人TRAM基因真核表达载体的构建
2022-06-29高建伟叶峰蒋宏峰
高建伟 叶峰 蒋宏峰
0 引言
TRAM(TRIF-related adaptor molecular)是Toll样受体4(Toll like receptor 4,TLR4)信号通路中的一种接头蛋白,参与了机体固有免疫应答产生干扰素(interferon,IFN)的信号通路[1]。TRAM蛋白含有235个氨基酸,C端含有Toll/IL-1 receptor(TIR)结构域可与TRIF蛋白作用,通过激活下游的转录因子IRF3诱导产生干扰素[2]。干扰素具有抗病毒活性,可通过激活信号级联反应,在病毒进入、转录、RNA稳定性、翻译、成熟和释放等多阶段干扰病毒在机体内增殖[3],在抗病毒方面发挥重要作用。TRAM作为一个TLR4信号通路的调节点,引导信号通路朝向产生干扰素的方向,然而TRAM蛋白是通过什么机制引导TLR4信号的尚不完全清楚。因此,本文拟构建TRAM的真核细胞表达质粒,通过转染入细胞以诱导或增强TLR4信号通路,为进一步研究TRAM的功能及在产生干扰素信号通路中的作用提供研究工具。
1 材料和方法
1.1 材料
人急性单核细胞性白血病细胞(THP-1)购自中国医学科学院基础医学研究所细胞资源中心,HEK-293T细胞购自美国菌种保藏中心(ATCC),Trizol试剂、Lipofectamine 3000转染试剂购自美国Invitrogen公司,M-MLV反转录酶购自Promega公司,KOD-Plus-Ver.2和KOD Dash购自TOYOBO公司,pCMV-N-Flag质粒购自碧云天公司,限制性内切酶SmaⅠ、EcoRⅠ和T4 DNA连接酶购自TAKARA公司,通用型DNA纯化回收试剂盒、凝胶回收试剂盒和DH5α感受态细胞购自康为世纪公司。
1.2 方法
1.2.1 TRAM蛋白编码序列的扩增
用Trizol试剂提取THP-1细胞总RNA,以RT-PCR的方法将RNA反转录为cDNA。再以cDNA为模板,采用PCR的方法扩增TRAM编码区的序列,上游引物:5′ tcccccggggggtatcgggaagtcta 3′,下游引物:5′ ccggaattctcaggcaataaattgt 3′。上下游引物分别引入SmaⅠ和EcoRⅠ酶切位点,采KOD-Plus-Ver.2高保真酶进行PCR。PCR 产物经1%的琼脂糖凝胶电泳,切取含目的序列的凝胶,使用凝胶回收试剂盒对PCR产物进行回收。
1.2.2 pCMV-N-Flag-TRAM重组载体的构建
应用限制性内切酶SmaⅠ和EcoRⅠ,对纯化的PCR产物和pCMV-N-Flag质粒分别进行双酶切,37 ℃,反应2 h。通过T4 DNA连接酶将酶切后的DNA片段和pCMV-N-Flag质粒进行连接反应,16℃过夜。将连接产物转化DH5α感受态细胞,接种到含卡那霉素的LB固体培养基,37℃培养过夜。
1.2.3 重组载体的筛选鉴定
随机挑取LB固体培养基上的克隆进行菌落PCR,将菌落PCR结果为阳性的克隆接种到含卡那霉素的LB液体培养基中,置于37℃摇床内,150 r/min培养过夜,将培养的菌液送公司测序。测序结果与NCBI blast进行比对,与TRAM序列一致的重组载体命名为pCMV-N-Flag-TRAM。
1.2.4 重组载体的表达验证
采用Lipofectamine3000,将重组载体pCMV-N-Flag-TRAM转染HEK-293T细胞。待细胞融合约70%时进行操作,以转染空载体pCMV-N-Flag的HEK-293T细胞作为对照,培养48 h后,提取蛋白进行SDS-PAGE电泳后,通过湿法将蛋白转至硝酸纤维素膜。抗体采用Anti-Flag单克隆抗体(1∶500)和GAPDH单克隆抗体(1∶1000),4 ℃孵育过夜。二抗采用荧光标记的羊抗小鼠IgG(1∶12000),室温孵育2 h。使用Odyssey Sa近红外激光成像系统对膜上的荧光信号进行检测。
2 结果
2.1 TRAM序列的扩增
提取细胞总RNA,经反转录获得cDNA,以cDNA为模板经PCR扩增得到的产物,加样到1%琼脂糖凝胶进行电泳,可见在800 bp附近有一条很亮的条带,且未见杂带,与预期的TRAM编码区目的片段大小位置一致,如图1所示。
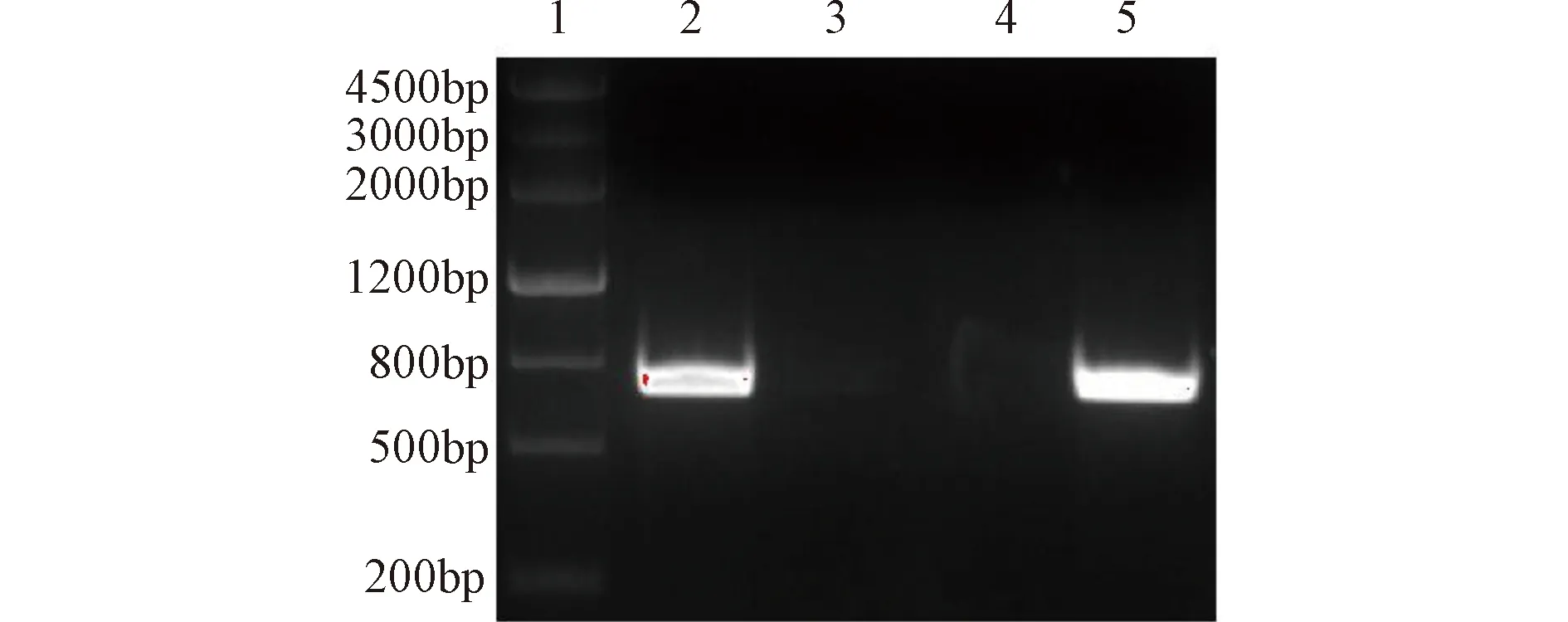
1—DMarker Ⅲ;2,5—PCR阳性产物图1 TRAM编码区PCR产物琼脂糖凝胶电泳结果Figure 1 The agarose gel electrophoresis results ofPCR product from TRAM coding sequence
2.2 重组质粒的筛选鉴定
扩增片段与质粒连接后的产物转化大肠杆菌后,挑取LB固体培养基上的克隆进行菌落PCR,琼脂糖凝胶电泳结果(图2)显示,泳道2和3在800 bp附近出现了目的条带,初步表明泳道2和3对应的菌落含有阳性的重组载体。取阳性菌落培养过夜,提取质粒,经生工公司测序显示如图3,此序列经与NCBI/BLAST RefSeq RNA数据库比对,构建的序列与TRAM序列100%一致。
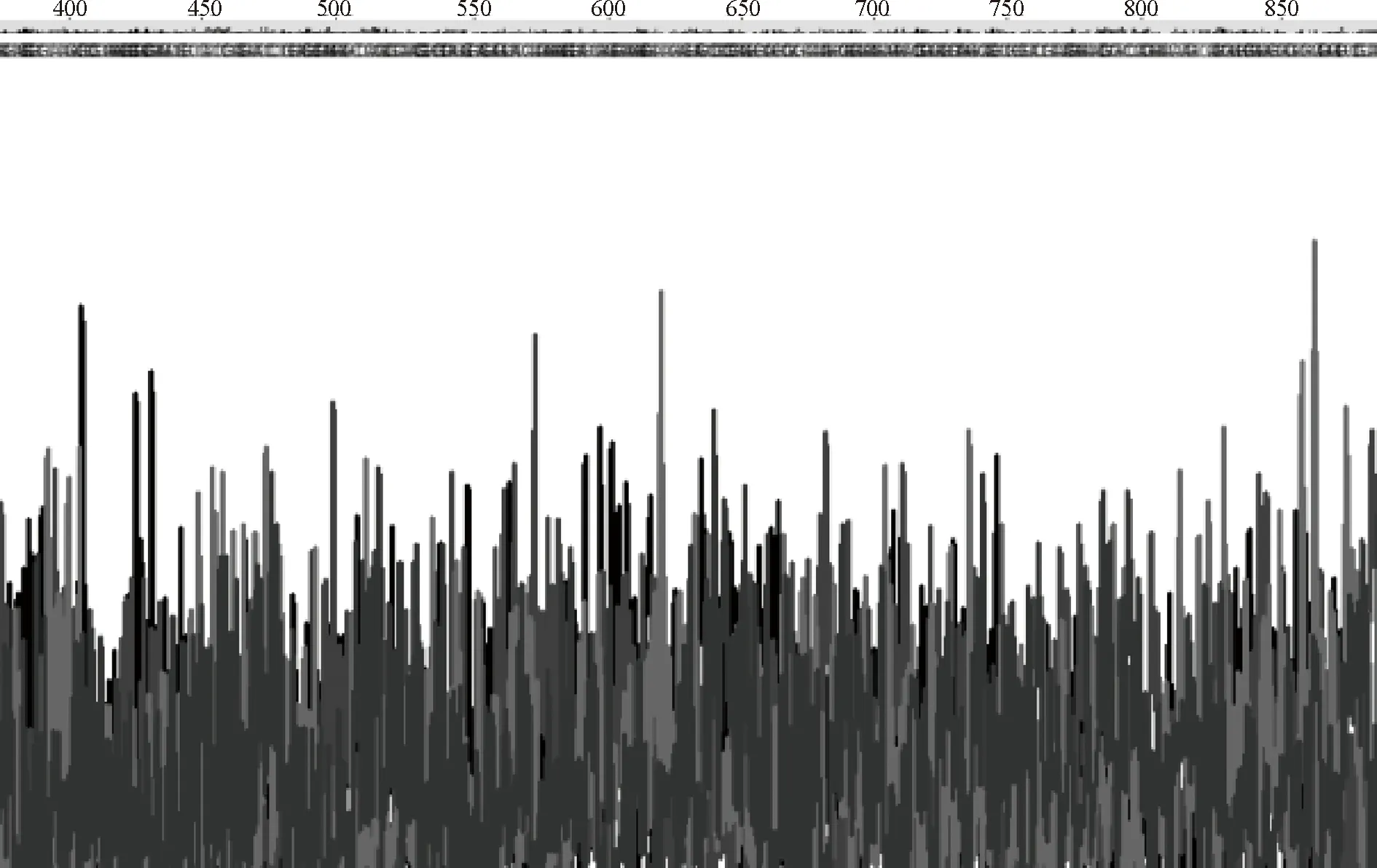
图3 测序片段峰图Figure 3 The sequencing segment peak figure
2.3 Western blot检测Flag-TRAM蛋白在HEK-293T细胞的表达
实验设立3组,分别是:未转染质粒组,作为空白对照;转染pCMV-N-Flag质粒为空质粒组,作为载体对照;转染pCMV-N-Flag-TRAM重组质粒组。Western blot结果显示,转染重组质粒组在约37kD处出现了特异性蛋白条带,空白对照组和载体对照组均未出现目的条带(图4)。
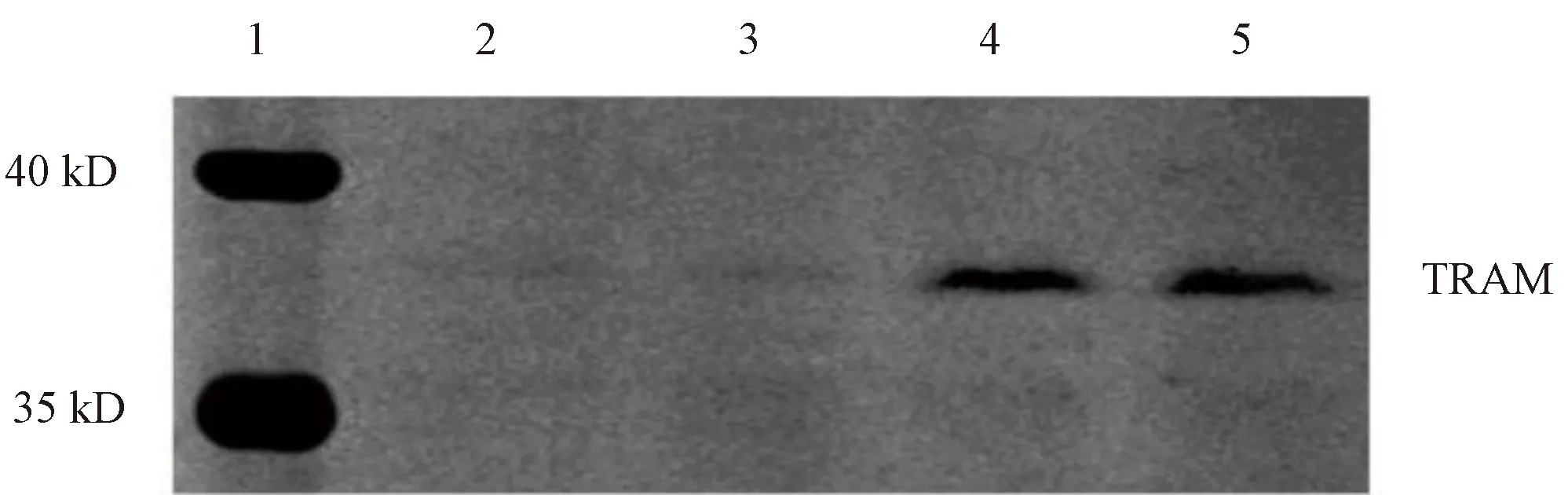
1—预染蛋白Marker;2—未转染组;3—空转染组;4,5—转染重组质粒组图4 Flag-TRAM重组蛋白的表达Figure 4 The expression of Flag-TRAM recombinant protein
3 讨论
干扰素是一种广谱抗病毒效应分子,通过多个靶点有效干扰病毒的复制。机体产生适量的干扰素,在对抗病毒感染性疾病中起着非常重要的作用[4]。病毒感染时释放的RNA和蛋白等病毒组分可激活TLR4介导的固有免疫应答反应,TLR4的信号可通过两组接头蛋白TRAM-TRIF和TIRAP-MyD88形成两条不同的信号通路进行转导,分别激活IRF3转录因子诱导IFN-β、激活NF-κB转录因子诱导炎症因子的产生[5],TLR4对接头蛋白的选择性决定了不同的转录因子激活途径。然而,这两个通路的不平衡可导致多种疾病,如果干扰素产生不足,病毒不能有效清除可导致病毒感染持续加重,而如果炎症因子产生过多,导致出现炎症因子风暴,对机体多脏器产生损伤[6]。因此,深入研究TRAM参与TLR4信号通路的机制,将有助于针对TRAM进行调控,引导TLR4介导的信号通路向产生干扰素的途径,这是抗病毒治疗的一个策略。
本研究采用pCMV-N-Flag质粒作为构建重组载体的骨架,将TRAM mRNA的CDS编码区构建到多克隆位点。实验中将PCR得到的TRAM的编码区通过多克隆位点插入到载体序列,应用菌落PCR的方法,鉴定插入目的片段的阳性克隆。由于PCR过程中可能会出现碱基错配、碱基缺失或碱基增加的情况,为了进一步确认插入片段的碱基序列与目的TRAM序列完全一致,因此应用DNA测序的方法,筛选序列完全一致的克隆。将菌落进行大量培养,提取重组质粒载体,通过Lipofectamine 3000转染的方法,将构建的重组载体pCMV-N-Flag-TRAM转染HEK-293T细胞中,使TRAM蛋白在细胞中高表达,应用Western blot检测到在37kD位置有Flag抗体的条带,可见是Flag的融合蛋白,而未转染质粒组和转染pCMV-N-Flag空质粒组在37kD处均未出现条带。
4 结论
本研究构建了TRAM的真核表达载体,方法简单易操作,重复性好。将TRAM表达载体转染入细胞,可使细胞高表达TRAM蛋白,从而诱导TLR4信号沿着TRIF-TREM依赖的通路产生干扰素。因此,本研究构建的质粒为干扰素的调控和抗病毒免疫治疗提供了一种研究工具。
