金纹细蛾飞行能力测定
2022-06-09侯国辉崔笑雄熊仁次姚永生
侯国辉, 崔笑雄, 朱 悦, 熊仁次, 姚永生
(塔里木大学农学院, 南疆农业有害生物综合治理兵团重点实验室, 新疆阿拉尔 843300)
金纹细蛾Lithocolletisringoniella属鳞翅目(Lepidoptera)细蛾科(Gracillariidae),是果树上一种常见的潜叶性害虫,主要寄主为苹果,其次是海棠、沙果、山定子、山楂、梨、李、桃、樱桃、槟子等果树(石勇强等, 2003; 于江南, 2010; 虞国跃等, 2014)。成虫将卵单粒散产于果树叶片背面,幼虫孵化后潜入叶肉内为害,呈“泡囊”状虫斑,影响叶片的光合作用。发生严重时每片叶上有10~20个虫斑,致使叶片皱缩提前脱落,果实生长受阻或脱落,还可能出现二次开花,影响翌年花芽的生长与结果,造成严重的经济损失(Fanetal., 2015)。2013年在新疆伊犁地区特克斯县首次报道金纹细蛾为害,发生盛期对苹果园为害率达到25%~55%(翟浩等, 2013; 陈川等, 2016; 陈卫民等, 2016)。近年来新疆大力发展特色林果业,苹果在南疆环塔里木盆地种植面积逐年扩大,栽培品种以红富士为主(新疆生产建设兵团统计局, 2013; 姜云斌等, 2019)。红富士品种对金纹细蛾表现为高感,加上气候变暖、栽培管理等诸多因素,为其提供了良好的生存环境,金纹细蛾在苹果园的为害日趋严重(秦玉川和杨书林, 2002),制约了新疆苹果产业的健康可持续发展。
金纹细蛾为东亚特有种,主要分布于日本、韩国、朝鲜及我国北部、中部和西北部地区(Yamadaetal., 1971; Leeetal., 1985; 王春蕾, 2016),国内包括黑龙江、吉林、辽宁、河北、河南、山东、山西、陕西、甘肃、四川、安徽、江苏、新疆等省(自治区),并有逐年扩散的趋势。近年来国内外学者对金纹细蛾形态特征、发生规律、生活习性、生物学特性及防治技术开展了大量研究(石永强等, 2003; 李进, 2012; 张玉琴, 2013; 陈川等, 2016; 刘雪莹等, 2021),但未见有关其飞行能力的相关报道。飞行是昆虫长期进化过程中对生存环境的适应,保持其种群繁衍的一种生存对策,也是昆虫进行短距离扩散、扩大种群危害范围的重要方式,而具有一定飞行能力的昆虫可以在特定的季节借助风力向较远距离的寄主扩散危害,因此昆虫的短距离扩散逐渐被人们重视(Dingle and Drake, 2007; 周国磊等, 2018)。测定昆虫飞行能力是研究昆虫飞行生物学的基础(Camposeetal., 2004),对探讨昆虫飞行扩散机制十分必要,目前普遍采用的方法是利用昆虫飞行磨吊飞(Rileyetal., 1995; 崔建新等, 2016; Keesetal., 2017),准确获取昆虫飞行能力参数。昆虫的飞行能力受多种因素的影响,包括日龄、性别、营养状态、交配状态、生殖、温度、湿度、光照等(袁瑞玲等, 2016; 刘莎等, 2018)。近年来金纹细蛾的为害程度愈加严重,对防治工作带来了诸多困难,为了减少其造成的灾害损失及防治投入的成本,探究金纹细蛾为害范围逐年扩大是否与其飞行能力有关,明确自身生理状态对其飞行能力的影响,对理解金纹细蛾的扩散蔓延、综合防治具有重要指导意义。因此,本研究利用昆虫飞行磨系统,测定了金纹细蛾成虫不同日龄、性别、补充营养与交配状态下的飞行能力参数,旨在了解金纹细蛾的飞行生物学,为阐明其扩散机制和规律、开展预测预报与防控提供理论依据。
1 材料与方法
1.1 供试虫源及饲养
在金纹细蛾越冬代成虫发生高峰期(5月下旬),于阿拉尔市周边及塔里木大学苹果园采集带有金纹细蛾老熟幼虫的虫疤枝叶,带回实验室插入营养液中保湿培养,待老熟幼虫化蛹后将其从虫疤叶中剥离出来,置于培养皿内人工饲养建立实验室种群,每培养皿饲养5头蛹,皿底垫入定性滤纸喷水保湿。饲养过程中,持续到苹果园进行虫疤枝叶的采集,补充室内饲养虫源。待实验室种群羽化后,区别雌雄分别放入相同规格的养虫笼(长×宽×高=75 cm×75 cm×75 cm)中,贴上标签注明羽化日期和性别供试验使用,每日适量添加5%的蜂蜜水为成虫补充营养。金纹细蛾饲养温度为25±1℃、相对湿度为70%±5%、光周期为14L∶10D。
1.2 试验仪器
仪器设备:昆虫飞行磨信息系统(FXMD-24-USB型,河南佳多科工贸股份有限公司)、智能人工气候箱(RXZ-280B,宁波江南仪器厂)、加湿器(Bear JSQ-C50X5)、空调(美的)、温湿度计(HTC-1)、体视显微镜(蔡司V20,北京普瑞赛司仪器有限公司)。
1.3 飞行能力测定
吊飞试验参考郭江龙等(2016)方法并做部分调整,从实验室种群中随机捕获金纹细蛾不同日龄的成虫,保证试虫健康且能正常飞行,置于离心管内并编号。用脱脂棉蘸取少量乙醚将试虫轻微麻醉,然后放于有凹槽的泡沫板上展翅,用毛笔将胸腹连接处的鳞毛除去。取下悬吊在飞行磨上的吊臂,吊臂为直径0.5 mm,长30 cm的细钢丝。蘸取少许502胶将自制吊环与试虫胸腹交界处连接,吊环与试虫虫体保持垂直,静置1 min使胶完全凝固。将吊环套接于飞行磨吊臂末端,待试虫完全苏醒后观察其是否能振翅飞行,若能振翅飞行,将吊臂放回飞行磨上进行测试,保持吊环垂直于吊臂向下,使试虫绕飞行磨中心轴在水平方向上做切线旋转。吊飞试验于每日21∶00时开始,吊飞时间为12 h,次日9∶00时结束。吊飞试验于温度25±1℃,相对湿度70%±5%,完全黑暗的室内进行。
1.3.1不同日龄成虫飞行能力:金纹细蛾羽化后,将雌、雄成虫分别放入相同规格的养虫笼中,饲喂5%蜂蜜水,选取1-10日龄健康的未交配成虫进行测试,连续吊飞12 h,确保每个日龄雌雄成虫分别测试15头。测定未交配成虫最大飞行参数,连续吊飞12 h,每个日龄雌雄成虫分别测试10头,重复3次。
1.3.2不同性别成虫飞行能力:根据1.3.1节测试结果确定未交配成虫的飞行能力最佳日龄,金纹细蛾羽化后,将雌、雄成虫分别放入相同规格的养虫笼中,饲喂5%蜂蜜水,雌、雄成虫各测定有效数量15头(实测20头左右,剔除偏差较大的个体),连续吊飞12 h,记录飞行参数。
1.3.3补充营养的成虫飞行能力:根据1.3.1节测试结果确定未交配成虫的飞行能力最佳日龄,金纹细蛾羽化后,将雌、雄成虫分别放入相同规格的养虫笼中,设置饲喂5%蜂蜜水和清水两个处理,分别测定不同处理雌、雄成虫各15头的飞行参数,连续吊飞12 h。
1.3.4交配与否成虫的飞行能力:根据1.3.1节测试结果确定未交配成虫的飞行能力最佳日龄,金纹细蛾羽化后,设置交配和未交配两个处理。一处理将雌雄成虫放入养虫笼中,任其自由交配;另一处理区分雌雄成虫放入不同养虫笼中,不进行交配。选取未交配雌、雄成虫和已交配雌、雄成虫,饲喂5%蜂蜜水,不同交配状态雌、雄成虫各测定有效数量15头,连续吊飞12 h,记录飞行参数。测试完毕后对雌虫进行卵巢解剖,通过有无精包来确定雌虫是否交配。如有精包则为交配雌虫,与其配对的则为交配雄虫,反之亦然,剔除未交配雌虫的飞行参数。
1.4 数据分析
采用Excel 2010, SigmaPlot和SPSS 20.0软件进行数据处理与分析及图表制作。不同因素对金纹细蛾成虫飞行能力的影响采用单因素方差分析,不同日龄成虫的采用Duncan氏新复极差法比较差异显著性;不同性别、补充营养和交配状态下成虫的飞行能力采用t检验比较差异显著性。数据分析前对平均飞行时间、平均飞行距离、平均飞行速度的原始数据进行正态性检验,如不符合正态分布,则对数据进行平方根转换后再做统计分析,差异显著水平为P<0.05。
2 结果
2.1 不同日龄及不同性别金纹细蛾成虫的飞行能力
金纹细蛾雌雄成虫的飞行能力与日龄之间关系密切,随着日龄增加呈先升高后降低的变化趋势(表1)。连续12 h成虫吊飞结果表明:不同日龄雌雄成虫飞行能力存在明显差异。不同日龄雌成虫的飞行距离(F=4.624,df1=9,df2=140,P<0.05)、飞行时间(F=7.731,df1=9,df2=140,P<0.05)、飞行速度(F=2.635,df1=9,df2=140,P<0.05)存在显著差异;不同日龄雄成虫的飞行距离(F=5.145,df1=9,df2=140,P<0.05)、飞行时间(F=8.465,df1=9,df2=140,P<0.05)、飞行速度(F=3.242,df1=9,df2=140,P<0.05)也存在显著差异。3-6日龄成虫飞行能力相对较强,此后随着日龄增加飞行能力逐渐减弱,到10日龄成虫飞行能力最弱,雌雄成虫的飞行能力均在5日龄达到峰值,雌成虫平均飞行距离2.293±0.254 km,平均飞行时间5.341±0.617 h,平均飞行速度0.711±0.126 km/h;雄成虫平均飞行距离2.142±0.276 km,平均飞行时间5.132±0.628 h,平均飞行速度0.620±0.132 km/h。虽然金纹细蛾雌成虫的平均飞行距离、飞行时间和飞行速度均大于雄成虫,但雌雄个体间平均飞行距离(t=0.40,df=28,P=0.69)、飞行时间(t=0.24,df=28,P=0.81)和飞行速度(t=0.50,df=28,P=0.62)均不存在显著差异(P>0.05)。
金纹细蛾雌雄成虫的最长飞行距离、最长飞行时间和最大飞行速度均在5日龄达到峰值(表2)。最长飞行距离分别为4.272±0.201和4.223±0.219 km,最长飞行时间分别为8.733±0.243和8.671±0.179 h,最大飞行速度分别为1.725±0.199和1.632±0.164 km/h。
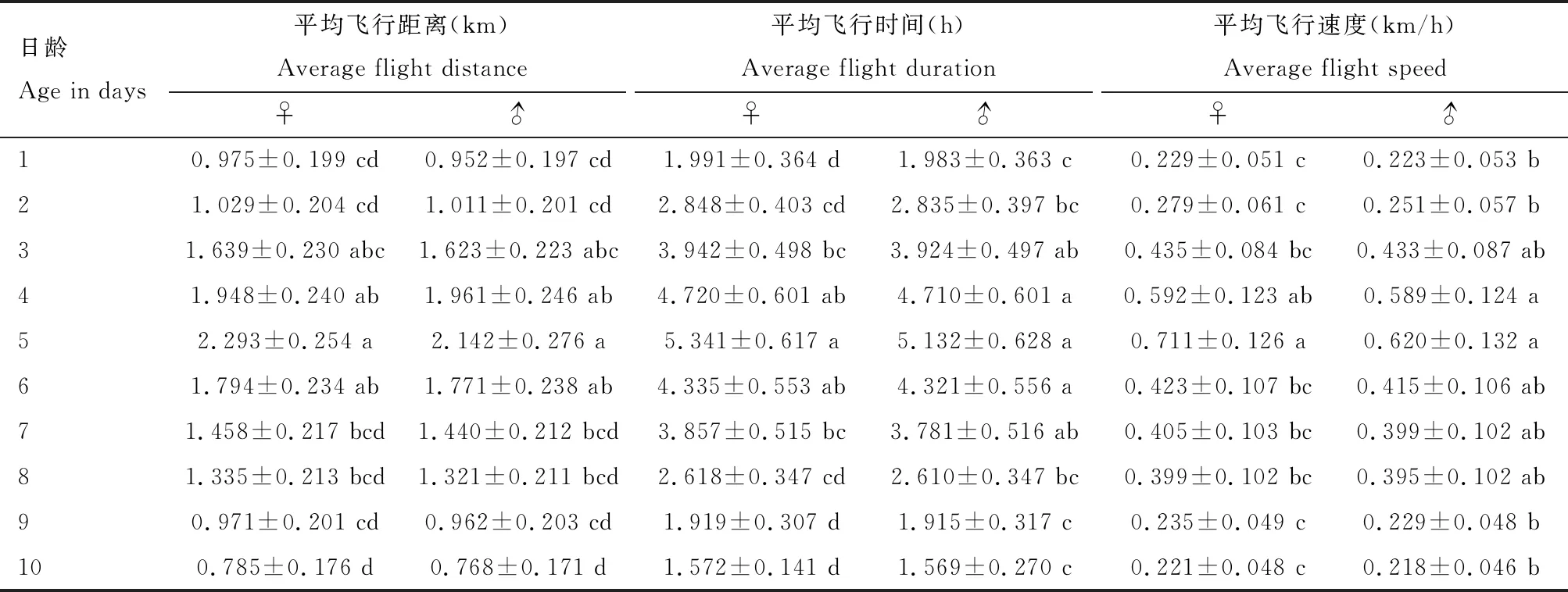
表1 金纹细蛾不同日龄成虫平均飞行距离、飞行时间和飞行速度Table 1 Average flight distance, flight duration and flight speed of different day-old adults of Lithocolletis ringoniella
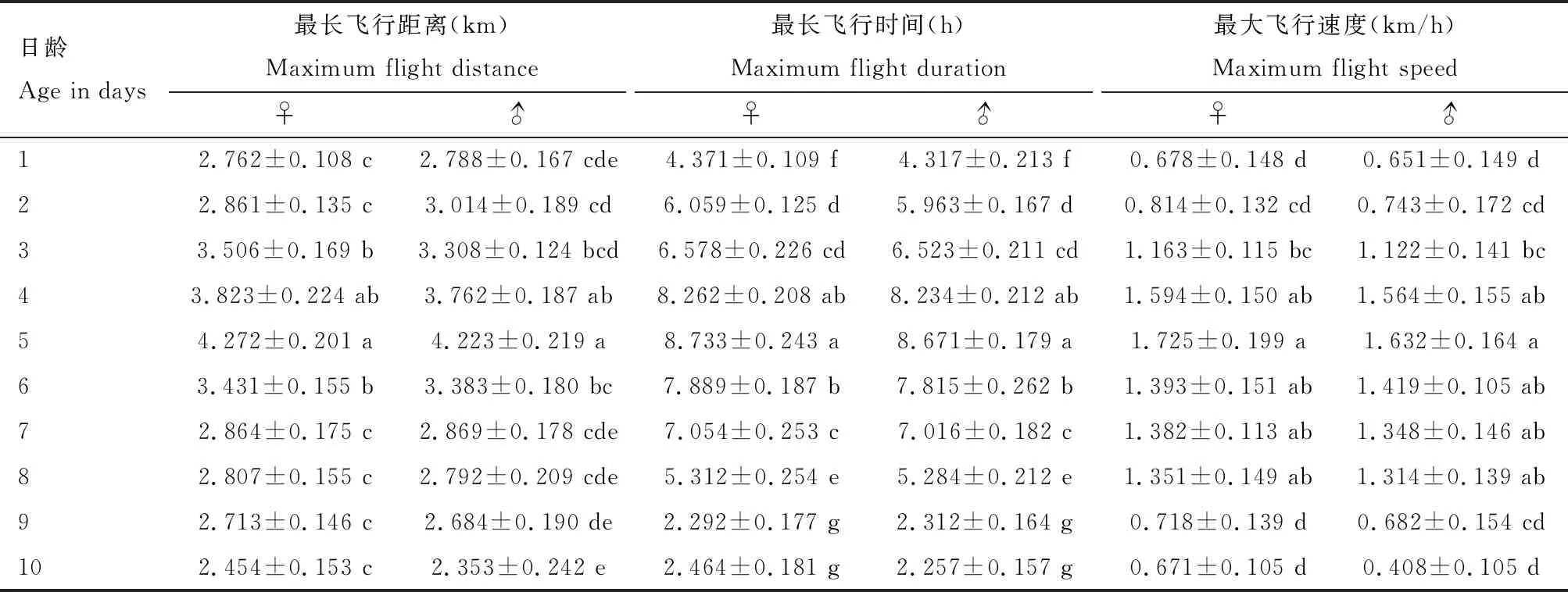
表2 金纹细蛾不同日龄成虫最长飞行距离和飞行时间以及最大飞行速度Table 2 Maximum flight distance, flight duration and flight speed of different day-old adults of Lithocolletis ringoniella
2.2 补充营养对金纹细蛾成虫飞行能力的影响
金纹细蛾未交配雌雄成虫的飞行能力均在5日龄达到峰值,分别测定不同营养状态的未交配5日龄雌雄成虫连续吊飞12 h的飞行能力,结果表明:取食5%蜂蜜水金纹细蛾雌雄成虫的飞行距离、飞行时间、飞行速度均大于对照(取食清水),取食5%蜂蜜水雌成虫的飞行距离(t=2.62,df=28,P<0.05)和飞行速度(t=0.71,df=28,P<0.05)与对照存在显著差异,飞行时间(t=1.07,df=28,P>0.05)与对照比无显著差异;取食5%蜂蜜水雄成虫的飞行距离显著高于对照(t=2.09,df=28,P<0.05),飞行时间(t=0.87,df=28,P>0.05)和飞行速度(t=0.24,df=28,P>0.05)与对照比差异不显著。金纹细蛾成虫在不同营养状态下飞行能力存在差异,雌雄成虫取食补充营养后飞行能力均有提高(图1)。取食后雌成虫的飞行距离(2.293±0.254 km)、飞行时间(5.341±0.617 h)和飞行速度(0.711±0.126 km/h)较未取食雌成虫的飞行距离(1.561±0.117 km)、飞行时间(4.627±0.252 h)和飞行速度(0.613±0.057 km/h)分别提高46.945%, 15.430%和15.978%;取食后雄成虫的飞行距离(2.142±0.276 km)、飞行时间(5.132±0.628 h)和飞行速度(0.620±0.132 km/h)较未取食雄成虫的飞行距离(1.502±0.132 km)、飞行时间(4.518±0.317 h)和飞行速度(0.582±0.084 km/h)分别提高42.610%, 13.590%和6.529%。
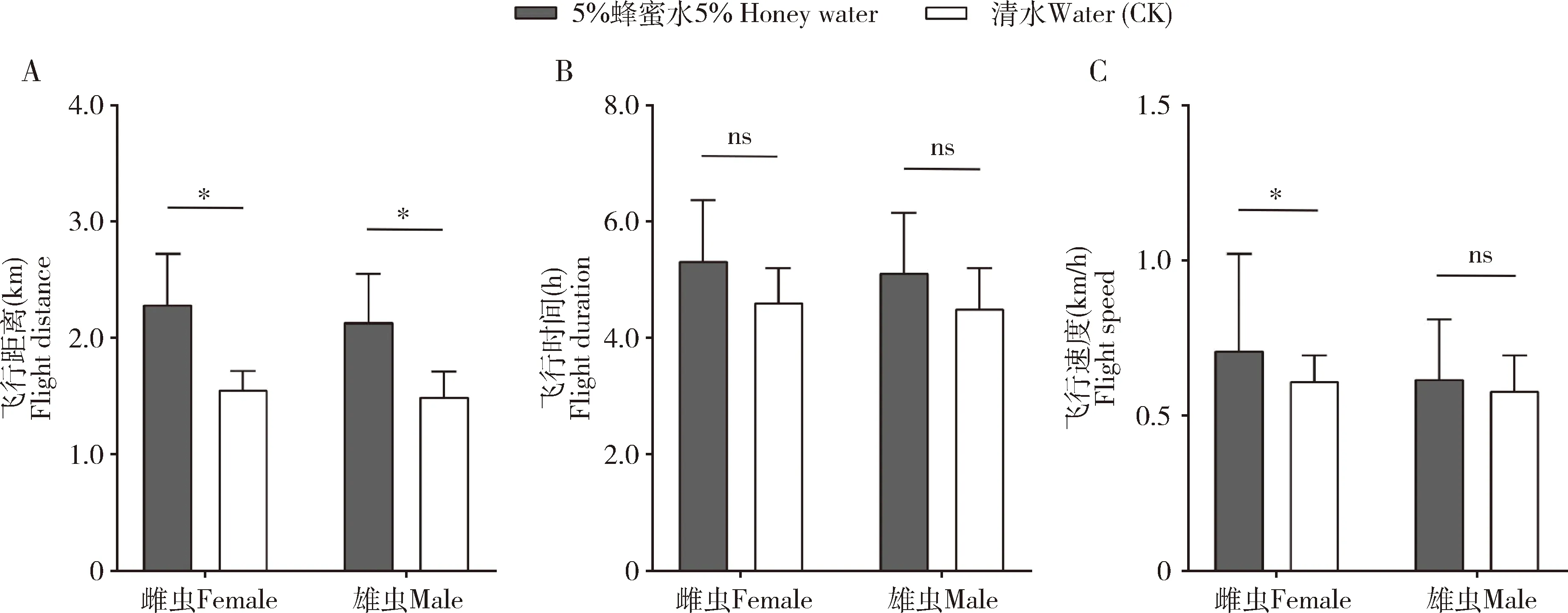
图1 补充营养时金纹细蛾5日龄雌雄成虫间飞行能力的比较Fig. 1 Comparison of flight capacity between the 5-d-old female and male adults ofLithocolletis ringoniella provided with complementary nutritionA: 飞行距离Flight distance; B: 飞行时间Flight duration; C: 飞行速度Flight speed. 图中数值为平均值±标准误;柱上星号和ns分别表示差异显著(P<0.05)和不显著(P>0.05)(t检验)。图2同。Data in the figure are mean±SE. Asterisk and ns above bars indicate significant difference (P<0.05) and no significant difference (P>0.05)(t-test), respectively. The same for Fig. 2.
2.3 交配状态对金纹细蛾成虫飞行能力的影响
金纹细蛾未交配雌雄成虫的飞行能力均在5日龄时达到峰值,分别测定不同交配状态的5日龄雌雄成虫连续吊飞12 h的飞行能力,结果表明:金纹细蛾雌成虫交配后飞行能力提高,雄成虫交配后飞行能力降低(图2)。金纹细蛾雌成虫交配后飞行距离(t=3.02,df=28,P<0.05)和飞行速度(t=2.14,df=28,P<0.05)显著大于未交配雌成虫,雄成虫交配后飞行距离(t=2.58,df=28,P<0.05)和飞行速度(t=2.14,df=28,P<0.05)显著小于未交配雄成虫;交配与未交配雌雄成虫飞行时间无显著差异(t=0.54,df=28,P>0.05;t=1.36,df=28,P>0.05),雌成虫交配后飞行时间延长,雄成虫交配后飞行时间缩短。交配后雌成虫的飞行距离(3.248±0.188 km)、飞行时间(5.723±0.340 h)和飞行速度(0.995±0.041 km/h)较未交配雌成虫的飞行距离(2.293±0.254 km)、飞行时间(5.341±0.617 h)和飞行速度(0.711±0.126 km/h)分别提高41.628%, 7.152%和39.925%;而交配后雄成虫的飞行距离(1.375±0.111 km)、飞行时间(4.214±0.254 h)和飞行速度(0.334±0.022 km/h)较未交配雄成虫的飞行距离(2.142±0.276 km)、飞行时间(5.132±0.328 h)和飞行速度(0.620±0.132 km/h)分别降低35.823%, 17.888%和46.129%。
3 讨论
本研究通过对金纹细蛾不同日龄成虫飞行能力的测定表明(表1):金纹细蛾成虫的飞行能力与日龄之间关系紧密,随着日龄增长表现为由弱到强,再由强到弱的变化趋势。羽化初期,飞行能力随日龄的增加逐渐增强,5日龄达到峰值,之后随日龄的增加逐渐下降,10日龄飞行能力最弱。这种飞行能力变化的规律与苹果蠹蛾Cydiapomonella、草地螟Loxostegestieticatis、草地贪夜蛾Spodopterafrugiperda、甘蓝夜蛾Mamestrabrassicae、二点委夜蛾Athetislepigone和伞裙追寄蝇Exoristacivilis等(罗礼智和李光博, 1992; Schumacheretal., 1997; 郑作涛等, 2014; 吴晓等, 2016; 岳方正等, 2016; 葛世帅等, 2019)相似。羽化初期飞行能力相对较弱,可能是由于营养的供应和吸收不够完全,其飞行肌的发育尚未完成,飞行能源物质和有关酶活性较低,各项飞行参数比较低。随着飞行肌等发育成熟,飞行能源物质的积累和酶活性的增强,飞行能力逐渐提高,当达到一定的发育阶段,昆虫的重心转向生殖,卵巢开始逐渐发育并且导致飞行肌的降解,从而飞行能力下降(Johnson, 1969; Sunetal., 2013)。本试验测定结果表明金纹细蛾最佳日龄成虫可连续飞行5 h左右,最长飞行距离4 km以上(表2),虽不及已报道的甜菜夜蛾Spodopteraexigua、草地贪夜蛾和二点委夜蛾等(郑作涛等, 2014; 邹金城等, 2017; 葛世帅, 2019),但仍具有迁飞扩散的潜力,这可能是导致近年来其为害范围逐年扩大的重要原因之一。金纹细蛾雌雄成虫的飞行能力几乎相同,虽然雌成虫的平均飞行距离、飞行时间和飞行速度均大于雄成虫,但雌雄个体间平均飞行距离、飞行时间和飞行速度均不存在显著差异(表1)。这一研究结果与苹果蠹蛾、黄地老虎Agrotissegetum、橘小实蝇Bactroceradorsalis和草地贪夜蛾等(Schumacheretal., 1997; 崔建新等, 2016; 郭江龙等, 2016; 葛世帅等, 2019)相似,与绿盲蝽Lyguslucorum、花曲柳窄吉丁Agrilusmarcopoli和家蝇Muscadomestica等(Luetal., 2007; 刘吉起等, 2011; 王辉等, 2015)相悖。金纹细蛾雌成虫的飞行能力基本参数均大于雄成虫,可能是由于雌虫为了寻求交配及完成交配后寻找适宜的产卵场所,对飞行的需求比雄虫更为强烈。一般而言,体型较大的昆虫飞行能力较强,雌性昆虫飞行能力强于雄性,这与雌雄生理或形态差异有关(Shirai, 1995; Luetal., 2007; 王辉等, 2015)。
成虫期取食对昆虫飞行能力具有重要影响,成虫在饥饿状态下或长期营养不足时飞行能力显著下降(张蕾等, 2006; 郭文超等, 2012)。本研究结果表明获取补充营养(5%蜂蜜水)的金纹细蛾雌雄成虫平均飞行距离均有显著提高(图1),营养物质的及时补充是金纹细蛾飞行扩散的关键。这一研究结果与黄地老虎、二点委夜蛾和草地贪夜蛾等(郑作涛, 2014; 郭江龙等, 2016; 葛世帅, 2019)一致。交配对生殖和飞行能力的影响在多种昆虫中已被证实。本研究结果表明金纹细蛾雌成虫交配后飞行能力提高,雄成虫交配后飞行能力降低(图2)。交配对雌成虫飞行能力有较大的影响,交配后雌成虫飞行能力更强,有利于其寻找合适的寄主植物来产卵(Hanskietal., 2006; Hashiyamaetal., 2013)。暗脉菜粉蝶Pierisnapi的飞行能力研究表明,未交配雄成虫的飞行速度大于已交配,交配后雄成虫飞行能力减弱(Almbro and Kullberg, 2012)。这一结论在本实验中也得到印证,交配过程中雄虫向雌虫体内转移了许多生理效应的物质,包括肽类物质、保幼激素类似物、前列腺素等多种“交配因子”(华荣胜等, 2010),促使雌虫将用于飞行的能源物质转移到生殖方面,飞行能力随之下降。
本研究在室内对金纹细蛾成虫飞行能力进行了初步探索,明确了不同日龄、性别、营养状态、交配状态对成虫飞行能力的影响。此外,影响金纹细蛾飞行能力的因素还包括振翅频率、生殖、种群密度、温湿度和光照强度等(程登发等, 2002; 江幸福和罗礼智, 2008),影响金纹细蛾飞行能力的因素仍有待于进一步探索。本试验供试虫源为实验室种群,利用飞行磨系统测定金纹细蛾飞行生物学参数,不能完全模拟该虫在自然条件下的飞行状态,所得结果与实际存在一定差异。因此,采取室外室内相结合的方法,进一步研究影响金纹细蛾飞行能力的因素十分必要。
