基于耳石微化学的东海和黄海南部近海小黄鱼幼鱼的空间连通性研究
2022-07-04王鹏凯李圣法刘尊雷姜亚洲
王鹏凯,李圣法,刘尊雷,金 艳,姜亚洲,张 辉,张 翼
(1.农业农村部东海渔业资源开发利用重点实验室,中国水产科学研究院东海水产研究所,上海 200090;2.上海海洋大学海洋科学学院,上海 201306)
鱼类在仔稚鱼阶段游泳能力较弱,海流等物理海洋过程的作用驱动了仔稚幼鱼的输运和扩散,为不同产卵场仔稚鱼的混合和交换提供了机会[1],从而影响了不同产卵场的仔稚鱼群体之间的种群连通性,扩散促进了这些复合种群的存续[2]。然而,受限于仔稚鱼阶段的个体大小和高死亡率,要追踪鱼类从孵化开始的生活史是有难度的。研究发现耳石微化学是研究鱼类生态学的有效工具[2]。耳石在鱼类的整个生命周期中不断生长,其化学成分保持惰性,栖息环境的化学元素经过一系列的生理过程被聚合到耳石的碳酸钙结构中[3],按时间顺序保存了不同生活史阶段经历的环境信息轨迹[4-5],随着耳石化学组成与水化学之间关系研究的不断深入,以及耳石元素分析准确性的不断提高,耳石微化学正逐步用于研究鱼类群体的起源、种群动态、洄游分布以及生境连通性[6-11]。耳石核区的微化学特征常用于追溯个体的出生来源[2,12-13],耳石边缘则可用于反映鱼类栖息的生境特征[2,14]。
小黄鱼(Larimichthys polyactis)是我国重要的经济鱼类,为近底层洄游性鱼类,广泛分布于我国沿海。目前,对黄海南部和东海的小黄鱼种群连通性的理解还存在着不少的分歧[15-16],黄海南部和东海小黄鱼群体是否为同一种群、不同群体的混栖、交换等种群连通性问题尚需通过新的技术手段和方法加以深入研究。近年来,在小黄鱼种群结构划分的研究中已开始探索应用耳石微化学方法[17-19]。
本文利用小黄鱼幼鱼耳石边缘的耳石元素分析东海至南黄海近海幼鱼栖息海域之间的连通性,并利用小黄鱼幼鱼耳石近核区耳石元素评估这些幼鱼的出生地来源。以期为小黄鱼不同群体的起源、洄游分布和生境连通性研究提供参考。
1 材料方法
1.1 样本采集与处理
2019年5—6月在黄海南部近岸(射阳、吕四)和东海近岸(舟山、大陈)4个采样点用仔稚鱼网(口径1.3 m,网目0.5 mm)采集小黄鱼幼鱼(表1)。小黄鱼样品于-20℃冷冻保存备用。鱼体解冻后测量体长,解剖取出矢耳石,清洗表面的有机质后,室温下干燥保存待用。耳石切片制作过程参照徐浩等[19]的方法。环氧树脂固定包埋耳石后,经StruersDiscoplan-TS上切割,用1 200目砂纸精磨至核心暴露,制成厚度约为0.5 mm的耳石薄片。合格的耳石薄片放入超纯水中清洗5 min,自然晾干后即可用于耳石微化学分析。

表1 小黄鱼幼鱼采集样品信息Tab.1 Sampling details of Larimichthys polyactis juveniles
1.2 耳石元素指纹分析
采用激光剥蚀电感耦合等离子体质谱仪(LA-ICP-MS)对耳石元素进行分析。激光剥蚀进样系统采用NWR-213仪器(New Wave Research,Fremont,美国),电感耦合等离子体质谱系统采用Thermo X SeriesⅡICPMS仪器(Thermo Fisher Scientific)。每批样品包括4个耳石切片和1个NIST612标样(National Institute of Standards and Technology,美国)、1个碳酸盐标样MACS-3(United States Geological Survey,美国)。元素分析参照徐浩等[19]的研究方法,详细步骤为:测试过程按NIST612—MACS-3—耳 石 样 品—NIST612—MACS-3的顺序循环进行,其中NIST612用于校正ICP-MS元素分析过程中仪器的漂移。激光剥蚀出的样品通过Ar和He混合气体送入ICP-MS进行元素分析,He气流量控制在0.8 L·min-1。在无任何样品条件下对Ar、He混合气体进行LAICP-MS测试,重复10次,3倍空白标准偏差所对应的浓度即为各元素检测限水平。LA-ICP-MS对小黄鱼耳石核区和边缘区域检测和识别出有效元素:Sr、Ba、Mn、Mg、Ca。测定的各元素值以其浓度与Ca2+浓度比值表示。样品分析采用多测点激光剥蚀技术:耳石核心区剥蚀点位于耳石原基附近,耳石边缘位置剥蚀点紧靠耳石边缘(图1)。每个激光剥蚀点直径40μm,采样深度15 μm,时间15 s。
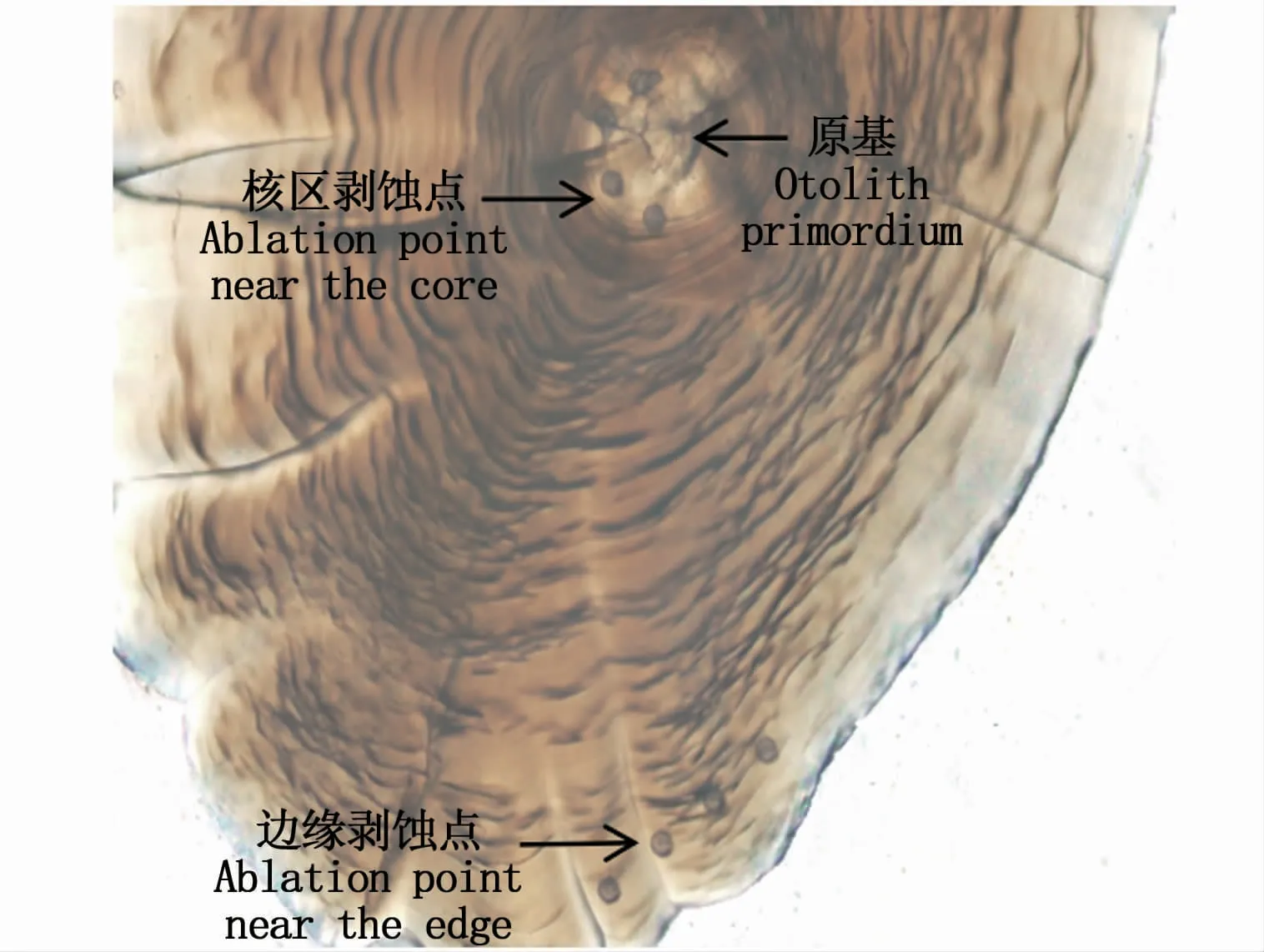
图1 小黄鱼耳石核区和边缘多点激光剥蚀定位Fig.1 Photographs showing the checking and edge of the multiple laser ablations on an otolith section of Larimichthys polyactis
1.3 元素指纹数据处理
小黄鱼耳石近核区和边缘元素比值数据的地理差异检验使用非参数检验方法,该方法无需考虑数据是否符合正态和方差齐性的假设条件。选用Kruskal-Wallis非参数检验方法分析单个元素比值地理上的总体差异显著性,对总体差异显著的元素比值再用Dunn多重比较检验分析两两采样点间的显著性差异[2]。Kruskal-Wallis检验使用R语言stat包中的kruskal.test()函数,Dunn多重比较检验使用pairwiseComparisons包中的parwise_comparisons()函数,并结合ggplot2包和ggsignif包绘制箱形图及显著性图示。
小黄鱼耳石近核区和边缘元素比值的采样点间差异的多元检验使用非参数置换方差分析(PERMANOVA)和主坐标分析(principal coordinate analysis,PCoA)方法评估和图示采样点间的空间关系[20]。元素比值进行开四次方转换,以平衡数据对相异性值的贡献,基于欧氏距离计算采样点间的相异性矩阵,PERMANOVA采用9 999次随机重复的无限制置换进行分析,当呈显著性差异时,再进行采样点间的两两多重比较以确认均值差异的来源。PERMANOVA分析使用R语言vegen包中的adonis2()函数,采样点间的两两多重比较使用pairwiseAdonis包中的pairwise.adonis2()函数,应用vegan包中的cmdscale()函数进行PCoA模型拟合,并用venfit()函数计算每个元素比值的方向和强度,使用ggplot2包绘制PCoA结果图和每个有样点的95%置信椭圆。
应用随机森林(RF)分类方法评估小黄鱼幼鱼耳石边缘元素比值的空间特征,以判断采样点间的空间判别效果[2]。RF方法已广泛应用于耳石元素比值的研究中,与其他分类方法相比,RF并不需要元素比值数据符合某种分布假设,且判别效果优于其他方法[21-22]。随机选择75%的小黄鱼幼鱼样本的耳石边缘元素比值构建和训练每个可能的RF分类器。剩余25%的样本用于检验预测未知耳石边缘元素比值来源的总体可靠性,用分类准确率和真实技巧统计量(TSS)评价模型的预测精度,准确率表示样本正确地重分类到其采样点的总数,而TSS也解释了真实的负面预测。TSS的范围为-1到+1,其中+1代表对给定栖息地存在或缺失的100%的正确预测,0表示随机预测,-1表示100%的错误预测。最后,通过计算将全局基尼指数(Gini index,GI)从最优分类器中去除后的平均下降量来评价各元素对空间分辨的贡献。GI的范围从0(所有元素的贡献率都均等)到100(当单个元素贡献率达到100%)。当一个元素从分类器中去除时,GI的降低越高,说明该元素对识别的重要性越大[2,23-24]。
应用无监督的随机森林(RF)聚类方法[25]分析小黄鱼耳石近核区元素比值以得到其可能的出生地来源,该方法分为两步[2,26]:第一步是忽略采样点信息,从不同元素的实际边际分布的乘积中随机取样,创建人造数据集。接着,RF用于区分观测的人造数据(用作分类因子)观测,生成相似性矩阵(similarity matrix),定义为两个个体最终在树的同一终端节点的频率。第二步,计算相异性矩阵(dissimilarity matrix),计算公式为:
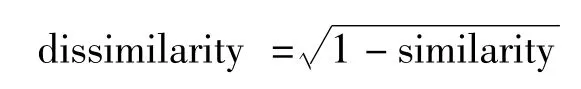
将相异性矩阵代入基于中心点(partitioning around medoid,PAM)聚类确定聚类数和分组,PAM聚类使用fpc包中的pamk()函数确定最佳的聚类数。
2 结果与分析
2.1 小黄鱼幼鱼耳石边缘元素比值的地理差异比较
小黄鱼幼鱼耳石边缘元素比值的Kruskal-Wallis检验结果显示,4种元素比值的地理差异都非常显著(表2)。两两采样点间的Dunn多重比较显示(表3),Sr∶Ca呈显著性差异的点对为射阳与吕四、吕四与舟山及吕四与大陈(图2-A);Ba∶Ca呈显著差异的点对只有射阳与吕四及射阳与舟山(图2-B);Mg∶Ca呈显著差异的点对为吕四与射阳、吕四与舟山及吕四与大陈(图2-C);Mn∶Ca呈显著差异的点对为射阳与吕四、射阳与舟山、吕四与大陈及舟山与大陈(图2-D)。

图2 小黄鱼幼鱼耳石边缘元素比值的地理差异比较Fig.2 Comparison about otolith edge microchemistry of juvenile Larimichthys polyactis among sampling locations
PERMANOVA检验显示,小黄鱼幼鱼耳石边缘元素比值在4个采样点间呈现极显著的地理差异(F3,80=13.37,P<0.001);两两采样点间的多重PERMANOVA检验显示,除了吕四与舟山采样点间不显著外,其他采样点两两间呈显著差异,射阳与吕四(F1,29=20.99,P<0.001),射阳与舟山(F1,47=35.46,P<0.001)、射阳与大陈(F1,32=6.77,P<0.01)、吕四与大陈(F1,33=7.68,P<0.01)和舟山与大陈(F1,51=10.38,P<0.001)。PCoA的结果显示(图3),第1主轴和第2主轴的累积贡献率达95.43%,小黄鱼幼鱼耳石边缘元素比值呈现较明显的空间差异,吕四和舟山小黄鱼样本位置相对较近,射阳小黄鱼样本的空间位置与其他取样点相对较远。
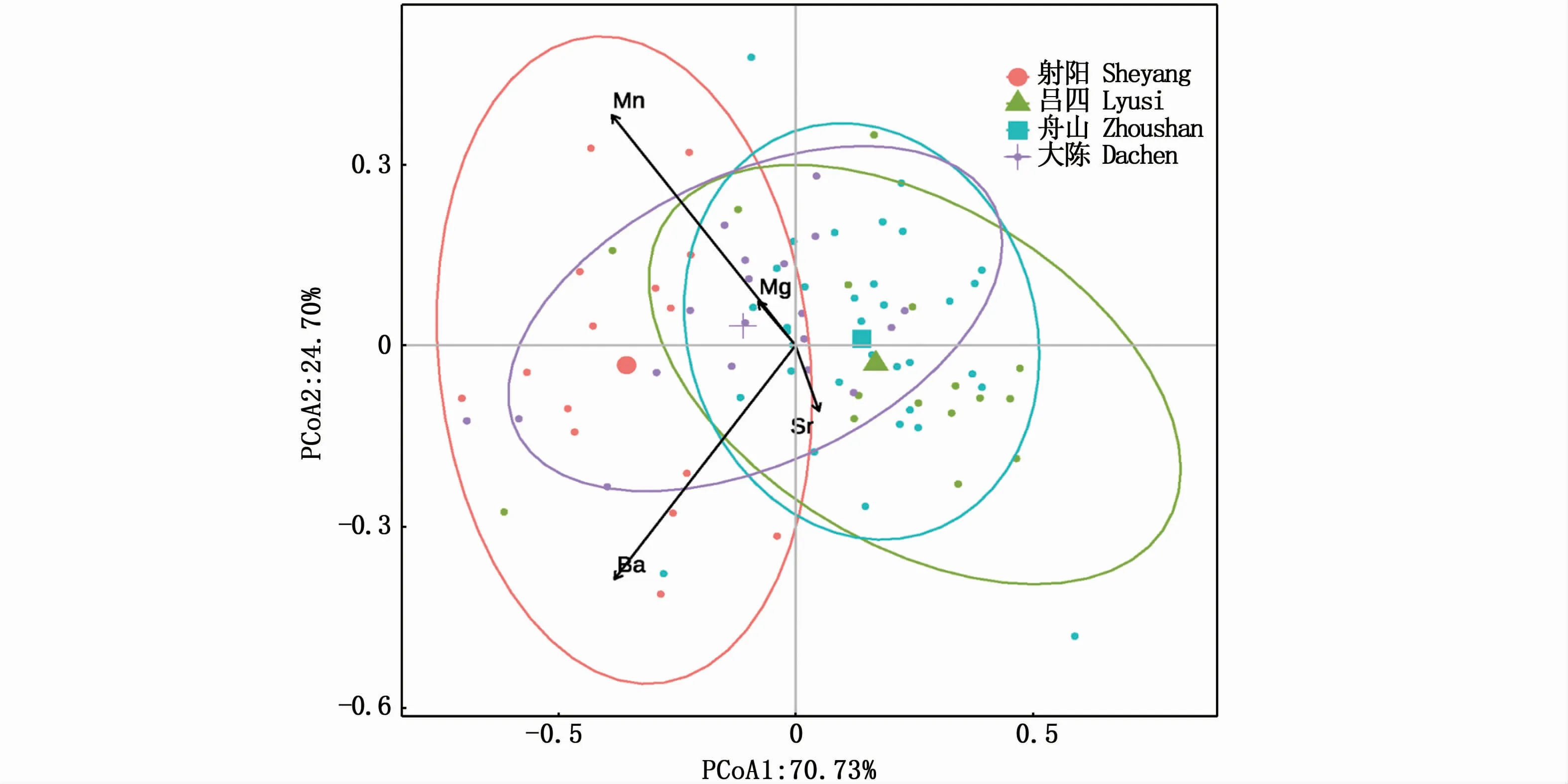
图3 小黄鱼幼鱼耳石边缘元素比值的空间PCoA排序图Fig.3 PCoA plot comparing otolith edge microchemistry of juvenile Larimichthys polyactis
小黄鱼幼鱼耳石边缘元素比值的RF分类的总分类准确度为63%,各采样点的分类准确度在50%~75%之间(表4),其中射阳和舟山小黄鱼幼鱼耳石边缘元素比值重新分配的准确度>60%,吕四和大陈为50%。TSS统计值均大于0,最高为射阳(0.67),最低为大陈(0.37)。各元素对采样点分类准确度的贡献率差异很大,4种元素比值的平均基尼系数下降(mean decrease of Gini index)变化不大(11.01~11.96),其中Mn比值的重要性最高(11.96),其次为Ba比值(11.61)和Mg比值(11.50),Sr比值的重要性最低(11.01)。
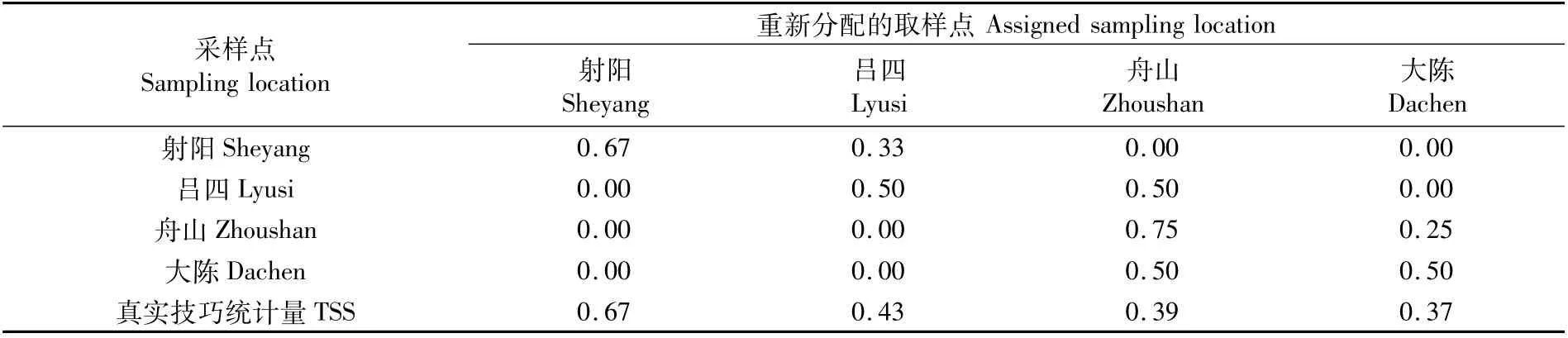
表4 小黄鱼幼鱼耳石边缘元素比值随机森林分类的准确度(%)和真实技巧统计量TSSTab.4 Classification accuracy(%)and true skill statistics(TSS)of random forest classification for the otolith edge of juvenile Larimichthys polyactis
2.2 小黄鱼幼鱼耳石近核区元素比值的地理差异比较
4个采样点小黄鱼幼鱼耳石近核区4种元素比值的Kruskal-Wallis检验结果显示,除了Mn∶Ca外(图4-D),其他元素比值呈现显著性的地理差异。对3种Kruskal-Wallis检验呈显著性的元素比值进行采样点两两间Dunn多重比较显示(表5),Sr∶Ca呈现显著性差异的采样点对为:射阳与吕四、吕四与大陈及吕四与舟山(图4-A);Ba∶Ca为射阳与舟山、射阳与大陈、吕四与舟山及吕四与大陈(图4-B);Mg∶Ca为吕四与舟山、吕四与大陈(图4-C)。
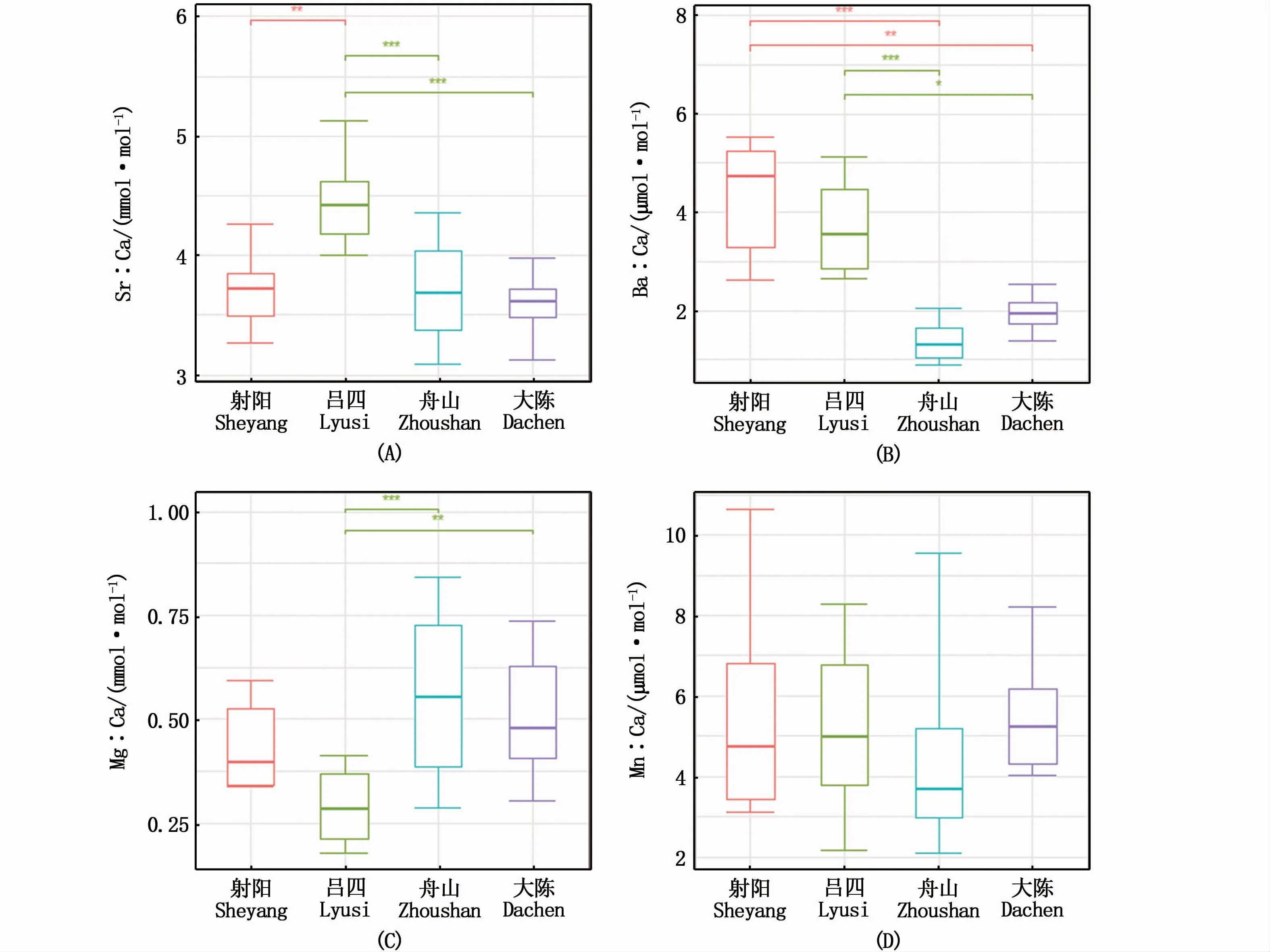
图4 采样点间小黄鱼幼鱼耳石近核区元素比值比较Fig.4 Comparison about otolith near-core microchemistry of juvenile Larimichthys polyactis among sampling locations

表5 小黄鱼幼鱼耳石近核区采样点间元素比值的Dunn多重比较Tab.5 Dunn’s multiple comparison of otolith near-core microchemistry of juvenile Larimichthys polyactis
PERMANOVA检验显示,小黄鱼幼鱼耳石近核区4种元素比值在4个采样点间呈现极显著的地理差异(F3,80=11.79,P<0.001);两两采样点间的多重PERMANOVA检验显示,射阳与吕四间差异不显著(F1,29=1.58,P=0.20>0.05),舟山与大陈间差异显著(F1,51=4.12,P=0.02<0.05),其他采样点间差异极显著(射阳与舟山:F1,47=18.98,P=1×10-4<0.001,射阳与大陈:F1,32=12.47,P=1×10-4<0.001,吕四与舟山:F1,45=17.28,P=1×10-4<0.001)。PCoA的结果显示(图5),第1主轴和第2主轴的累积贡献率达88.56%,小黄鱼幼鱼耳石元素比值呈现较明显的空间差异,射阳和吕四小黄鱼样本位置相对较近,舟山和东引岛小黄鱼样本的空间位置相对较近。
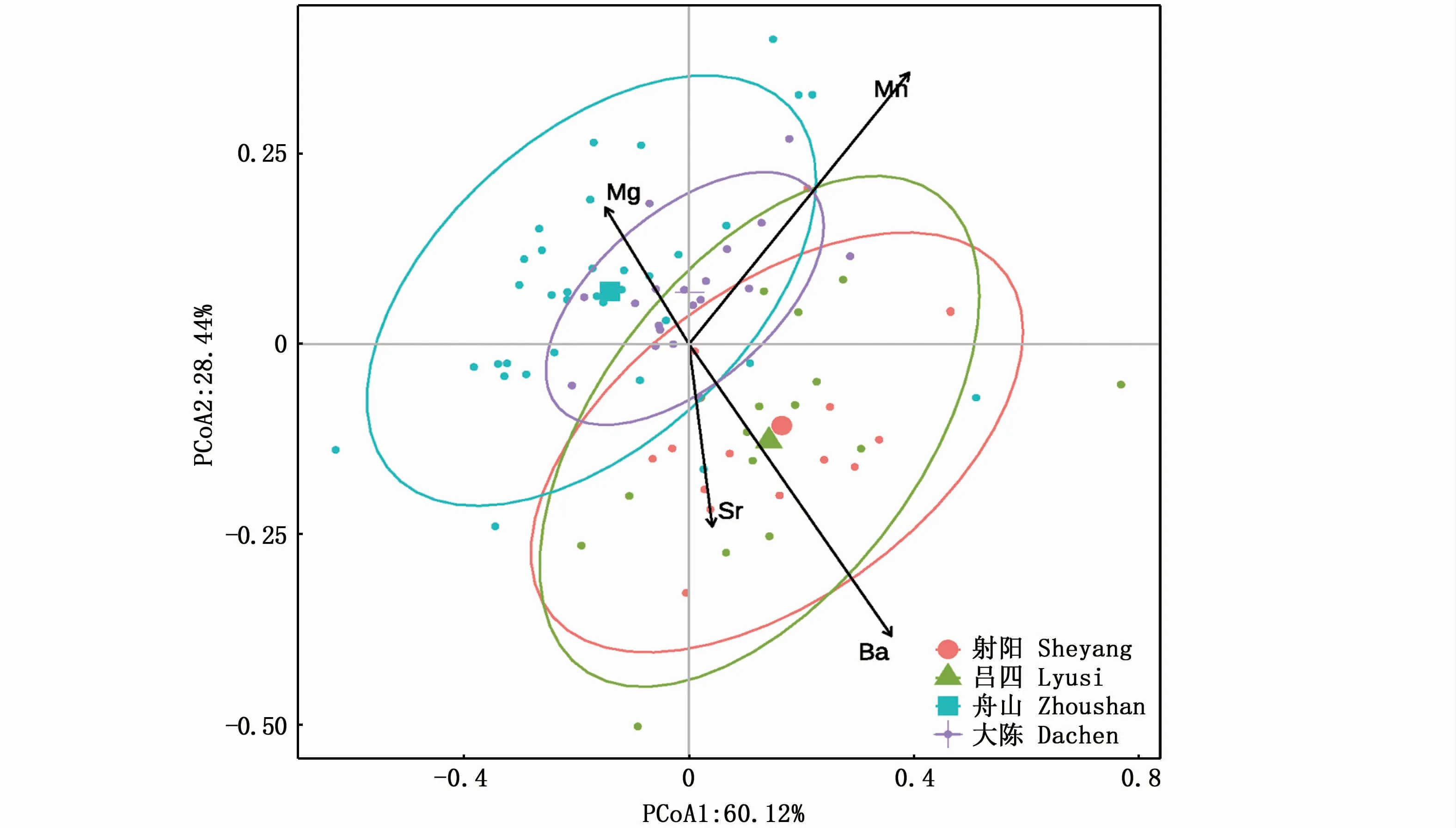
图5 小黄鱼幼鱼耳石近核区元素比值的PCoA排序图Fig.5 PCoA plot comparing otolith near-core microchemistry of juvenile Larimichthys polyactis
2.3 小黄鱼幼鱼出生地来源
根据RF聚类分析结果显示,4个采样点小黄鱼幼鱼耳石近核心区元素比值大致可分为2个聚群(图6,A和B),大陈采样点73.68%的个体属于A,26.32%的个体属于B;舟山采样点82.35%的个体属于A,17.65%的个体属于B;吕四采样点的所有个体均属于B,射阳20%的个体属于A,80%的个体属于B。由此可看出,东海近岸的两个采样点主要属于A,黄海南部的两个采样点主要属于B(图6)。从两个聚群的元素比值来看,A群的元素比值中除了Mg比值均值高于B群外,Sr、Ba、Mn的平均比值低于B群(表6)。
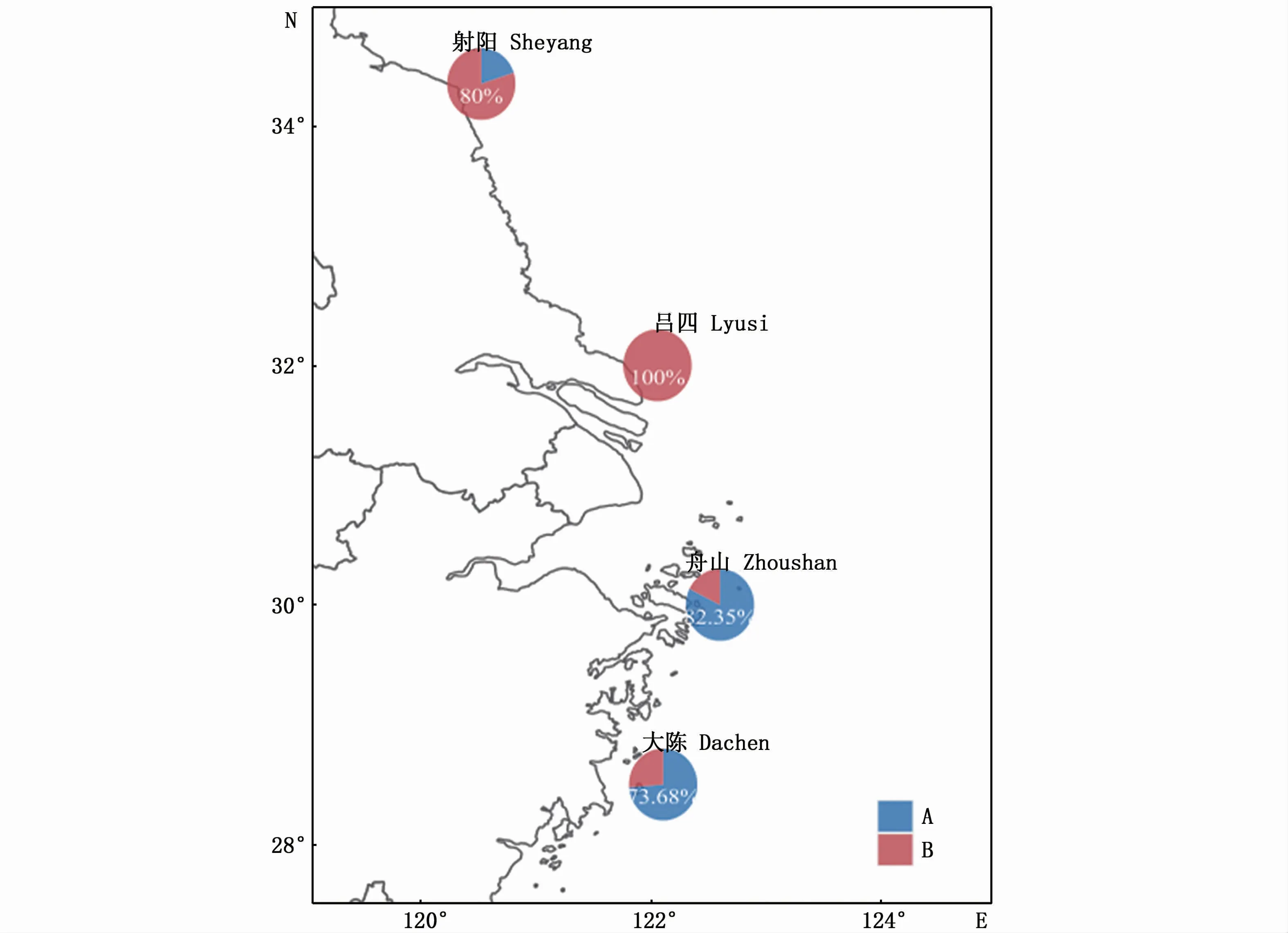
图6 RF聚类确定的各采样点小黄鱼幼鱼可能来源的饼图Fig.6 Pie charts representing the contribution of natal sources of Larimichthys polyactis identified by RF clustering

表6 RF分类确定的2个聚类组的耳石近核区元素比值(均值±方差)Tab.6 Mean element value for two natal clusters identified by RF clustering(mean±SD)
3 讨论
3.1 群体空间连通性分析方法比较
研究表明,不同的数据分析方法会影响到耳石元素的分类准确度,随机森林(RF)对于不同类型数据的分析性能要好于线性判别函数分析(LDFA)、二次判别函数分析(QDFA)、人工神经网络(ANN)等方法,而且对数据的分布假设要求不高[22,27]。因为无需确定采集样本出生源基准,幼鱼耳石元素无监督聚类学习方法似乎非常适合于仔稚鱼扩散的研究,该聚类方法不能直接提供关于不同幼鱼来源位置的信息,但能够推断出这些幼鱼出生源的数量[26],比如在北海,应用该聚类方法确定了海玉筋鱼(Ammodytes marinus)的4个出生源[13]。同时,无监督的RF聚类方法对于连续多元数据的要求以及分布假设均比较宽泛,很适合应用于耳石微化学数据的分析。GIBB等[26]认为,虽然有监督的学习方法也很好,但是获得具有代表性的出生源基准的难度很大,为了获取某种鱼的出生源基准样本就必须开展大规模的专业调查。因此要在接下来的研究中确定小黄鱼仔稚鱼的出生源,评估环境对早期生活史阶段的扩散、移动距离和范围的影响,为定量确定不同产卵场对小黄鱼资源补充的贡献以及小黄鱼种群的洄游提供科学依据。
3.2 小黄鱼幼鱼群体的空间连通性
采样点间小黄鱼幼鱼耳石边缘元素的分类准确度(63%)与徐浩等[19]使用耳石核区元素的差别成功率(70.6%)较为相似,不同海域不同种类耳石边缘元素的空间分类准确度变化较大,耳石微化学分类准确性一般处于中等水平(60% ~80%),如北海海玉筋鱼为48.8%[13]~80%[26],大西洋圆鲭(Scomber colias)为80%[11],大西洋棘白鲳(Chaetodipterus faber)为76%[10],葡萄牙沿海细鳞绿鳍鱼(Chelidonichthys lucerna)为73%[8],北海鳎(Solea solea)为79%[24]等。本研究中随机森林分析的结果显示,采样点间小黄鱼个体分配准确度范围为50%~75%,且分配准确度较低的都出现在相近的采样点上,比如射阳小黄鱼样品中有33.3%的样品被分配到吕四采样点,舟山采样点有25%的样品被分配至大陈采样点。空间上不相邻的采样点间的分配准确度均较高,比如射阳与舟山、大陈采样点之间均不存在着交叉的现象,这表明了不同海域小黄鱼幼鱼的扩散、移动范围是有限的,随着空间距离的增加,不同海域小黄鱼幼鱼群体间空间连通性越来越表现出相互隔离的趋势。从耳石边缘各元素比值对小黄鱼幼鱼RF分类准确度的贡献来看,Mn∶Ca的重要性最高,其次为Ba∶Ca,而Sr∶Ca的重要性最低,这表明小黄鱼幼鱼空间差异主要由Mn∶Ca、Ba∶Ca等元素比值所驱动。
耳石中Mn为Ca的可替换元素,也是基质蛋白辅助因子,反映了环境(浓度和盐度)和生理(个体发生和生长)的历史[28],但环境因素的影响还不清楚[10]。溶解态的Mn在水环境中迅速沉淀,导致海洋沉积物中Mn的浓度比海水中高出几个数量级,沉积物在海流的作用下重新悬浮[10],可能成为小黄鱼幼鱼的饵料。因此,Mn∶Ca的空间差异不一定表示海水化学组成的不同,可能反映的是小黄鱼幼鱼摄食生境的异质性。耳石边缘Ba∶Ca的空间差异可能归因于环境水体中Ba浓度的不同,Ba等微量元素渗入耳石的比例与周围环境浓度成正比[28],近岸海域Ba浓度的影响因素包括了陆地径流、流域地质、水体污染、潮汐变化、河口的混合作用以及地下水的输入等[7,10]。
鱼类耳石不同位点的元素比值与其所栖息环境的化学组成的相关性较高[3,28],耳石边缘的化学元素往往反映了采样时栖息海域的海水化学组成[2,29],在河口、沿海海域由于流域水质的局部变化,水化学组分常存在着广泛的空间变化(如潜在的地质、沉淀物、径流、微生物过程、人为影响、以及不同组成成分水团混合)[3,7]。东海和黄海南部沿海小黄鱼幼鱼耳石边缘4种元素比值除了吕四与舟山采样点差异不显著外,其他采样点间均存在显著差异。小黄鱼幼鱼耳石边缘元素比值的差异性反映了小黄鱼幼鱼采样海域海水化学组成的差异性,舟山和吕四采样点位于长江口的南北侧,它们均受到“长江口冲淡水双向扩展”的影响[30],这两个采样点小黄鱼耳石边缘的元素比值具有更高的相似性(图3);而其他两个采样点距离较远,影响射阳和大陈采样点附近海域的流系不同,可能导致这两个海域的化学组成差异性较高,从而使栖息在这两个海域的小黄鱼幼鱼耳石边缘化学元素具有较大的差异性。
3.3 小黄鱼幼鱼出生源
确定产卵场出生源对育幼和索饵群体的贡献将有助于更好理解种群结构动态[24],海洋鱼类仔稚鱼的死亡率很高,很难追踪到幼鱼阶段的所有可能仔鱼来源,因此一些研究通过对仔稚鱼生活史阶段形成耳石的化学组成进行聚类分析,进而推断仔鱼来源的数量[2,26]。当受精卵在孵化过程中,海水中的Sr、Ba等化学元素被吸收整合到鱼类胚胎耳石中(即孵化前区域),从而胚胎耳石中产生反映出生地环境的化学特征[12,31]。依据小黄鱼幼鱼耳石近核区的4种元素比值的RF分类结果,4个采样点的小黄鱼幼鱼样品主要来自两个类型产卵场,分别为东海近岸(A)和南黄海近岸(B)的产卵场,这也证实了东海近岸、黄海南部近岸存在着两个小黄鱼种群的产卵场[15]。由于小黄鱼产卵地(出生源)准确海域尚不清楚,目前尚无法确定聚类在多大程度上反映不同的出生源[26]。
东海近海(A)的小黄鱼耳石近核区的Sr∶Ca、Ba∶Ca和Mn∶Ca低于黄海南部近海(B),Mg∶Ca则高于黄海南部(表6)。小黄鱼幼鱼耳石近核区的元素比值中除了Mn∶Ca无空间差异外,其他元素比值表现出不同程度的空间差异。采样点之间的Ba∶Ca空间差异性最大,主要是Ba驱动了小黄鱼出生源的空间差异。多数情况下,耳石上的Sr、Ba 丰度与盐度等水环境因子相关[3,28,32-33],Mg则与鱼类生长发育相关[28],表明了东海近海和黄海南部小黄鱼产卵场的化学组成存在不同,这也就证实了两个海域都存在着自身的小黄鱼产卵种群[3,7,34]。两个产卵场个体的多种元素也存在明显差异,说明两个产卵场出生的小黄鱼基本上都是在本海域生长发育,并逐渐向产卵场海域附近的育幼场和索饵场扩散洄游[15]。
