金斑蝶觅食求偶过程中的视觉嗅觉利用
2022-06-09李明涛陈顺安陈晓鸣
刘 杰, 李明涛, 陈顺安, 姚 俊, 石 雷, 陈晓鸣
(中国林业科学研究院高原林业研究所, 国家林业局资源昆虫培育与利用重点实验室, 昆明 650224)
蝴蝶是昆虫纲(Insecta)鳞翅目(Lepidoptera)锤角亚目(Rhopalocera)昆虫的统称。蝴蝶成虫的任务是繁衍后代,觅食和求偶是成虫的主要活动。蝴蝶觅食和求偶是通过视觉和嗅觉来完成的,不同蝶种在觅食和求偶过程中,视觉和嗅觉利用具有不同的权重(Tangetal., 2013; 李成哲, 2017)。
在觅食过程中,花的颜色和气味等是最显著的蜜源特征(Andersson, 2003;mura and Honda, 2005; Balkeniusetal., 2006)。蝴蝶的视网膜较为复杂,可依靠色觉来识别花朵、寻找配偶以及确定产卵地点(Perryetal., 2016)。花的颜色、形状和大小等远距离视觉信号在蝴蝶寻找蜜源的过程中起着重要的作用,而蜜源植物的花香作为一种近距离嗅觉信号也可以吸引蝴蝶访花(王翻艳, 2015; 李明涛等, 2020)。如白带锯蛱蝶Cethosiacyane以视觉为主定位寻找食物,对花色趋向多偏向长波长的红、橙和黄色花,对短波长的紫色和混合光白色花访问较少(Tangetal., 2013)。孔雀蛱蝶Inachisio、荨麻蛱蝶Aglaisurticae和钩粉蝶Gonepteryxrhamni访花时以植物的花香作为嗅觉线索识别花蜜(Andersson, 2003)。虎斑蝶Danausgenutia在访花时仅依靠嗅觉(唐宇翀, 2013)。青斑蝶Tirumalalimniace、大帛斑蝶Idealeuconoe和红锯蛱蝶Cethosiabiblis在访花过程中充分利用嗅觉(唐宇翀, 2013; 王翻艳, 2015; 王华, 2017)。大红蛱蝶Vanessaindica访花过程中主要依靠颜色,其次依靠气味(mura and Honda, 2005)。
蝴蝶在求偶过程中面临的首要问题是雌雄蝶如何区分同性和异性,以迅速找到异性交配从而避免时间的浪费。一般认为视觉信号在远距离识别中发挥主要作用,而嗅觉信号在近距离识别中发挥重要作用(Silberglied and Taylor, 1978; Grula and Taylor, 1980; Costanzo and Monteiro, 2007)。求偶过程中蝴蝶对视觉信号的利用主要包括翅面特征(颜色、眼斑和大小等)、翅面反射光和偏极光等,如性二色性的蝴蝶多通过自然光下的翅面色彩来识别异性(Stavenga and Arikawa, 2006; Lietal., 2017; 李明涛, 2020),菜粉蝶Pierisrapae雄蝶可以通过翅面的视觉线索来识别同性和异性(Obara and Majerus, 2000; Obaraetal., 2008),青衫黄袖蝶Heliconiuscydno在异性识别中雄蝶通过偏极光识别雌蝶(Sweeneyetal., 2003)。Huq等(2019)发现偏瞳蔽眼蝶Bicyclusanynana在求偶中雄蝶通过雌蝶前翅腹面眼点识别雌蝶。近距离求偶时,雄蝶通过散发的气味来传递化学信号,这种化学信号,被认为是对蝴蝶视觉模式的补充(Vane-Wright and Boppré, 1993),对于视觉不发达的蝴蝶(如枯叶蛱蝶Kallimainachus),必须利用其他信号对同类或配偶做进一步辨别,嗅觉信号往往是这类蝴蝶的主要识别依据(唐宇翀, 2013)。黑纹粉蝶Pierismelete雌蝶依靠挥发物(雄蝶翅面释放的物种特异性气味)来识别同类配偶(Kan and Hidaka, 1997)。mura等(2001)研究发现金凤蝶Papiliomachaon雌雄蝶释放的共有挥发物在异性识别中起到重要作用。
金斑蝶Danauschrysippus隶属于鳞翅目(Lepidoptera)斑蝶科(Danaidae)斑蝶属Danaus(周尧, 1994)。目前对金斑蝶的研究主要集中在性信息素(Boppréetal., 1978)、生物学特性(王文明, 2011)、访花(唐宇翀, 2015)、学习和求偶行为(王华, 2017)等方面。有关金斑蝶觅食求偶过程中视觉嗅觉利用系统研究尚未见有报道。本研究通过金斑蝶觅食和求偶过程中的视觉和嗅觉选择行为观察,掌握金斑蝶觅食求偶时视觉嗅觉信息利用机制,为开展蝴蝶的规模化人工繁育提供理论和实用依据;另外蝴蝶对视觉嗅觉两个功能的偏爱可能反映了其进化的线索,本研究可为进一步探索蝴蝶的进化提供基础资料。
1 材料与方法
1.1 试验地点
试验于中国林业科学研究院高原林业研究所的禄丰试验站进行,试验站海拔1 382 m,属中亚热带季风气候。年均气温16.2℃,年平均降雨量930~950 mm,年无霜期322 d。行为学观察试验在网室(8 m×5 m×5 m)内进行。网室内透光良好,光照均匀。
1.2 试验材料
1.2.1供试蝴蝶:金斑蝶来自云南省元江县人工饲养种群,幼虫用牛角瓜Calotropisgigantean饲喂,饲养条件为温度26±2℃,相对湿度50%~70%,光周期13L∶11D。供试蝴蝶均为羽化后未交配的健康成虫。试验前1 d将待试蝴蝶释放于网室内,以适应网室环境。试验前,仅提供清水供所有蝴蝶任意吸食。
1.2.2供试假花:选择红、橙、黄、蓝、紫、绿和白色花冠的人造花(图1: A)。7种颜色假花均由棉布制成,且形状基本相同。用光谱仪(USB200 +, Ocean Optics, Inc., 美国)测量花色的反射光谱,该光谱仪针对MgO涂覆的表面作为参考进行校准,在380~780 nm波长范围内各色假花的光谱反射率如图1(B)。
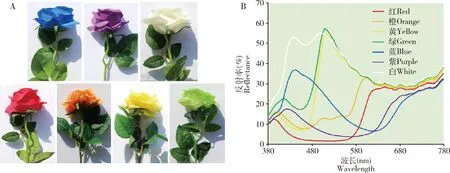
图1 7种颜色的假花和可见光范围内光谱反射率Fig. 1 Artificial flowers with seven colors and their spectral reflectance in visible lightA: 7种颜色的假花Artificial flowers with seven colors; B: 7种颜色假花可见光范围内光谱反射率Spectral reflectance of the seven colors in visible light.
1.2.3供试蜂蜜水:以枣花蜂蜜为原料,用纯净水稀释配制为10%浓度的蜂蜜水(唐宇翀, 2013)。
1.2.4塑封模型:塑封翅模型制作:将蝴蝶真翅按照背腹面粘在黑色木条作的虫体上,再粘上黑色假触角,用PVC塑料塑封(图2)。
1.2.5金斑蝶色彩及光谱分析:在自然光下,金斑蝶雌雄翅面颜色和斑纹非常相近,翅面由棕、黑、桔黄、黄和白等颜色混合而成。以棕色为主,除了雄蝶后翅腹面比雌蝶后翅腹面多一白色性标斑点,雌雄蝶翅色斑无明显区别(图3: A)。蝶翅塑封后,自然蝶翅与蝶翅模型在视觉上差异很小(图3: A-E),通过光谱分析可知,在自然蝶翅与塑封蝶翅模型的光谱反射波特征基本一致,虽然塑封模型的光谱反射率略低于自然种群,但在重要的反射率峰阶段差异细微,塑封后的蝶翅对金斑蝶寻找配偶干扰较小。在紫外光(UV)下,金斑蝶雌雄蝶翅与蝶翅模型的区别明显,蝶翅模型图像较自然蝶翅更清晰,更易识别(图3: D-E)。

图2 金斑蝶成虫塑封模型Fig. 2 Artificial models of Danaus chrysippus adultsA: ♀, 背面Dorsal surface; B: ♀, 腹面Ventral surface; C: ♂, 背面Dorsal surface; D: ♂, 腹面Ventral surface.
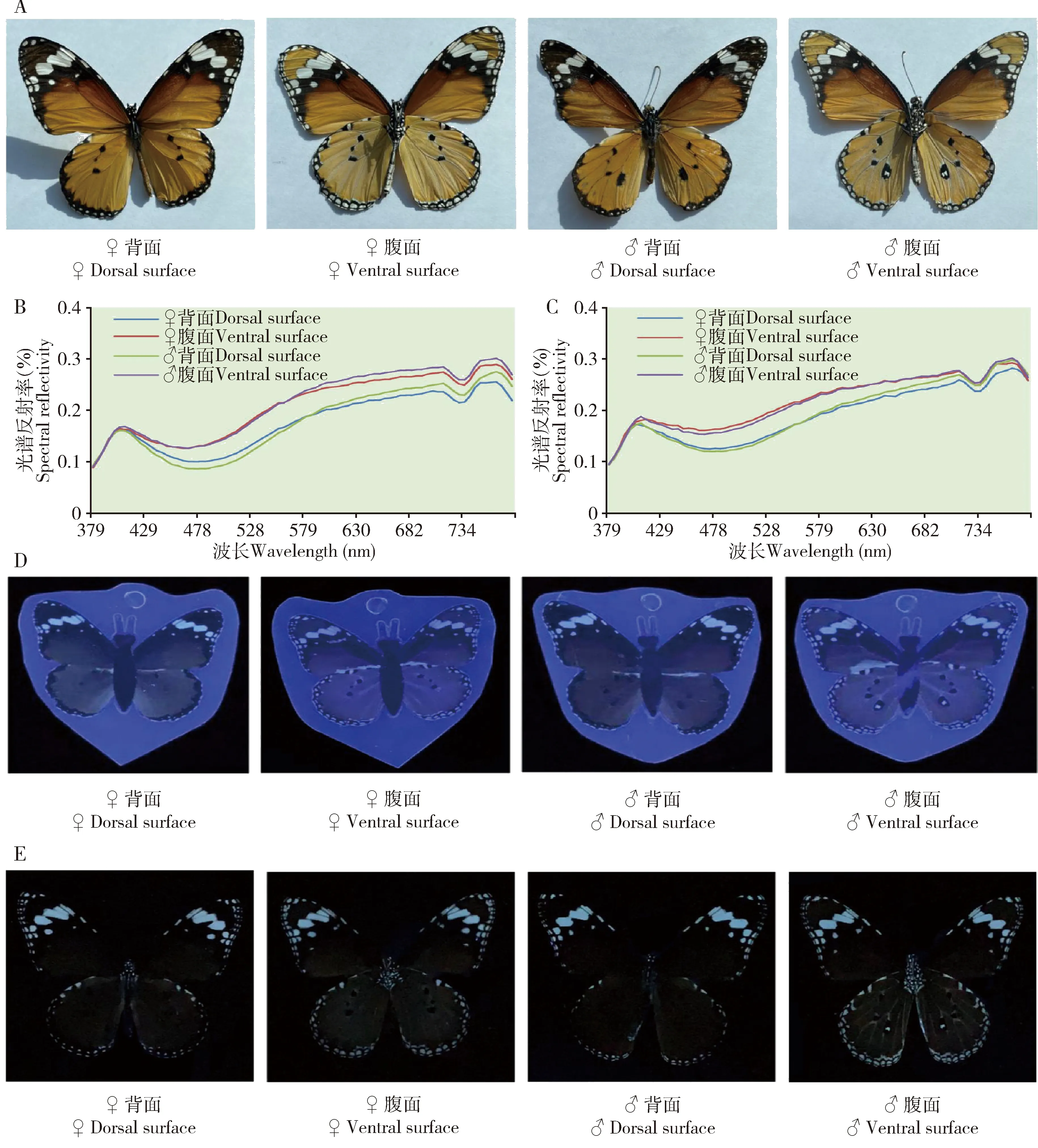
图3 金斑蝶雌雄蝶自然种群及塑封模型色彩Fig. 3 Color of natural population and artificial model of female and male adults of Danaus chrysippusA: 自然光下自然种群色彩Color of natural population under natural light; B: 自然种群光谱分析 Spectral analysis of natural population; C: 塑封模型光谱分析Spectral analysis of artificial model; D: 紫外光下金斑蝶成虫自然种群色彩Color of natural population of D. chrysippus adults under ultraviolet(UV); E: 紫外光下金斑蝶成虫塑封模型色彩Color of artificial model of D. chrysippus adults under UV.
1.3 金斑蝶成虫觅食过程中对颜色的行为响应观察
将红、橙、黄、蓝、紫、绿和白色7种颜色的假花分别按相同颜色各自集中在一起,花朵距地面约0.5~0.7 m,相邻花束间距为80 cm。在天气晴朗、光照充足,温度18~30℃,相对湿度60%~80%的环境条件下,放入金斑蝶雌、雄蝶各10头,观察时间为9∶30-11∶30和14∶30-17∶30时,7种颜色的假花30 min位置按顺时针方向移动1次,避免位置带来的误差。重复试验3次(下同)。
取食:金斑蝶停落在假花上,探食后伸出喙管并开始吸食假花上的蜂蜜水,记录为取食。
访花次数:从金斑蝶着落在花朵上伸出喙管探食或取食,至金斑蝶卷曲喙管飞离花朵记为1次访花。若同一头蝴蝶飞离后,在不超过1 m距离内又折回到同一花朵上,则仅记为1次访花(下同)。
1.4 金斑蝶成虫觅食过程中的视觉和嗅觉行为响应观察
在红、橙、黄、绿、蓝、紫和白色7种假花花束上各喷洒10%的蜂蜜水10 mL,为避免蒸发带来的影响,每隔1 h在各花束上补喷10%蜂蜜水10 mL。观察并记录金斑蝶对不同颜色花朵的访问次数。
为了明确气味对金斑蝶的诱集效果,试验将花冠去掉,剩下花枝和叶片,将10%蜂蜜水喷洒在无花冠的花枝上,观察并记录金斑蝶对无花冠的花枝的访问次数。
1.5 金斑蝶求偶过程中的视觉和嗅觉行为响应观察
在网室中放入20头(雌∶雄=1∶1)处于求偶期的金斑蝶,观察雌雄蝶之间的追逐情况,为区别雌雄,雌蝶后翅用红色记号笔标记,观察记录雌雄蝶的追逐行为。
在求偶期,在网室内垂直悬挂雌、雄蝶塑封模型各4只,模型距地垂直高度约1.8 m,间距1 m左右。分别放入雌雄金斑蝶各10头,每2 h交换一次模型位置,观察并记录雌雄蝶追逐模型行为的次数。
1.6 金斑蝶色彩及光谱分析
将雌雄金斑蝶标本及模型按背腹面顺序平展开,用光谱仪(USB200+, Ocean Optics, Inc, 美国)测量自然光波长范围内的反射光谱,利用Excel绘制色谱图。用紫外仪(北京六一紫外仪, 型号WD-9403C, 反射波长365 nm)拍摄标本和蝴蝶模型紫外图像。
1.7 数据分析
使用SPSS 25.0统计分析软件,对金斑蝶访问不同颜色假花和自然种群求偶的数据分别采用单因素方差分析,当方差齐性时用邓肯法进行多重比较;对金斑蝶访问喷洒蜂蜜水前后的数据分别采用独立样本t检验。
2 结果
2.1 金斑蝶成虫觅食过程中对颜色的行为响应
观察结果表明,雌雄蝶对7种颜色的假花均有访问,共访问138次。雌雄蝶均对红色假花访问次数最多,为72次(占总访花次数的52.17%);其次是对蓝色(21次,占总访花次数的15.22%)和橙色(16次,占总访花次数的11.59%),对其他4种颜色假花的访问相对较少(图4: A),说明不同颜色对金斑蝶觅食有不同的引导作用,金斑蝶对长波长红色趋性最强。雌雄蝶访花次数有差异,雌雄蝶对7种颜色假花访问次数占总访问次数的比例分别为42.02%和57.97%,其中,雌雄蝶对红色假花的访问比例分别为21.74%和30.43%,雌雄蝶对蓝色假花的访问比例分别为5.80%和9.42%,雌雄蝶对橙色假花的访问比例分别为5.07%和6.52%。雄蝶对红色的响应显著高于雌蝶(P<0.05)(图4: A)。
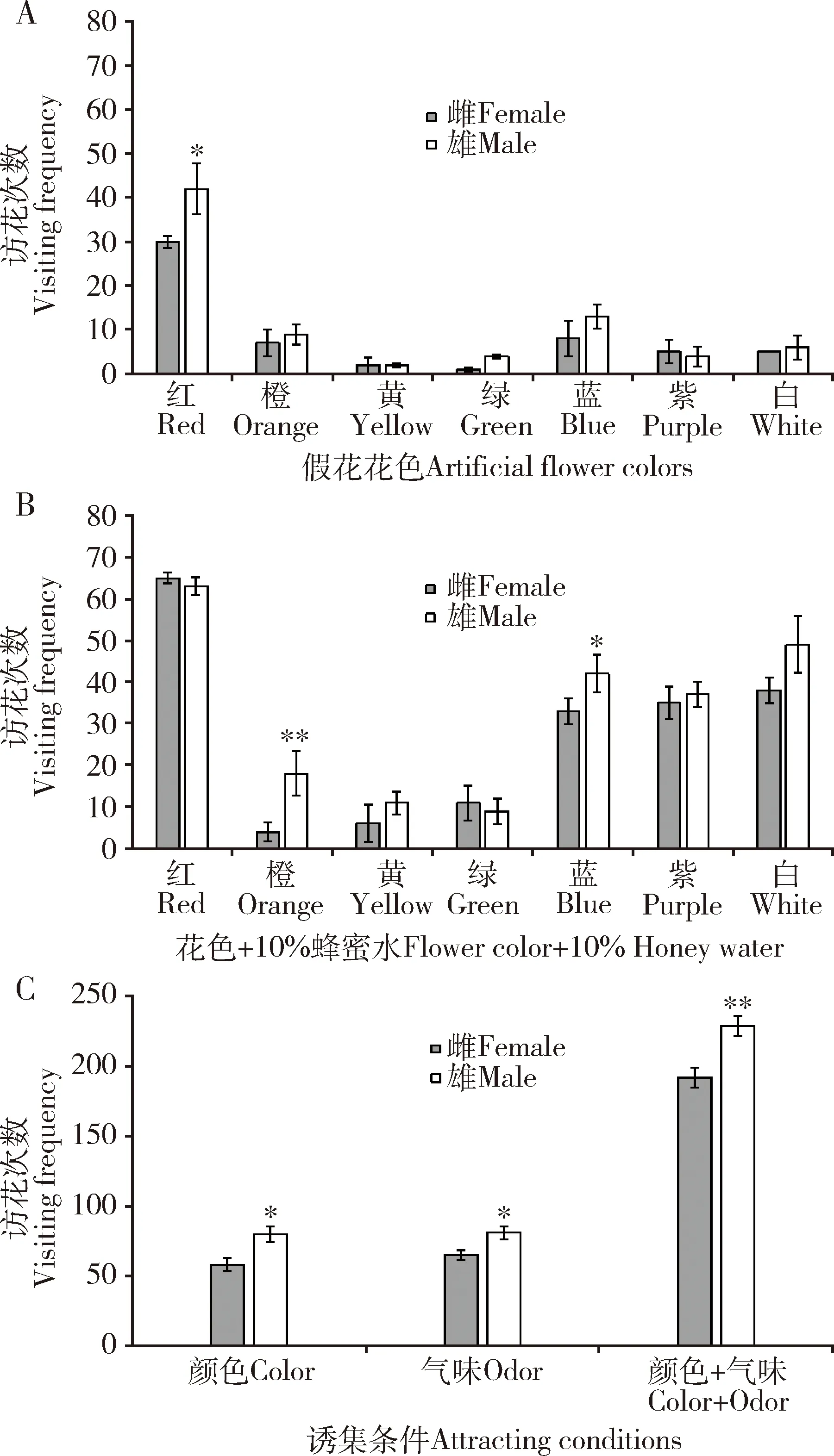
图4 金斑蝶成虫对颜色及颜色+气味的行为反应Fig. 4 Behavioral responses of Danaus chrysippus adults to colors and colors plus odorA: 金斑蝶成虫访问7种颜色假花次数Visiting frequencies of D. chrysippus adults to artificial flowers of seven colors; B: 金斑蝶成虫访问7种颜色假花+10%蜂蜜水次数Visiting frequencies of D. chrysippus adults to artificial flowers of seven colors+10% honey water; C: 金斑蝶成虫对颜色、气味及颜色+气味的识别反应比较Comparison of the recognition responses of D. chrysippus adults to color, odor, and color+odor; 颜色表示单一视觉诱集时金斑蝶的访花次数,气味表示单一嗅觉诱集时金斑蝶的访问次数,颜色+气味表示视觉嗅觉综合利用时金斑蝶的访花次数Color indicates the visiting frequencies of D. chrysippus to a single visual luring, odor indicates the visiting frequencies of D. chrysippus to a single olfactory luring, and color+odor indicates the visiting frequencies of D. chrysippus in the combination utilization of visual and olfactory. 图中数据为平均值±标准差;柱上星号和双星号分别表示雌雄蝶之间差异显著(P<0.05)和极显著(P<0.01)(t检验)。Data in the figure are mean±SD. The asterisk and double asterisk above bars indicate significant difference (P<0.05) and extremely significant difference (P<0.01), respectively, between the female and male butterflies by t-test.
2.2 金斑蝶成虫觅食过程中的视觉和嗅觉行为响应
在无气味假花上喷洒10%蜂蜜水后,雌雄蝶总访花次数为421次,与单一颜色诱集相比增加了3.05倍。雌雄蝶对7种颜色假花访问次数分别为192次和229次,其所占总访问次数的比例分别为45.60%和54.39%。雄蝶总访花次数极显著高于雌蝶的(P<0.01),蝴蝶对颜色+蜂蜜水的趋性与对单一颜色基本一致,雌雄蝶对红色假花的访问次数最多,占总访花次数的30.40%,白色加蜂蜜水后访花上升到第2位(20.67%),对蓝色(17.81%)和紫色(17.10%)假花的访问次数次之(图4: B),说明金斑蝶视觉与嗅觉联合响应提高觅食能力。
金斑蝶几乎不访问单一无花冠(花色)的花枝,在无花冠的花枝上喷洒10%的蜂蜜水后,共访问146次(雌蝶65次, 雄蝶81次),与单一颜色诱集(138次)比较无显著差异(P>0.05)。综合比较视觉和嗅觉诱集蝴蝶访问,视觉+嗅觉与单一视觉诱集相比,访花总次数增加3.05倍,雌雄蝶分别增加3.31和2.86倍,而视觉+嗅觉与仅单一嗅觉诱集相比,访花总次数增加2.88倍,雌雄蝶分别增加2.95和2.83倍(图4: C),2组间无显著差异(P>0.05)。说明金斑蝶在觅食过程中,视觉和嗅觉具有相似的权重,视觉和嗅觉在觅食过程中扮演着同等重要的作用。
2.3 金斑蝶成虫求偶过程中的视觉和嗅觉行为响应
自然种群求偶行为观察发现,金斑蝶求偶有雄追雄(49次, 占56.98%)、雄追雌(32次, 占37.21%),雌追雄(4次, 占4.65%)和雌追雌(1次, 占1.16%, 偶然现象)4种相互追逐现象。雄追雄与雄追雌占主导地位,2种现象的占比差异不显著(P>0.05),远远高于其余两种现象(P<0.01)(图5)。在求偶过程中,雄蝶占主动地位,雌蝶处于被动地位。雄追雄发生率最高,可能是雄蝶之间为争夺交配权而激烈竞争。
在金斑蝶对无气味的蝴蝶标本模型求偶追逐中,同样有雄追雌(8次, 占33.33%)、雄追雄(12次, 占50.00%)、雌追雄(2次, 占8.33%)和雌追雌(2次, 占8.33%)4种相互追逐现象(图5)。模型与自然种群求偶相比较,虽然都有4种求偶模式,但金斑蝶对蝶翅模型的追逐与自然种群追逐规律不一致,雄追雌最多。
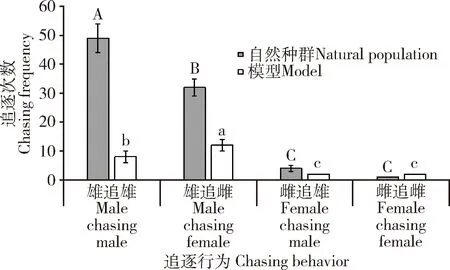
图5 金斑蝶对自然种群和标本模型求偶追逐行为Fig. 5 Chasing behavior of Danaus chrysippus adults to natural populations and models图中数据为平均值±标准差;柱上不同大写和小写字母分别表示不同自然种群间和模型间追逐次数差异显著(P<0.05, 邓肯氏多重比较)。Data in the figure are mean±SD. Capital and small letters above bars indicate significant differences in the chasing frequency among natural populations and models, respectively (P<0.05, Duncan’s multiple comparisons).
3 讨论与结论
蝶类成虫通过觅食行为获得补充营养物。在觅食过程中,蝴蝶主要利用视觉和嗅觉信号搜寻、发现、定向食物(Andersson and Dobson, 2003;mura and Honda, 2005; Kinoshitaetal., 2006; Koshitakaetal., 2008; 唐宇翀, 2015)。在觅食过程中,蝴蝶靠视觉和嗅觉寻找和识别食物资源,蝴蝶觅食视觉和嗅觉利用主要有4种方式(Tangetal., 2013):主要依靠视觉、主要依靠嗅觉、视觉和嗅觉同等利用和完全依赖嗅觉。不同种类蝴蝶视觉和嗅觉利用差异较大,在视觉利用方面,柑橘凤蝶Papilioxuthus对红色敏感(Koshitakaetal., 2004),白带锯蛱蝶对黄色敏感,而青斑蝶偏爱黄色(Tangetal., 2013)。蝴蝶对颜色的趋性可能放映了其生存环境和进化过程,本试验中金斑蝶对红色最敏感,而其主要寄主植物马利筋Asclepiascurassavica的花为花冠红色和花蕊黄色,这种颜色的趋性可能与金斑蝶觅食和识别同类有关联,金斑蝶在觅食过程中,寄主植物的花色可能驱动了以红色为主的视觉系统发育。研究表明,凤蝶、粉蝶及蛱蝶复眼中存在感觉红光波段的视觉色素,可能是四原色视觉者(Wakakuwaetal., 2004; Zaccardietal., 2006; Koshitakaetal., 2008),结合本研究的结果,推测金斑蝶复眼可能也存在感觉红光波段的视觉色素。
本研究证实,金斑蝶觅食过程中,视觉和嗅觉利用相似,但视觉和嗅觉叠加诱集蝴蝶访问数量远高于单一的颜色和气味诱集,通过视觉和嗅觉指示,蝴蝶寻找到食物资源,再通过蝴蝶的味觉感受,最终确定食物资源,完成寻找和取食食物的过程。金斑蝶觅食中视觉和嗅觉利用同等重要,而在观察中发现,金斑蝶对颜色的反应很快,而对气味的反应较慢,需要花较长时间寻找,说明金斑蝶对颜色的反应更迅速,这可能与颜色较鲜明和稳定,容易被发现,而气味复杂、不稳定、易受干扰有关。
视觉和嗅觉线索对于寻找和区分配偶都是非常重要的(Costanzo and Monteiro, 2007)。在自然种群求偶过程中,金斑蝶雄追雄和雄追雌远远多于雌追雌和雌追雄,雄追雌较多说明雄蝶能够准确地识别异性,而雄追雄最多可能是雄蝶为争夺交配权而激烈竞争,驱逐同性。在蝶翅模型的求偶试验中,雄追雌和雄追雄居多,与自然种群求偶相似,但雄追雄模型相较于自然种群所占比例更大,可能是缺乏雄蝶挥发物信息,导致竞争,驱逐同性减少;而雄追雌高于自然种群,说明通过蝶翅颜色,金斑蝶基本可以识别异性,雌蝶后翅腹面特有的白色斑点可能是识别异性的主要特征。在自然光下色彩对比中,雌雄蝶的色彩差异体现在雄蝶后翅腹面的性标上,而此性标也成为雌雄蝶在紫外光下最显著的差异,有研究表明蝴蝶翅的光谱反射率差异(自然光及紫外光)可用于异性的识别(Silberglied and Taylor, 1978; Marini-Filho and Benson, 2010),金斑蝶可能通过性标的光谱反射来识别异性。
在觅食和求偶中,金斑蝶雄蝶较雌蝶活跃,觅食过程中雄蝶访花次数多于雌蝶,求偶过程中雄蝶占有绝对的主动性,可能是由于雄蝶处于主动地位,飞行消耗更多的能量,通过多次访花获取更多的营养。
