]对荒漠草原NEE、ER 日变化的影响程度。图7 为冗余分析结果,NEE、ER 在第1轴和第2轴的解释量分别为58.60%和25.17%,即第一排序轴和第二排序轴10 种环境因子累计解释NEE、ER 的83.77%,表明第一排序轴和第二排序轴可以较好地反映NEE、ER 与环境因子的关系,且主要由第1轴决定。SWC(5 cm)、Ta 在ER 箭头连线上的投影长度较长,表明ER 日变化的主导因子主要是 SWC(5 cm)、Ta;PAR在NEE 箭头反向延长线上的投影较长,表明NEE 日变化由PAR 主导,并呈较强的负相关关系;Ts(20、40 cm)、SWC(20、40 cm)均与 NEE、ER 无显著相关关系。

图7 NEE 及ER 与环境因子相关性的冗余分析排序Fig. 7 Redundancy analysis on the correlation between NEE and ER with environment properties
冗余分析排序图仅反映出NEE、ER 与环境因子间存在相关性,难以量化环境因子对NEE、ER 的贡献程度。为此,本研究采用蒙特卡洛检验和前向选择排序法对10 种环境因子进行排序。由表4 可知,各环境因子对NEE、ER 的影响存在一定差异;各环境因子对NEE、ER 影响的重要性排序从大到小依次为SWC(5 cm)>Ta>PAR>VPD>Ts(5 cm)>RH>Ts(20 cm)>Ts(40 cm)>SWC(40 cm)>SWC(20 cm),其中 SWC(5 cm)、Ta、PAR、VPD、Ts(5 cm)、RH 对NEE、ER 有显著影响(P<0.01),其他环境因子对NEE、ER 的影响均未达显著水平(P>0.05),其中SWC(5 cm)、Ta、PAR 对 NEE、ER 的贡献率较高,分别为 41.7%、34.0%、32.1%,表明 SWC(5 cm)、Ta、PAR 是影响研究区NEE、ER 的关键因素。

表4 环境因子解释量及显著性检验Table 4 Explanation rate and significance test of environment factors
3 讨论
3.1 荒漠草原碳通量日变化特征
天山北坡荒漠草原生态系统6-9月净生态系统碳交换速率日变化呈倒“U”型曲线变化,日间表现为碳汇,夜间表现为碳源。10月荒漠草原处于生长季末期,温度和光合有效辐射较低,导致净生态系统碳交换速率日变化趋于平缓,这与前人的大多数研究结果一致[24-26]。6、7、8月荒漠草原植被处于生长旺盛期,光合作用较强,表现为碳吸收,9、10月,荒漠草原植被覆盖度低,光合作用较弱,表现为碳排放。荒漠草原净生态系统碳交换速率最大为 2.16 μmol·m-2·s-1,低于云贵高原草地(3.82 μmol·m-2·s-1)[24]、天山北坡草甸草原(7.62 μmol·m-2·s-1)[27]、黄土高原半干旱区草地生态系统(7.90 μmol·m-2·s-1)[28],这是由于荒漠草原植被稀疏,覆盖度较低,并且长期过度放牧导致草地退化,使得该地区草地固碳能力较弱。
荒漠草原生态系统呼吸速率日变化呈“单峰型”变化趋势,峰值出现在14:00-16:00,且日间高于夜间,夜间生态系统呼吸速率维持在较低水平,这是由荒漠草原土壤温度日变化引起的。孙学凯等[29]研究表明科尔沁沙质草地生态系统呼吸速率日变化为单峰曲线,生态系统呼吸速率峰值出现在15:00;邢鹏飞等[30]研究表明晋北农牧交错带草地生态系统速率的日变化呈单峰型变化趋势,峰值出现在13:00-16:00;李愈哲等[31]研究得出不同利用方式下锡林郭勒温性草原生态系统呼吸速率日变化表现出单峰变化趋势,本研究结果与之相似。生长季荒漠草原生态系统呼吸速率为7>9>8>6>10月,主要原因是5月荒漠草原开始进入返青期,生态系统呼吸速率较弱,7、8、9月植被进入生长高峰期,适宜的水热条件促进了植物根系活动,使生态系统呼吸速率增大,10月以后,植被进入生长季末期,生态系统呼吸速率逐渐减弱。
3.2 荒漠草原碳通量的影响因子
通过冗余分析得出日间净生态系统碳交换速率主要影响因子是光合有效辐射,净生态系统碳交换速率与光合有效辐射符合直角双曲面模型,与前人相关研究结果一致[32-34],光合有效辐射是影响荒漠草原日间净生态系统碳交换速率的主要环境因子。光合有效辐射在500 μmol·m-2·s-1以下时,净生态系统碳交换速率随光合有效辐射升高而迅速降低,光合有效辐射在1200 μmol·m-2·s-1以上时,净生态系统碳交换速率随光合有效辐射变化程度逐渐减小,并接近饱和,这与大多数文献的研究结果一致[25,35]。
本研究中,荒漠草原生态系统呼吸速率受温度和水分共同影响,生态系统呼吸速率与空气温度和5 cm 土壤温度呈指数相关。研究表明,温度和水分都显著影响生态系统呼吸速率,但因水热条件的区域差异导致不同生态系统间呼吸速率存在较大的异质性[36-37]。例如,在比较干旱的生态系统中,生态系统呼吸速率与温度呈指数关系,同时受土壤含水量的影响[38];也有研究表明土壤温度越低,生态系统呼吸速率越大[39];而在降水充足的草地生态系统中,增温可以促进生态系统呼吸速率[40]。Q10值是评价生态系统呼吸速率对温度变化敏感程度的指标[41-42],全球草地生态系统Q10介于 1.35~3.48 之间[43],通过分析得出,6-10月荒漠草原土壤温度与生态系统呼吸速率Q10值介于1.85~2.71 之间,空气温度与生态系统呼吸速率Q10值介于2.31~2.82 之间,与全球草地生态系统呼吸Q10相一致,Q10随着温度的升高而下降,这与许多试验结果相一致[44]。通过冗余分析得出,土壤含水量是影响天山北坡荒漠草原生态系统呼吸速率的主要环境因子,6-9月生态系统呼吸速率与土壤含水量呈显著正相关。胡毅等[27]研究表明,天山北坡草甸草原生态系统呼吸速率与土壤含水量呈指数相关关系;李成一等[45]分析表明,青藏高原高寒湿地生态系统呼吸速率与土壤含水量无相关关系;本研究中荒漠草原生态系统呼吸速率与土壤含水量呈二次函数关系,这可能是荒漠草原土壤含水量较低,土壤有机质含量较低,生态系统呼吸速率对水分变化极为敏感造成的。
4 结论
本研究通过分析天山北坡荒漠草原碳通量的变化特征及其与环境因子的相关关系,研究结论如下:1)天山北坡荒漠草原净生态系统碳交换速率日变化呈倒“U”型曲线,净生态系统碳吸收速率大小依次为8>7>6>9>10月;生长季各月生态系统呼吸速率均呈单峰型曲线,生态系统碳排放量白天大于夜间;2)荒漠草原生态系统生长季总固碳量为15.50 g C·m-2,表现为微弱的碳汇;6-8月荒漠草原生态系统表现为碳汇,9-10月表现为碳源;3)日间净生态系统碳交换速率与光合有效辐射之间符合直角双曲线模型;影响净生态系统碳交换速率日变化的主导因子是光合有效辐射,影响生态系统呼吸速率的主要因子是5 cm 土壤含水量和空气温度。
