宰后成熟期间能量物质、pH值和肌原纤维小片化指数对秦川牛肉嫩度的影响及其机理
2022-07-02李亚蕾罗瑞明张杏亚马思丽
罗 辉,杨 波,李亚蕾*,罗瑞明,张杏亚,马思丽,姬 琛
(宁夏大学食品与葡萄酒学院,宁夏 银川 750021)
秦川牛是陕甘宁地区优势畜种,近5 年存栏量在300万 头左右,年均出栏量有100万 头左右,年产肉量在50万 t以上,具有一定地域特色。秦川牛肉质细致、肉味浓郁、营养全价,肉用性能好。其背最长肌具有瘦肉比例高、大理石花纹丰富等特点,可用于制作高档牛排,提升附加值,因此本研究选用秦川牛背最长肌为研究对象。
目前肉类行业无论是在活体动物中还是在屠体中缺乏一种快速、客观的技术来预测评估牛肉贮藏过程中的品质指标变化。解决这一技术难题能提高牛肉行业的效率,并提高工业生产更高质量肉制品的能力。为了实现这一目标,必须确定影响肉质性状变化的潜在机制,为此,研究肉质的科研工作者普遍认为,在试图开发预测肉品质的技术之前,首先要揭示负责调控表观变化的基因和蛋白质网络。这样便于准确地管理肉类质量性状,并为生产者开发基因选择方案,以优化种牛品质。在过去的几十年里,人们为实现这一目标已经做出了许多尝试,特别是利用蛋白质组学技术来研究肉类品质变化,以更好地揭示牛肉嫩度的潜在机制。Sawdy等研究表明牛背最长肌贮藏7 d后嫩度与肌球蛋白重链断裂有关,在肌肉嫩化过程中,肌球蛋白轻链1、2和重链1的表达量明显降低;李升升利用非标记定量(label-free quantitative,LFQ)蛋白质组学技术研究牦牛平滑肌贮藏过程嫩度的变化机理,结果表明SLC25A4、Calpastatin、TPM3、UQCRC1、LMOD1、THBS4和ZYX 7种蛋白可能是牦牛平滑肌贮藏期嫩度变化的指示蛋白;Jia等也利用LFQ蛋白质组学方法对牛胸最长肌肌浆蛋白进行研究发现,在贮藏的前期peroxiredoxin-6表达量高的组随着贮藏时间的延长嫩度也会显著增高,由此可以通过检测活体组织或屠宰后不久牛胸最长肌中peroxiredoxin-6含量来预测嫩度变化,因此peroxiredoxin-6可作为肉类嫩度的潜在蛋白标记物;Poleti等利用高清晰度质谱仪(high definition mass spectrometry,HDMS)检测Nellore牛背最长肌蛋白表达量,发现118种蛋白质的丰度发生了显著变化,经过生物学分析发现这些蛋白主要富集在丙酮酸代谢、分解代谢、肌动蛋白结合、细胞骨架蛋白结合和钙离子结合等途径,由此推断能量代谢和细胞骨架结构的变化可能影响到嫩度。在几十年的研究中,人们利用蛋白质组学技术不断揭示了调控嫩度变化的指示蛋白,调控嫩度的蛋白质图谱也在逐渐完善,嫩度变化预测技术也有了部分理论的支撑。但不同牛品种之间表现也会有较大差异,秦川牛背最长肌嫩度变化机理报道鲜有发现,本研究希望能进一步完善这一理论基础。
本研究中利用4维-LFQ(4 dimension-LFQ,4D-LFQ)蛋白质组学技术筛选秦川牛差异蛋白,4D-LFQ蛋白质组学技术相对于3维蛋白质组学在质荷比(/)、离子强度和保留时间3个维度添加了离子淌度(mobility)。离子淌度为牛肉肌肉组织这种复杂样品体系的蛋白组检测提供了更多可能,可根据离子的截面和形状进一步对蛋白进行分离鉴定,使蛋白组检测数据更为准确、全面。
1 材料与方法
1.1 材料与试剂
25 月龄秦川公牛 甘肃省平凉市泾川县旭康食品有限责任公司。
蛋白酶抑制剂 德国Calbiochem 公司;胰酶美国Promega公司;乙腈 美国Fisher Chemical公司;甲酸瑞士Fluka公司;三氟乙酸、尿素、碘代乙酰胺、二硫苏糖醇、三乙基碳酸氢铵、三氟乙酸 美国Sigma公司;BCA试剂盒 上海碧云天生物技术有限公司;pH校准液上海仪电科学仪器股份有限公司。
1.2 仪器与设备
超声波细胞破碎机 那艾精密仪器(上海)有限公司;205便捷式pH计 德国德图集团;MiniVac Alpha冷冻离心机 美国Scan speed公司;timsTOF Pro质谱仪、NanoElute超高效液相色谱仪 德国Bruker公司;Forma 994超低温冰箱 美国Thermo公司;TA-XT plus质构仪英国Stable Micro Systems公司。
1.3 方法
1.3.1 样品采集
选用3 头生长发育正常、体壮无病、25 月龄左右的去势秦川公牛,屠宰后取牛两侧背最长肌并分割为质量约(100f10)g的肉样,随机分成5 组并置于透气性为23.5 g/(m•d)的聚乙烯保鲜袋中。立即取其中一组样品检测各项指标(视为宰后第0天样品),将蛋白质组学、能量物质检测和剪切力检测实验所用样品置于液氮中保存。其他4 组贮藏于0~4 ℃的冰箱中,分别在第2、4、6、8天时取出,测定相应指标。
1.3.2 能量物质含量的测定
称取50 mg样品置于离心管中,加入4 ℃预冷的1 mL 混合液(含0.6 mol/L高氯酸和1 mmol/L乙二胺四乙酸),匀浆2 min后4 ℃、10 000h条件下离心10 min,取700 µL上清液加入0.2 mL 1 mol/L NaOH。摇匀后反应5 min,取0.5 mL混合溶液用聚碳酸酯滤膜过滤后于-80 ℃冰箱保存备用。采用高相液相色谱法检测ATP、ADP、AMP、NADH的含量。流动相为220 mmol/L硫酸钾缓冲液(含10%甲醇并使用四丁基氢氧化铵调节至pH 6.5),等度洗脱,流速为0.8 mL/min,检测波长为254 nm。
1.3.3 剪切力测定
参考文献[16]测定样品剪切力,以表征样品的嫩度。取出贮藏在0~4 ℃环境下的肉样,插入热电偶温度计至肉样中央并置于蒸煮袋中,在85 ℃水浴锅中加热,待肉样中心温度达80 ℃时取出,流动自来水冲洗蒸煮袋将肉样冷却至室温,切除表面,避开筋腱等部分,平行于肌纤维方向用口径为1.27 cm的取样器切割制作肉柱,用质构仪“V”型剪切刀检测肉柱,剪切刀垂直于肌纤维方向,剪切速率与剪切距离分别为1.5 mm/s、40 mm,每个肉样检测3 次,结果取平均值。
1.3.4 pH值测定
参照文献[17]的方法,pH计使用前用pH校准液校正,取4 cmh4 cmh4 cm样品(60 g左右)背最长肌,探头沿肌纤维方向插入肉样内部,每个样品取5个点测定,结果取平均值。
1.3.5 4D-LFQ蛋白质组学技术检测蛋白质量浓度
参照文献[18]的方法提取蛋白质。4 ℃预冷后的研钵放入适量样品和液氮研磨成粉末状,加样品4 倍体积的裂解缓冲液(等体积的8 mol/L尿素和1%蛋白质酶抑制剂)。超声裂解后移入离心管,4 ℃、12 000h条件下离心10 min,取上清液利用BCA试剂盒测定蛋白质量浓度。
参照文献[19]的方法对提取的蛋白进行胰酶酶解。取上述蛋白溶液加入二硫苏糖醇溶液,调节蛋白质浓度为5 mmol/L。56 ℃避光环境下还原30 min再加入100 mmol/L碘代乙酰胺溶液,将蛋白质浓度调节至1 mmol/L,25 ℃避光环境下静止12 min。用四乙基溴化铵溶液将样品中的尿素稀释到2 mol/L以下,按照蛋白质量的2%添加胰酶,37 ℃酶解12 h,再按照蛋白质量的1%添加胰酶,37 ℃酶解4 h。
液相色谱-质谱联用分析:使用NanoElute超高效液相系统进行肽段分离,流动相A为0.1%(体积分数)甲酸水溶液,用来溶解蛋白片段;流动相B为0.1%(体积分数)甲酸-乙腈溶液;流速为300 nL/min。梯度洗脱程序为:0~3 min,4%~6% B;3~70 min,6%~25% B;70~82 min,25%~32% B;82~87 min,32%~80% B;87~90 min,80%~100% B。肽段分离后注入到capillary离子源中进行电离,电压设置为1.4 kV,使用飞行时间质谱对肽段母离子及其二级碎片进行检测和分析,设置扫描范围100~1 700/。通过平行累积串行碎裂模式(parallel accumulation serial fragmentation,PASEF)进行数据采集。将采集后一级质谱用10 次PASEF模式采集母离子电荷数在0~5范围获得二级谱图,为了避免母离子的重复扫描串联质谱扫描的动态排除时间设置为24 s。
1.3.6 数据库检索
二级质谱的数据检索采用Maxquant(v1.6.6.0)软件进行。各项参数设置如下:选择Bos_taurus_9913_PR_20190915.fasta(37 948 条序列)为数据库,加入常见的污染库排除污染蛋白质的影响,加入反库特征信息匹配排除计算随机匹配造成的假阳性率(false discovery rate,FDR)影响;酶切方式为Trypsin/P,Main search与First search的二级碎片离子的质量误差容忍度为0.04 Da,一级母离子质量误差容忍度设为40h10Da;肽段最大修饰数、漏切位点数和肽段最小长度分别设置为5、2、7。固定修饰为半胱氨酸烷基化,可变修饰为蛋白N端的乙酰化和甲硫氨酸的氧化。肽匹配图谱鉴定的FDR和蛋白鉴定均设置为1%。
1.3.7 生物信息学分析
筛选出的差异蛋白质利用软件ClusterProfiler(3.4.4)进行生物信息学分析,比对后获得差异基因的基因本体(gene ontology,GO)(包括生物学过程(biological process,BP)、细胞组分(cellular component,CC))功能信息。利用String 10.0数据库(http://string-db.org/)和Cytoscape 3.6.1软件绘制差异蛋白相互作用网络图。
1.3.8 肌原纤维小片化指数的测定
称取肉样1 g使用冷冻研磨仪搅碎后置于离心管中,加入10 mL肌原纤维小片化指数(myofibril fragmentation index,MFI)缓冲液(含100 mmol/L氯化钾、11.2 mmol/L磷酸氢二钾、8.8 mmol/L磷酸二氢钾、1 mmol/L乙二醇双四乙酸、1 mmol/L氯化镁、1 mmol/L叠氮化钠),4 ℃、1 200 r/min匀浆3 min,然后4 ℃、1 000h离心15 min收集上清液;加入10 mL MFI缓冲液重悬沉淀并按上述步骤再次匀浆后离心,收集上清液;再加入5 mL预冷MFI缓冲溶液使沉淀充分悬浮后用200 目筛过滤,再用5 mL MFI缓冲液清洗离心管,然后用200 目筛过滤,合并上清液和滤液,并采用考马斯亮蓝法测定混合溶液的蛋白质量浓度。用MFI缓冲液调整蛋白质量浓度为0.5 mg/mL左右,在595 nm波长处测定吸光度,每个样品重复3 次,取平均值带入下式计算MFI。

1.4 数据处理及分析
所有数据均以平均值±标准差表示,采用SPSS 25软件进行单因素分析,采用邓肯检验进行显著性分析,<0.05表示差异显著,<0.01表示差异极显著。采用Origin 2018软件作图,采用微生信在线平台(http://www.bioinformatics.com.cn/)进行秦川牛肉背最长肌差异蛋白数据分析并作图。采用SPSS 25软件分别对差异蛋白表达量与剪切力、能量物质含量进行Pearson相关性分析,筛选出与嫩度相关的蛋白质。
2 结果与分析
2.1 秦川牛背最长肌贮藏过程中ATP、ADP、AMP、NADH含量的变化
采用高效液相色谱法测定了秦川牛背最长肌不同贮藏期能量物质含量。如表1所示,随着贮藏时间的延长,ATP的含量呈显著下降趋势(<0.05),由0 d时的(10.15f0.60)μmol/g下降至8 d时的(0.50f0.20)μmol/g,其中在0~2 d下降速率较快,在2~8 d下降速率较为缓慢,第8天时基本检测不出ATP,ADP、AMP的含量在整个贮藏期(0~8 d)均呈现下降趋势。秦川牛宰后肌肉细胞中ATP进行着不可逆的降解过程,由于ATP含量的减少,ADP、AMP含量随之减少。贮藏过程中NADH含量变化可反映组织细胞中氧化还原和能量变化状态。实验结果中NADH含量逐渐降低,说明宰后肌肉细胞发生变化,使细胞呼吸功能失调,氧化磷酸化进程受阻,引起NADH含量出现显著下降的现象。

表1 秦川牛背最长肌不同贮藏时间下ATP、ADP、AMP、NADH含量变化Table 1 Changes in ATP, ADP, AMP and NADH contents in longissimus dorsi muscle of Qinchuan cattle with storage time
2.2 秦川牛背最长肌贮藏过程剪切力的变化
嫩度是影响秦川牛肉口感的重要品质属性,也是影响消费者购买欲的重要指标。咀嚼破碎肉制品时口腔所受的抵抗力大小可称为肉制品的嫩度,实验结果以剪切力表征嫩度。如图1所示,秦川牛肉在贮藏过程中剪切力总体呈先上升后下降的趋势(<0.01),其下降幅度大于上升幅度。第0天时剪切力为(120.25f5.15)N,在第4天时达到最大值,为(157.94f2.53)N,第8天时又降至(93.41f5.08)N,通过以上剪切力的变化可以看出秦川牛背最长肌在贮藏中的嫩度变化情况,在0~4 d时逐渐降低,第4天时到达最低点,之后逐渐升高,直至第8天。
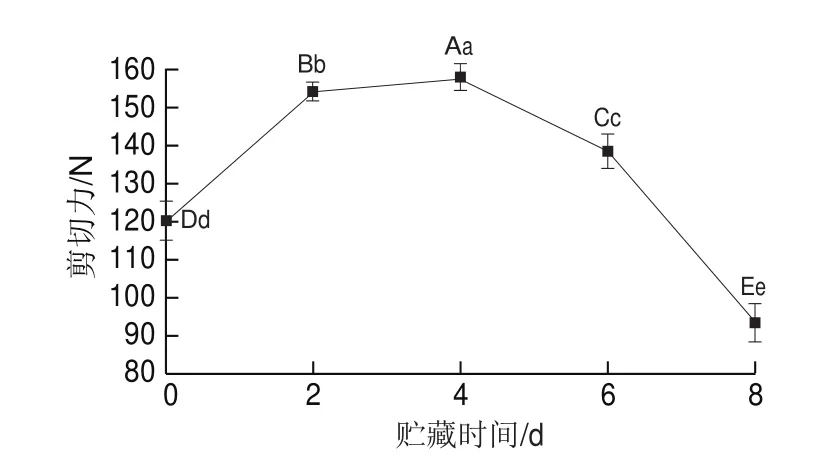
图1 秦川牛背最长肌贮藏期间剪切力变化Fig.1 Changes in shear force of longissimus dorsi muscle with storage time
2.3 秦川牛背最长肌贮藏过程中pH值变化
秦川牛背最长肌在贮藏过程中pH值变化情况如图2所示,宰后0 d时pH值为6.78f0.02,在0~2 d贮藏期内下降速率较快,2~4 d贮藏期下降速度放缓后到达最低点5.37f0.03,4~8 d时呈缓慢上升趋势,8 d时上升至5.50f0.04。整体下降幅度大于上升幅度。无氧条件下,机体通过无氧糖酵解产能的同时产生乳酸;此外,宰后ATP水解产生磷酸,这些酸性物质长期积累并无法分解与转运,使得肌肉组织的pH值迅速下降。

图2 秦川牛背最长肌贮藏期间pH值的变化Fig.2 Changes in pH in longissimus dorsi muscle of Qinchuan cattle with storage time
2.4 蛋白质组学数据分析及关键蛋白质筛选结果
实验中采用4D-LFQ对贮藏0、4、8 d的样品蛋白质丰富度变化进行定量分析,利用Maxquant(v1.6.6.0)软件对所得的蛋白质二级结构进行质谱检索,共检索确定蛋白1 149个,设置1.200 倍为差异显著倍数,共筛选出120个差异显著的蛋白质,为进一步确定数据的可信度,对差异蛋白进行了聚类分析与主成分分析。
秦川牛背最长肌差异蛋白丰富度层次聚类分析如图3A所示,各组的3个平行样品均可聚为一类,说明样品重复性较好,且不同贮藏时间样品差异明显;同时,4、8 d的样品距离更近,与0 d样品相区分。差异蛋白的主成分分析结果如图3B所示,PC1、PC2的累计贡献率达到72.23%,因此这两个主成分可较好地代表差异蛋白表达的信息。PC1能将0 d与其他样品区分开,PC2能将4 d与8 d样品区分开。综上可判断,秦川牛背最长肌差异蛋白0、4、8 d样品重复性较好且有显著差异,具有一定代表性和可信度。


图3 秦川牛肉背最长肌差异蛋白整体数据分析结果Fig.3 Analysis of differential proteins between longissimus dorsi muscle of Qinchuan cattle stored for different periods
采用SPSS 25软件对差异蛋白表达量与剪切力进行Pearson相关性分析,筛选出与嫩度变化相关的11种差异蛋白,如表2所示。
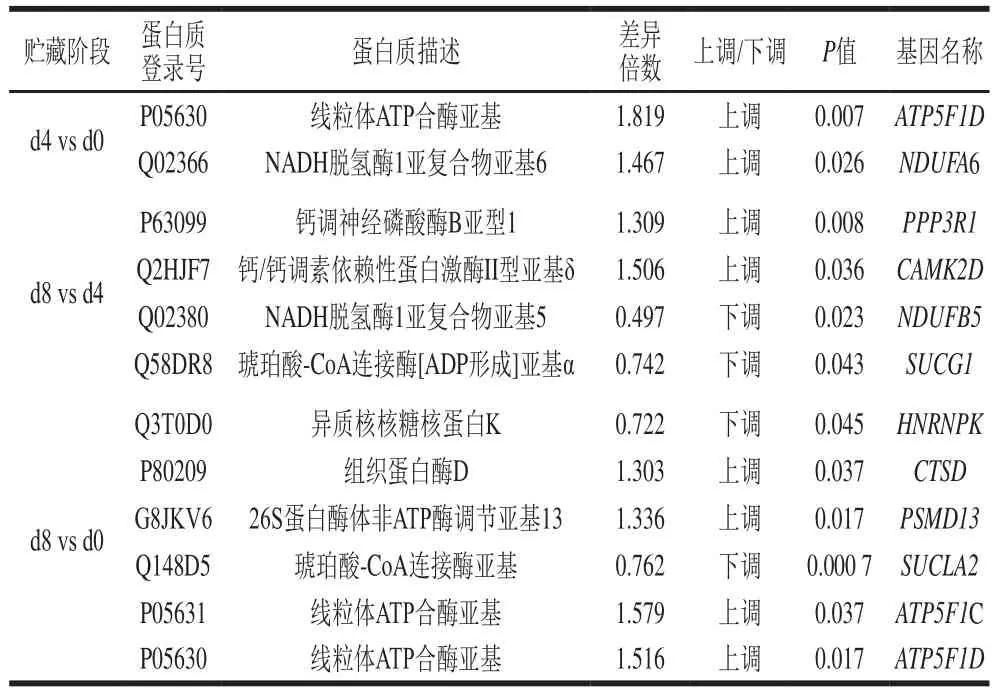
表2 秦川牛背最长肌与嫩度相关的差异蛋白Table 2 Differential proteins related to beef tenderness in longissimus dorsi muscle of Qinchuan cattle
2.5 差异蛋白GO注释结果
对秦川牛肉背最长肌筛选出的差异蛋白进行GO分析后,所涉及到的生物过程与细胞组分前10的条目情况如图4所示,生物过程主要集中在氧化还原过程(ATP5F1D、NDUFA6、NDUFB5、SUCLA2)、线粒体组织(ATP5F1D、NDUFA6、NDUFB5)、NADH脱氢酶符合体组装(NDUFA6、NDUFB5)、细胞呼吸(ATP5F1D、SUCLA2)等过程;细胞组分主要集中在催化复合物(NDUFA6、PPP3R1、NDUFB5、HNRNPK、PSMD13)、线粒体蛋白质复合物(ATP5F1D、NDUFA6、NDUFB5、ATP5F1C)、细胞器内膜(ATP5F1D、NDUFA6、NDUFB5、ATP5F1C)、肌纤维膜(PPP3R1、CAMK2D)等。
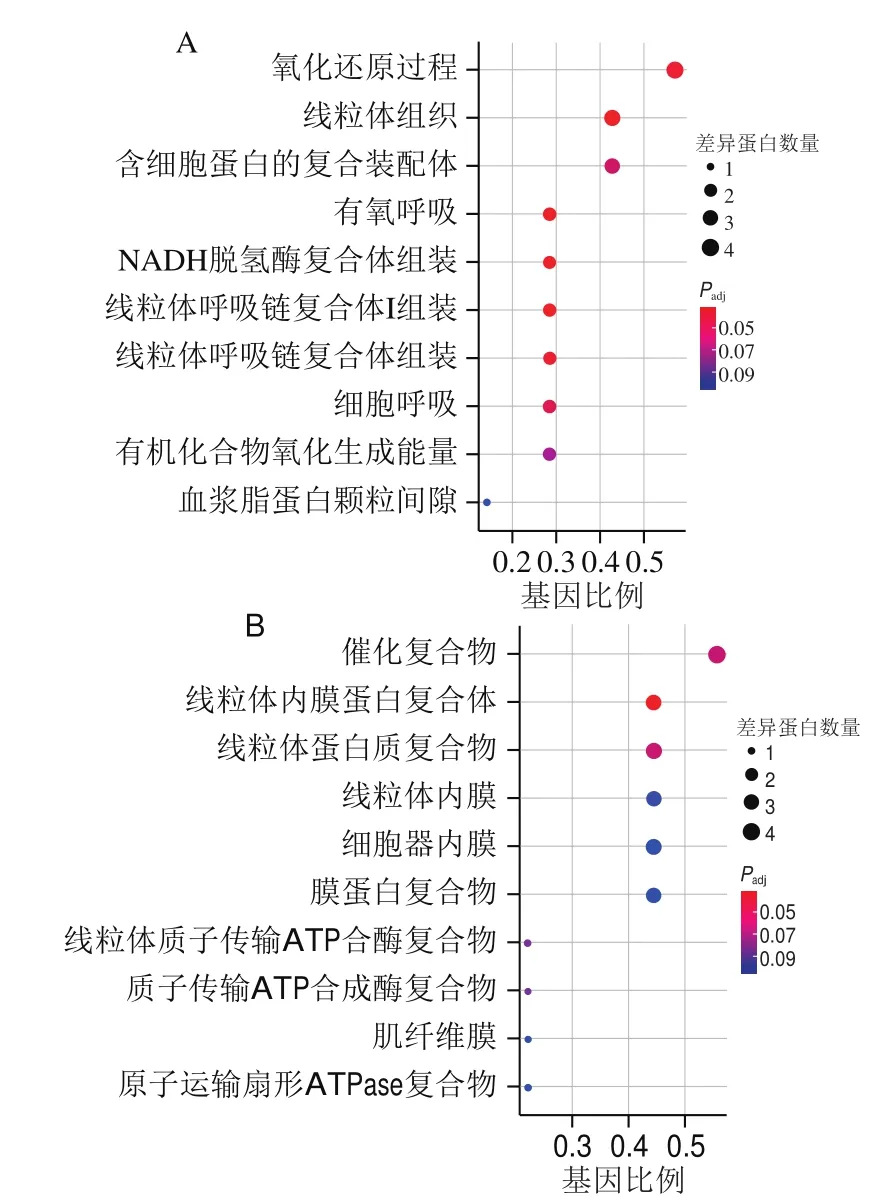
图4 秦川牛肉背最长肌差异蛋白GO分析结果Fig.4 Gene ontology (GO) analysis of differential proteins in longissimus dorsi muscle of Qinchuan cattle
2.6 差异蛋白相互作用网络图
图5是利用Cytoscape 3.6.1软件和String 10.0数据库制作的差异蛋白相互作用网络图。此网络图聚类系数为0.691,蛋白质相互作用富集值为1.39h10。如图5所示,ATP5D、ATP5C1、SUCLA2、SUCLG1、NDUFB5、NDUFA6的相互作用较为强烈,通过GO分析发现在氧化还原过程,线粒体组织条目有较密切互作关系,分析发现以上6种蛋白质组成的集团均参与调解宰后能量代谢,最终通过ATP含量和pH值变化影响嫩度变化;CAMK2D与PPP3R1的相关作用较为强烈,分析发现两者具有协同关系,共同参与细胞凋亡,促进MFI的增大,肌原纤维受到破坏,最终使得嫩度降低;CTSD、HNRNPK、PSMD13暂未与其他蛋白发现相互作用。

图5 差异蛋白相互作用网络图Fig.5 Interaction network between differential proteins
2.7 pH值、能量物质含量及相关差异蛋白对嫩度的影响
利用SPSS 25软件对表2中11种蛋白质的表达量与能量物质含量进行Pearson相关性分析,筛选出6种与能量变化相关的差异蛋白质,结果如表3所示,它们分别为ATP5F1D、ATP5F1C、NDUFA6、NDUFA6、SUCLA2、SUCG1。
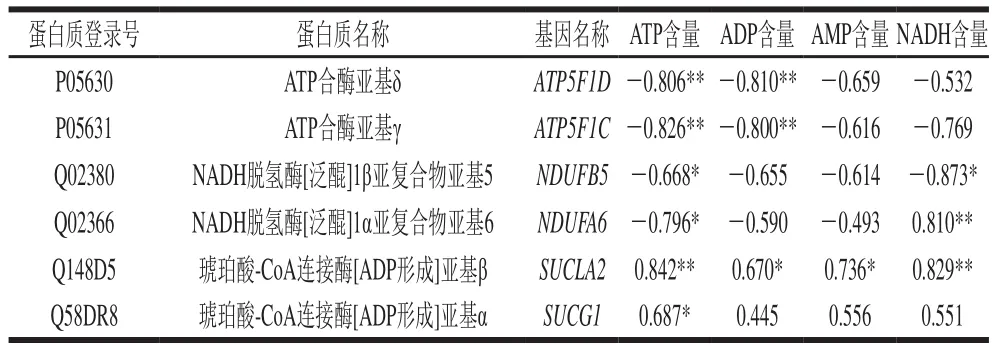
表3 差异蛋白质表达量与能量物质含量的相关性Table 3 Correlation between expression of differential proteins and energy substance contents
表2中ATP5F1D是ATP合酶的亚基δ,ATP5F1C是ATP合酶亚基γ,这两种蛋白质参与ATP合成酶的组装,是ATP合酶5个亚基中的2个,对ATP合酶催化活性有重要作用,承担着离子传递的作用。ATP含量合酶参与ATP的合成与消耗,所以ATP合酶直接影响机体细胞的供能和耗能,其表达量也会直接反映细胞中能量需求情况和三羧酸循环产能的情况。由表3可知,ATP5F1D、ATP5F1C表达量与能量基础物质ATP含量呈极显著负相关(<0.01)。表2中NDUFA6为NADH脱氢酶的附件亚基。NADH脱氢酶参与线粒体内膜上的呼吸作用,对电子从NADH传递给辅酶Q有催化作用,NDUFA6表达量与能量基础物质ATP含量呈显著负相关(<0.05),与NADH含量呈极显著正相关(<0.01)。ATP5F1D和NDUFA6在0~4 d的贮藏过程中均有高水平的表达,ATP5F1D上调1.819 倍,NDUFA6上调1.467 倍。两者均是糖酵解过程中关键酶的组件,其高水平的表达说明机体细胞在切断氧气供应时,能提高参与产能的酶含量,利用无氧糖酵解过程产能,在此过程中大量产生乳酸,使得肌肉pH值下降。
琥珀酰辅酶A合成酶在柠檬酸循环中发挥作用,在琥珀酸硫激酶的催化下,琥珀酰辅酶A高能键水解生成琥珀酸,同时使GDP磷酸化生成GTP,GTP再与ADP作用生成ATP。表2中SUCLA2作为琥珀酸-CoA连接酶[ADP形成]亚基β,提供酶的核苷酸特异性,并与底物琥珀酸结合,而辅酶A和磷酸的结合位点在α亚基。而筛选出SUCG1就是琥珀酸-CoA连接酶[ADP形成]亚基α。通过表3也可以看出,SUCLA2的表达量与ATP、NADH含量呈极显著正相关(<0.01),与ADP、AMP显著正相关(<0.05);SUCG1表达量与ATP含量呈含量显著正相关(<0.05)。表2中NDUFB5与NDUFA6同为NADH脱氢酶的附件亚基,在磷酸氧化过程中有着同等作用,从表3中也可以看出NDUFB5的表达量与ATP、NADH含量均呈显著负相关(<0.05)。从表2可以看出,SUCLA2的表达量在0~8 d调整0.762 倍,NDUFB5的表达量在4~8 d调整0.497 倍,SUCG1的表达量在4~8 d调整0.742 倍。SUCLA2、NDUFB5、NDUFA6是在三羧酸循环、呼吸电子链传递、通过化学渗透耦合进行ATP合成、解偶联蛋白质中起关键作用的蛋白,在4~8 d的贮藏期内均有下调趋势,说明无氧糖酵解过程可能会受到抑制,能量和乳酸的生成量相应减少。
秦川牛屠宰后血液流动停止,肌肉细胞通过调节以上蛋白表达量,启动糖酵解过程产能,本实验数据显示,0~4 d时秦川牛肉的ATP的含量减少至(1.20f0.45)μmol/g,为初始状态的11.82%,在能量物质ATP下降到宰前初始含量的20%以下时,肌纤维发生交联,肌肉会出现僵直现象,由图1可知,在0~4 d贮藏期秦川牛肉剪切力也随之上升,4 d后能量物质含量下降速率有所减慢,对肌肉收缩的影响可能会减小。无氧糖酵解产生能量的同时产生乳酸;此外宰后ATP水解产生磷酸,这些酸性物质,使得肌肉组织的pH值迅速下降。4 d时pH值为5.37f0.03,低pH值改变了质子的稳态,降低了肌原纤维蛋白之间的静电斥力,导致肌肉纤维不可逆侧向收缩(肌节延伸),使得肉嫩度变差;低pH值使纤维蛋白暴露出更多水解位点,有助于水解酶的水解作用;低pH值会降低溶酶体膜的稳定性,使得肌肉细胞释放内源水解酶,同时pH值的降低会使肌肉细胞处于酸性环境,打破细胞中的离子平衡,促进钙离子的释放,Ca浓度的提高会激活钙蛋白酶,降解纤维蛋白,促使嫩度升高,但pH值的降低也会使得-钙激活酶发生自溶现象,降低酶活性,降低肌原纤维蛋白的水解速率;也有研究表明pH值小于5.8时,组织酶D对水解鸵鸟纤维有促进作用,由此可判断pH值会影响到内源酶的催化活性,对肌原纤维蛋白的降解速率产生一定影响。通过图6也可以看出,剪切力和ATP含量、pH值均呈显著负相关(<0.05)。综上可推测,0~4 d能量物质含量的减少和pH值的降低会使得肌肉的僵直,肌肉嫩度降低,4~8 d能量物质含量变化较小,对肌肉的影响也会相应降低,长期低pH值下的各种途径综合作用对嫩度提高的速率和嫩度提高的程度均有影响。
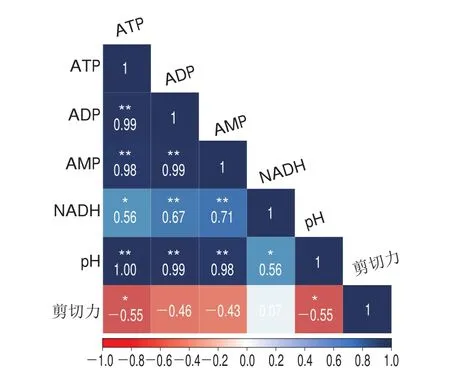
图6 秦川牛背最长肌能量物质含量、pH值和剪切力之间的相关系数Fig.6 Correlation coefficients between energy substance contents, pH and shear force
2.8 与MFI相关的差异蛋白对宰后秦川牛肉嫩度变化的影响
秦川牛肉在贮藏过程中MFI变化情况如图7所示,在整个贮藏过程中MFI呈显著上升趋势(<0.05),由0 d时28.50f5.1持续增长至8 d时的99.98f1.20,变化了250.81%。此结果与李升升对牦牛平滑肌嫩度形成机理研究中的MFI增加趋势一致。

图7 秦川牛背最长肌不同贮藏时间MFI变化情况Fig.7 Changes in MFI of longissimus muscle of Qinchuan cattle with storage time
利用SPSS 25软件将表2中11种与嫩度相关的蛋白质表达量与与MFI进行相关性分析,筛选出5种(CTSD、PSMD13、PPP3R1、CAMK2D、HNRNPK)与MFI有关的关键蛋白质,相关性分析结果如表4所示。

表4 秦川牛背最长肌贮藏期间剪切力、MFI与差异蛋白含量相关系数Table 4 Correlation coefficients between shear force, MFI and differential protein contents
CTSD为组织蛋白酶D,位于溶酶体,pH值降低等因素会影响溶酶体膜稳定性,使得CTSD被释放,后经过ADAM30D切割后可激活活性,在蛋白质的降解与细胞凋亡中起作用,CTSD在0~8 d中的表达量上调1.303 倍(表2),CTSD表达量与剪切力呈显著负相关(<0.05),与MFI呈显著正相关(<0.05),CTSD的高表达可能会出现肌肉细胞更高水平的自噬。PSMD13是26S蛋白酶体非ATP酶调节亚基13,26S蛋白酶体非ATP酶可以去除可能损害细胞功能的错误折叠或受损的蛋白质,还可以通过去除不再需要的功能蛋白质,维持蛋白质动态平衡,同时参与细胞凋亡的进程,PDMD13在0~8 d中上调1.336 倍(表2),PSMD13表达量与剪切力呈极显著负相关(<0.01),与MFI呈显著正相关(<0.05),PSMD13表达量发生变化会影响到细胞凋亡和受损蛋白质去除。HNRNPK是与mRNA结合的主要蛋白质,可能在核内不均一RNA(heterogeneous nuclear RNA,hnRNA)的核代谢中通过转录激活和抑制发挥作用,在糖基化中起促进作用,作为抑癌基因/的转录辅激活因子,在/和/诱导中起作用;在转录抑制方面,它通过与/诱导的非编码RNA(-)相互作用发挥作用,这种相互作用是诱导细胞凋亡所发生的,HNRNPK在0~8 d调整0.722 倍(表2),HNRNPK表达量与剪切力呈显著正相关(<0.05),与MFI呈显著负相关(<0.05),HNRNPK在贮藏期内的异常表达,会直接影响到肌细胞的凋亡进程。
研究中发现了两种关键蛋白质在细胞凋亡进程中具有协同作用。PPP3R1是钙调磷酸酶B亚基1型,是一种钙调节蛋白刺激的蛋白磷酸酶,提供细胞对Ca的敏感性,在4~8 d上调1.309 倍,提高了细胞对钙离子的敏感性。CAMK2D是钙/钙调素依赖性蛋白激酶II型亚基δ,CAMK2D通过靶向钙离子通道、转运蛋白参与细胞钙离子内流、肌浆网钙释放、肌浆网钙摄取,影响细胞的钙稳态和兴奋收缩偶联的调节,CAMK2D表达量在4~8 d上调1.506 倍。由表4可以看出,PPP3R1表达量与剪切力呈极显著负相关(<0.01),与MFI呈极显著正相关(<0.01);CAMK2D表达量与剪切力呈显著负相关(<0.05),与MFI呈显著正相关(<0.05)。初步判断PPP3R1和CAMK2D在细胞凋亡进程中具有协同作用,CAMK2D促进细胞对Ca的吸收,提高细胞中Ca的浓度;PPP3R1可提高细胞对钙离子的敏感性,Ca超负荷的积累是触发细胞凋亡的因素,这两种蛋白同时在4~8 d时高表达,共同作用促使细胞凋亡进程的发生,降低肌原纤维结构强度,使得嫩度提高。
MFI可直接反映肌纤维的降解程度,增大原因是由于Z线蛋白的降解和肌原纤维细胞凋亡。宰后无氧糖酵解导致pH值降低,使得细胞离子失衡,钙离子浓度升高;降低溶酶体膜与细胞膜的稳定性,释放并激活内源酶,CTSD、PSMD13、PPP3R1、CAMK2D、HNRNPK 5种参与细胞凋亡、蛋白质水解的蛋白丰富度发生了改变,加快了肌原纤维蛋白的水解与细胞凋亡的发生,促使MFI升高了250.81%,降低了肌原纤维骨架强度,使得秦川牛背最长肌嫩度升高。
2.9 能量物质、pH值、MFI对秦川牛肉背最长肌嫩度变化不同时间段产生影响的分析
秦川牛宰后能量和pH值变化较为迅速,对肌纤维收缩的影响较早,0~4 d时肌肉僵直对嫩度变化影响较大,4~8 d时随着能量物质趋于稳定,对肌肉收缩的影响减小,对肌肉嫩度的影响也会随之降低。低pH值与5种参与细胞凋亡有关的内源酶相互作用使得MFI升高,是长期积累的过程,当MFI升高到一定程度,肌原纤维骨架强度显著降低时,才会使得肌肉嫩度升高,由图1剪切力数据可以看出MFI作用主要集中在4~8 d时,所以MFI作用具有滞后性。
3 结 论
秦川牛肉在贮藏过程中6种蛋白(ATP5F1D、ATP5F1C、NDUFB5、NDUFA6、SUCLA2、SUCG1)通过呼吸电子链传递、通过化学渗透耦合进行ATP合成、解偶联蛋白质等途径影响宰后糖酵解过程,ATP含量降至初始状态(0 d)的11.82%,引起肌纤维发生交联,肌肉会出现僵直,造成肌肉剪切力增大,嫩度下降。能量物质含量宰后变化较快且后期稳定,所以对肌肉嫩度影响主要集中在0~4 d时。宰后酸性物质的积累造成肌肉pH值下降,导致肌原纤维蛋白之间的静电斥力减小,影响了肌肉收缩;参与细胞钙离子浓度、促使肌原纤维受体暴露、调节内源酶活性等过程使得嫩度升高。5种蛋白(PPP3R1、CAMK2D、HNRNPK、PSMD13、CTSD)通过肌纤维膜、膜蛋白复合物、细胞器内膜、钙信号通路调控肌纤维细胞的凋亡和蛋白的水解,造成肌原纤维结构被破坏,使得MFI逐渐升高,肌原纤维骨架强度降低,促使肌肉嫩度升高。长期低pH值和5种与细胞凋亡和蛋白水解有关的内源酶相互作用下的MFI升高是逐渐积累的过程,作用时间滞后,从剪切力数据可以看出,MFI途径对嫩度降低的影响主要集中在4 d后。
