黄氏枝膜叶蜂生物学特性及金龟子绿僵菌对其幼虫的致病力
2022-06-09曾丽琼罗明永林曦碧陈志平罗建松陈红梅蔡守平魏美才何学友
曾丽琼, 罗明永, 林曦碧, 陈志平, 罗建松, 陈红梅,蔡守平, 魏美才, 何学友,*
(1. 福建省林业科学研究院, 南方山地用材林培育国家林业和草原局重点实验室, 福州 350012; 2. 永安市林业局, 福建永安 366000;3. 明溪县林业局, 福建明溪 365200; 4. 三明市林业局, 福建三明 365000; 5. 江西师范大学生命科学学院, 南昌 330022)
乳源木莲Manglietiayuyuanensis属木兰科(Magnolinaceae)木莲属Manglietia常绿乔木,产于安徽(黄山)、浙江南部、江西、福建、湖南南部、广东北部(刘玉壶, 1996),是优良乡土速生用材、四旁绿化和庭园观赏树种,其天然林资源稀少(陈存及等, 2001; 李因刚等, 2008; 王旭军等, 2009)。福建省永安市林业局于2000年在燕东街道种植的7.5 hm2乳源木莲大径材培育示范林,2014年开始叶蜂暴发成灾,其幼虫通常几十头群集在同一叶片取食(图1),虫口密度高的单株达上千头,在数天内叶片被取食殆尽;2016年虽然利用“森得保”粉剂进行防治(邓林富, 2017),但2017和2018年仍然存在较高的虫口密度,严重影响林木的生长。危害木兰科植物的叶蜂种类较少,明确危害木兰科植物的叶蜂有2科3属7种,可能危害木兰科植物的叶蜂有4属5种,总计2科5属12种(魏美才, 2010),这12种叶蜂在我国分布于安徽、江西、广西、湖南、重庆、四川、贵州、云南,未见有危害乳源木莲的报道。牛耕耘等(2022)鉴定该取食乳源木莲的叶蜂为叶蜂科(Tenthredinidae)枝膜叶蜂属Cladiucha一新种,命名为黄氏枝膜叶蜂Cladiuchahuangbki。

图1 黄氏枝膜叶蜂幼虫危害状(何学友摄, 2019-05-08)Fig. 1 Damage symptoms of Cladiucha huangbki larvae(Photo by HE Xue-You, 2019-05-08)
利用昆虫病原微生物防治害虫研究已有较长的历史,绿僵菌是最早用于防治农业害虫的真菌,自1879年用于防治奥国金龟子以来已有140多年(蒲蛰龙和李增智, 1996),是当前研究和应用较多的虫生真菌之一,它有着广泛的昆虫寄生谱,寄主昆虫达200多种(丁苗苗等, 2009),涉及半翅目、鞘翅目、鳞翅目、膜翅目、直翅目、蜚蠊目、双翅目等,还能寄生螨类、线虫(丁苗苗等, 2009; 农向群等, 2015),且对人、畜、作物无害。近40几年来,已有近百个绿僵菌产品在十几个国家或地区获得注册(农向群等, 2015),被广泛应用于防治蝗虫、白蚁、苍蝇、天牛、叶甲、尺蛾、象甲等多种农林害虫(雷仲仁和问锦曾, 2004; 陆永跃和梁广文, 2004; St Leger and Wang, 2010; 何学友等, 2011, 2020; 刘春来, 2017; Khunetal., 2020; 郑宏, 2020),已发展成为仅次于白僵菌的真菌杀虫剂。应用真菌防治叶蜂的研究,有见对鞭角华扁蜂Chinolydaflagellicornis(肖育贵和郭亨孝, 2000)、苜蓿切叶蜂Megachilerotundata(李茂海等, 2005)、梨黏叶蜂Caliroacerasi(Aslantaetal., 2008)、小麦茎蜂Cephuscinctus(Jaronski, 2019)、苹叶蜂Hoplocampaetal., 2016)、蔷薇三节叶蜂Argeochropus(Khosravietal., 2015)等的报道,而有关黄氏枝膜叶蜂的研究,除了邓林富(2017)开展防治试验外,未见其他报道。为此,本课题组2019-2020年对其生物学特性进行研究,同时开展金龟子绿僵菌Metarhiziumanisopliae对其幼虫致病力的测定,拟筛选优良菌株,以期为黄氏枝膜叶蜂的综合防治提供基础资料和菌剂资源。
1 材料与方法
1.1 供试菌株
供试金龟子绿僵菌(以下简称绿僵菌)均来自于福建省林业科学研究院森林保护研究所,菌株具体信息见表1。
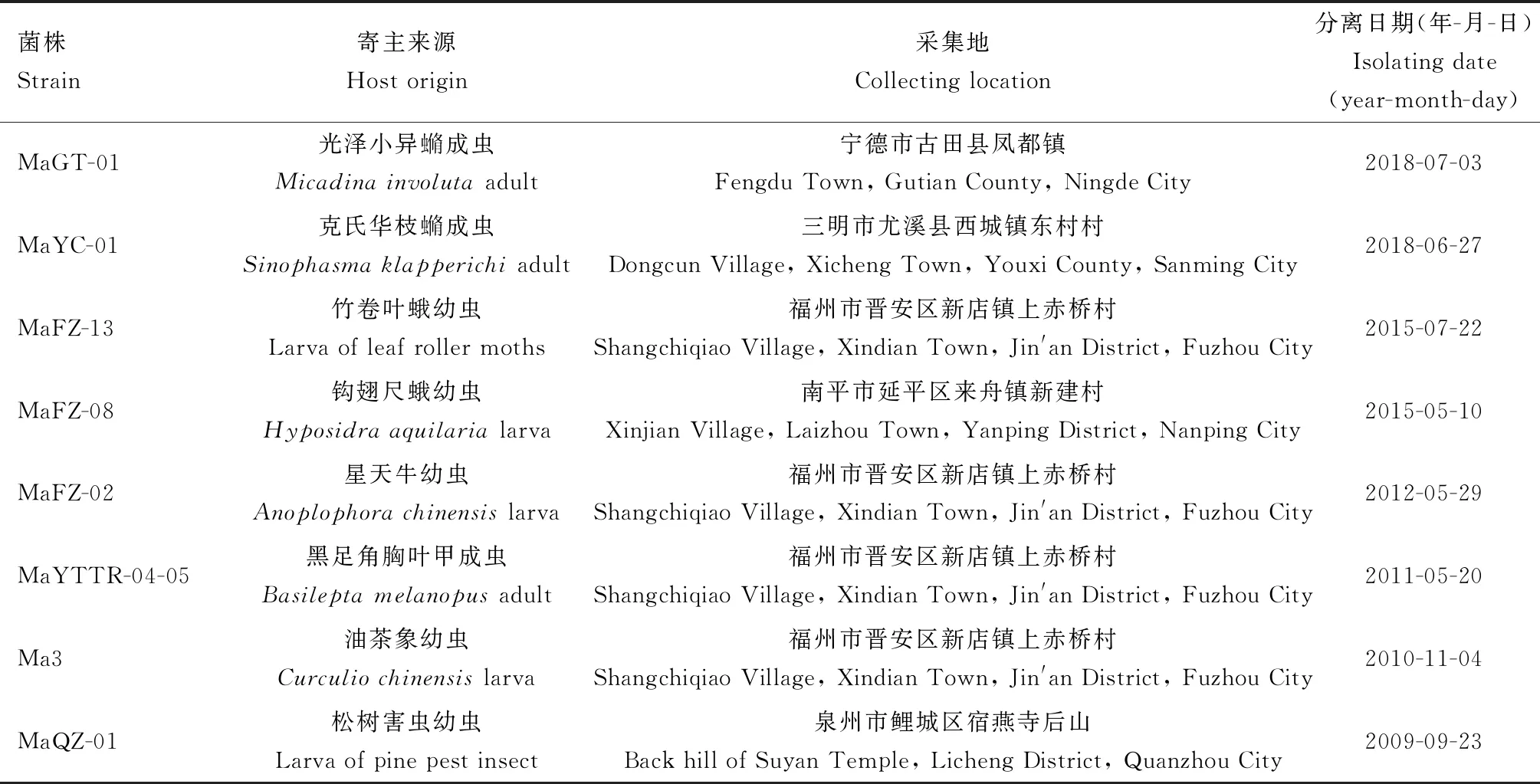
表1 供试金龟子绿僵菌菌株采集信息Table 1 Collecting data of Metarhizium anisopliae strains tested
1.2 供试昆虫
2020年5月上旬林间采集黄氏枝膜叶蜂幼虫,连枝叶一起带回实验室(室温22~29℃),在养虫笼中饲养(长×宽×高=40 cm×30 cm×60 cm),笼内放乳源木莲新鲜枝条供其取食。
1.3 叶蜂生物学特性观察
1.3.1林间调查:试验点位于福建省永安市燕东街道3林班8大班4小班,2000年种植的乳源木莲示范林,2 250株/hm2。2019和2020年3-5月,每隔3~5 d到林间巡视,观察林间黄氏枝膜叶蜂成虫、卵、幼虫的生物学特性。2019年6月-翌年2月每月观察土中蛹一次。同时,调查福建省三明、福州等地乳源木莲上黄氏枝膜叶蜂的分布情况。
1.3.2室内饲养观察:2019和2020年3-4月,从试验林捕捉黄氏枝膜叶蜂成虫带回室内,放置在养虫笼中(长×宽×高=40 cm×30 cm×60 cm),笼内放乳源木莲新鲜枝条供其产卵(室温18~25℃),枝条插于清水的三角瓶中;同时采集带卵叶片观察。在解剖镜下观察记录卵的大小、形状。林间挖取土中预蛹、蛹带回实验室,常温下置于养虫盒(长×宽×高=20 cm×15 cm×8 cm)的泥土中(保持一定湿度),盒盖上打若干个通气小孔,每天观察其化蛹、羽化情况。4-5月,采集树上幼虫叶片带回实验室,置于养虫笼中饲养,每天观察记录其生物学特性。
采集木兰科木莲属灰木莲M.glauca、木兰属Magnolia望春玉兰Magnoliabiondii以及绿化中常用的香樟(猴樟)Cinnamomumbodinieri、杜鹃Rhododendronsimsii、枫香Liquidambarformosana、榕树Ficusmicrocarpa、桂花(木犀)Osmanthusfragrans、闽楠Phoebebournei8种植物的嫩叶,分别放在养虫盒常温饲养黄氏枝膜叶蜂幼虫(室温22~29℃),每种饲养5头,每天观察幼虫取食情况并记录;同时,将8种植物嫩叶在一个养虫笼中随机摆放,并放入30头叶蜂幼虫,观察其选择取食情况。
1.4 金龟子绿僵菌对幼虫致病力测定
选取健康、活力强的室内饲养黄氏枝膜叶蜂3-4龄幼虫用于不同菌株(表1)接种试验。将供试菌株接种于PDA平板培养基上,25℃恒温培养箱培养10 d,用含0.03%吐温-80的无菌水洗下孢子,在振荡器上充分振荡使孢子团分散,用血细胞计数板计数配制成浓度为1×107孢子/mL孢子悬浮液备用。
采用喷雾法将菌悬液分别均匀喷于黄氏枝膜叶蜂虫体上,至体表湿润为止,放在滤纸吸去多余菌液,待虫体表面稍干后将虫放入装有乳源木莲嫩叶的养虫盒中饲养,置于室温观察。另设试验对照,用浓度0.03%吐温-80无菌水将叶蜂体表湿润。每处理3次重复,每重复20头幼虫,室温22~29℃,养虫盒内相对湿度80%左右。持续15 d隔天观察记录死虫数,定期更换新鲜枝条,保持乳源木莲叶新鲜;同时,将虫尸在培养皿中保湿,观察产孢和僵虫情况,明确是否被绿僵菌感染。最后统计死亡率和僵虫率,计算校正死亡率和致死中时(LT50)。
1.5 分析方法
以时间(d)的对数值为自变量x,校正死亡率的机率值为因变量y,采用机率值分析法,利用SPSS25.0计算LT50和回归方程,采用Duncan氏新复极差进行方差分析。
死亡率(%)=死亡数∕供试虫数×100;
校正死亡率(%)=(处理组死亡率-对照死亡率)∕(1-对照死亡率)×100;
僵虫率(%)=僵虫数∕供试虫数×100。
2 结果
2.1 形态特征
2.1.1成虫:雌蜂体长12~15 mm;体黑色,具金属蓝色光泽,上唇和腹部第1背板气门后侧小斑白色;足蓝黑色,具少量黄白斑;翅透明,翅痣和翅脉黑褐色;额区和单眼区刻点粗糙密集,上眶、单眼后区和小盾片中部刻点稀疏;中胸前侧片上部无明显刻点;唇基端部截型,颚眼距0.4倍于侧单眼直径;触角22+3节,第3-17节腹侧各具1明显齿突;后足基跗节约等于其后4个跗分节之和,中端部明显膨大,爪内齿显著长于外齿;前翅cu-a脉交于1M室基部,后翅臀室具柄式。雄蜂体长10~13 mm;体色和构造类似雌虫,但后足股节几乎全部蓝黑色,抱器白色;中胸侧板上部刻点稍明显;触角双栉齿状,29+1节,第3-28节各具1对长齿突(图2: A, B)。
2.1.2卵:卵整齐排列于叶背主脉两侧的表皮下(图2: C),短香蕉型,长1.95~2.05 mm,宽0.52~0.55 mm,表面光滑;初产时米白色,近孵化时灰黄色,孵化前尖端变黑(图2: D, E)。

图2 黄氏枝膜叶蜂的各虫态的形态特征(何学友摄)Fig. 2 Morphological characteristics of various developmental stages of Cladiucha huangbki(Photo by HE Xue-You)A: 雄成虫Male adult; B: 雌成虫Female adult; C: 产卵痕Oviposition mark; D: 卵块Egg mass; E: 卵Egg; F: 幼虫Larva; G: 预蛹Prepupa; H: 蛹Pupa.
2.1.3幼虫:初孵幼虫体长约3 mm,宽约1 mm,头灰褐色,体乳白色,胸背颜色稍带褐色。老熟幼虫体长25~28 mm,宽4~6 mm。头、胸足漆黑色,体土黄色,背面黑褐色,胸背和腹部第8节背面的颜色较浅。胸部背面有2~3个环褶,腹部背面有5~6个环节。胸足3对;腹足7对,第2腹节至第7腹节以及第10腹节各1对。臀节背板有一指甲盖形状黑斑,形似头部(图2: F)。
2.1.4预蛹和蛹:预蛹体背紫黑色,体长14.5~16.0 mm,宽5.5~7.5 mm(图2: G)。 蛹体长14.5~16.5 mm,宽6.0~7.5 mm,乳白色,后期逐渐转变为黄白色,羽化前变为黑褐色至黑色。雌蛹体大于雄蛹体。蛹室蚕茧形,长15.5~17 mm,宽7.5~8.5 mm,内壁光滑(图2: H)。
2.2 生物学特性
2.2.1寄主植物与分布:灰木莲、望春玉兰、香樟、杜鹃、枫香、榕树、桂花(木犀)和闽楠8种不同植物室内单独饲养黄氏枝膜叶蜂幼虫,仅灰木莲叶片当天就被大量取食,望春玉兰叶片被啃穿成孔,其他供试植物叶片均未发现咬痕;72 h后,灰木莲上的幼虫依然有3头存活,其他植物上幼虫均死亡。8种植物混合喂养幼虫情况与单独饲养类似,叶蜂幼虫主要爬到灰木莲嫩叶上取食,望春玉兰有少许被啃食痕迹,其他植物未被取食。
已知分布:福建(永安)。
2.2.2生活史:在福建永安市一年发生1代,以老熟幼虫(预蛹)在土室越冬;翌年3月上旬开始化蛹,3月下旬为化蛹高峰期,并有成虫开始羽化,4月上旬为成虫羽化和雌虫产卵高峰期,4月中旬开始幼虫出现,5月中旬老熟幼虫入土作土室越夏越冬。
2.2.3习性:成虫:成虫出土后一般在林下植被上活动、交尾、栖息,林缘的灌木、杂草上常见;晴天较活跃,飞翔力不强,容易捕捉;未见补充营养;成虫寿命6~10 d,雌虫产卵后1-2 d内死亡,雄虫寿命比雌虫多2~3 d;雌成虫交尾后,飞到树冠选择当年生完全展开的新叶产卵,产卵时多头朝叶尖方向,每次产卵1粒,产完1粒卵后稍向前移动,再产另1粒。
2019年4月9日,在林间观察生物学特性时,发现一对雄性叶蜂呈“一”字型交配(图3),在叶片上互相拖拽爬行、在空中短距离双飞,时间长达120 s以上。

图3 雄性黄氏枝膜叶蜂雄蜂同性交配中(何学友摄, 2019-04-09)Fig. 3 Same-sex sexual behavior of male adults of Cladiuchahuangbki (Photo by HE Xue-You, 2019-04-09)
卵:卵产于贴近叶背主脉两侧的表皮下,卵粒与主脉叶尖方向大多呈锐角对称整齐紧密排列成直线,产卵部位可见卵形的隆起,一侧卵数量大多30粒左右,少的10粒,多则50几粒,两列卵粒数差异多在3粒以内;产卵痕与主脉叶尖方向也大多呈锐角,浅褐色,状如缝纫走过的针线,可以作为叶片是否有卵块的快速判别依据。有卵块的主脉上有细条状黑色物质,是雌虫在产卵过程中的排泄物,整体呈浅褐色。绝大多数为靠近主脉的一次性产下的卵列,极少数卵列不靠近主脉。新产卵叶表皮未变色,3 d后叶肉组织逐渐变黄褐色。卵期13~16 d。
上一年的老叶和当年尚未展开的新叶均很少产卵。因此,在林中展叶期比成虫羽化期早的木莲单株受害较重;而当年新叶少或展叶迟的受害较轻。但暴发成灾的年份,雌虫产卵选择性少,可造成整个林分叶片食光。
随机采集3株乳源木莲样株的侧枝,调查卵在叶片上的分布情况(表2)。侧枝新叶有卵块叶片比例最高的为50%,每个卵块最少卵量20粒,最多107粒。
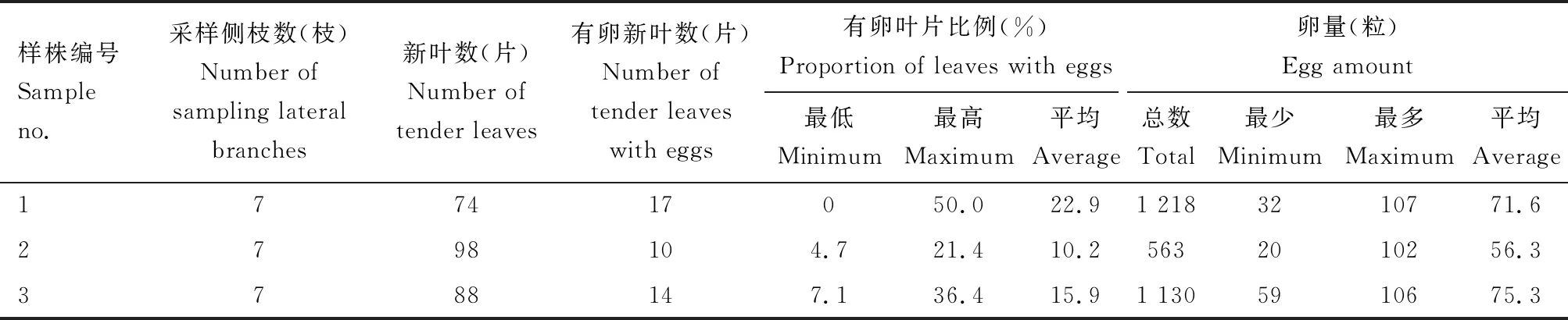
表2 黄氏枝膜叶蜂卵在乳源木莲叶片上的分布情况(福建永安市燕东街道2019年4月)Table 2 Distribution of Cladiucha huangbki eggs on leaves of Cladiucha huangbki in Yandong Street, Yong′an City, Fujian Province in April, 2019
幼虫:幼虫多在夜间孵化,孵化率96%以上。初孵幼虫有取食卵壳的习性,30 min左右以后开始取食叶背叶肉。大雨对初孵幼虫有冲刷作用。低龄幼虫食量少,随着虫龄的增大,取食量剧增,可取食全叶,大发生时将整株树叶连同叶柄也取食殆尽。幼虫有群集性,少则4~5头,多则70余头,取食时群集叶缘靠近叶尖整齐排列,多在早晚或夜间群集或分散取食。一个卵块的幼虫常群集在一起取食,数十头幼虫互相缠绕形成虫团,给人造成视觉上冲击;一片叶子食光后,群体转移到它叶危害;如整株树叶吃光后,幼虫下到地面随机爬上其他树取食危害,未找到寄主的幼虫被天敌捕食或饿死。4月中旬-5月中旬为幼虫危害高峰期,幼虫取食25~30 d后化蛹。
预蛹和蛹:5月中旬开始,老熟幼虫沿树干爬下或直接从树上掉落,大多在树干投影范围内的土壤中做土室变为预蛹越夏越冬;95%以上入土深度多在30~150 mm范围内。3月上旬开始化蛹,蛹期20 d左右。
林间天敌有蚂蚁、蜘蛛、胡峰等,也发现林间有多种鸟类活动。
2.3 高致病力金龟子绿僵菌菌株筛选
2.3.1不同菌株感染黄氏枝膜叶蜂幼虫的死亡率:幼虫被金龟子绿僵菌(1×107孢子/mL)感染2 d后,食量下降,反应迟钝,活动明显减弱,感染后虫体颜色变暗,3~4 d后逐渐僵硬,直至死亡,最后虫尸表面长出墨绿色的孢子(图4)。各绿僵菌菌株(1×107孢子/mL)感染叶蜂幼虫不同时间段的累计死亡率见图5。在接菌第3天,叶蜂幼虫累计死亡率上升速度比较平缓,其中Ma3菌株感染的效果相对较好,死亡率在25%以上,其他菌株感染不超过5%;7 d后,Ma3, MaFZ-13和MaYTTR-04-05 3个菌株感染叶蜂幼虫累计死亡率较高,在85%左右,MaYC-01和MaGT-01 2个菌株感染效果相对较差,7 d累计死亡率不到60%;感染后11 d,菌株MaQZ-01, Ma3, MaFZ-13和MaYTTR-04-05 4个菌株感染叶蜂累计死亡率达到100%,MaFZ-02和MaFZ-08感染的累计死亡率也超过96%。

图4 金龟子绿僵菌(1×107孢子/mL)感染的黄氏枝膜叶蜂幼虫僵虫(何学友摄, 2019-05-23)Fig. 4 Symptoms of Cladiucha huangbki larvae infectedwith Metarhizium anisopliae at the concentration of 1×107spores/mL (Photo by HE Xue-You, 2019-05-23)
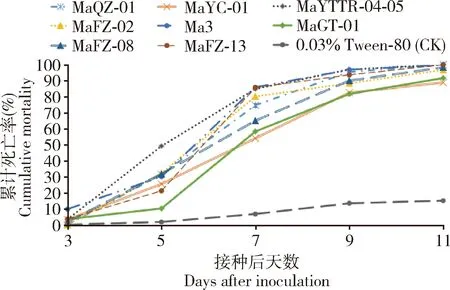
图5 不同金龟子绿僵菌菌株(1×107孢子/mL)感染后黄氏枝膜叶蜂幼虫的累计死亡率Fig. 5 Cumulative mortality of Cladiucha huangbki larvaeafter inoculation with different strains of Metarhiziumanisopliae at the concentration of 1×107 spores/mL
2.3.2金龟子绿僵菌对黄氏枝膜叶蜂幼虫的致病力比较:不同金龟子绿僵菌菌株感染黄氏枝膜叶蜂幼虫的死亡机率值与时间回归方程见表3,经检验,各回归方程的拟合度均达到了显著水平,表明时间与累计死亡率显著相关。由表3可知,感染11 d后,不同菌株对黄氏枝膜叶蜂幼虫的校正死亡率均超过85%,其中感染MaQZ-01, MaYTTR-04-05, Ma3和MaFZ-13 4个菌株的黄氏枝膜叶蜂幼虫校正死亡率为100%。僵虫率结果表明,感染MaFZ-08的黄氏枝膜叶蜂僵虫率最高,达到60%;其次是MaYTTR-04-05菌株,引起的僵虫率为50%。比较各处理的LT50可知,各菌株对黄氏枝膜叶蜂幼虫的致死中时间(LT50)都在6 d左右,其中MaYTTR-04-05菌株的LT50最短,为5.090 d;菌株Ma3的次之,为5.204 d;两者与菌株MaGT-01的LT50差异显著(P<0.05)。综合各菌株感染叶蜂幼虫的校正死亡率、僵虫率和LT50可知,MaYTTR-04-05对黄氏枝膜叶蜂幼虫致病力相对较强,具有良好的生防潜力。
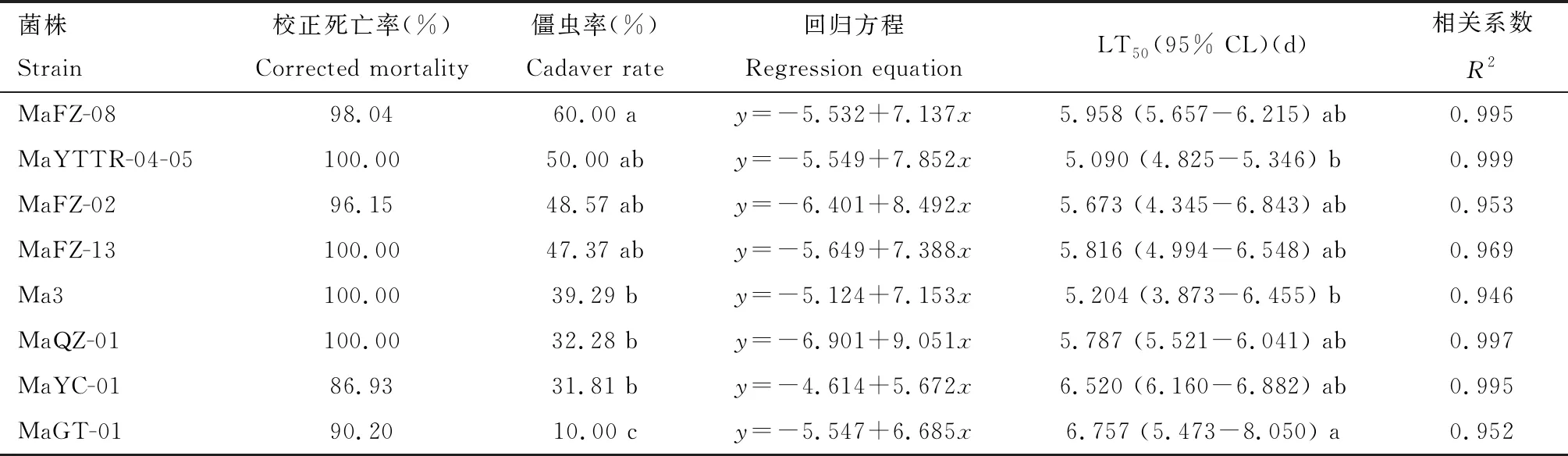
表3 不同金龟子绿僵菌菌株侵染11 d对黄氏枝膜叶蜂幼虫的致病力Table 3 Pathogenicity of different strains of Metarhizium anisopliae to Cladiucha huangbki larvae at 11 d post inoculation
3 结论与讨论
通过室内饲养与野外调查,观察了黄氏枝膜叶蜂成虫、卵、幼虫和蛹的形态特征,初步掌握了该虫的生物学特性。黄氏枝膜叶蜂在福建永安市一年发生1代,以老熟幼虫在土室中变为预蛹越夏越冬;翌年3月上旬开始化蛹,3月下旬成虫开始羽化,4月上旬为雌虫产卵高峰期,4月中旬幼虫出现,5月中旬老熟幼虫开始入土做土室越夏越冬。黄氏枝膜叶蜂幼虫主要危害木兰科木莲属植物。
关于黄氏枝膜叶蜂幼虫的分布与危害,在福建三明市(三元、沙县、明溪)、福州市等乳源木莲及其他木兰科植物分布地区也开展过调查,仅在永安市的乳源木莲示范林上发现危害,示范林周边其他植物并未被取食。研究表明,叶蜂科巨基叶蜂亚科(Megabelesesinae)的起源时间和多样化模式与其寄主植物木兰科的分化时间一致(Niuetal., 2021)。黄氏枝膜叶蜂属于巨基叶蜂亚科,本研究人工饲养表明,黄氏枝膜叶蜂幼虫仅取食木兰科木莲属植物,木兰科的其他植物在胁迫环境中只被少量取食,非木兰科植物不取食,可见幼虫仅危害木兰科木莲属植物,同时也说明昆虫与寄主植物的长期协同进化,在已适应寄主植物存在的环境中,昆虫一般不会再去主动适应(取食)其他植物。昆虫对寄主植物的选择不仅受到遗传因素的影响,还受到环境因素的影响(戴华国和孙丽娟, 2002),除了种间竞争、天敌作用等方面,起决定作用的是植物的理化性质,尤其是植物中种类繁多的次生物质(王琛柱等, 2007),而对该叶蜂的调查仅在三明市、福州市两地,是否有其他寄主植物以及其分布范围还有待进一步调查。
在观察中发现该叶蜂有雄虫同性交配现象(图3)。由于雄性昆虫从解剖学上来说并不接受雄性生殖器,因此在具有攻击性的交配过程中不合适的插入会给虫体造成损伤,因此,同性性行为在昆虫和蜘蛛物种里比较罕见,Scharf和Martin(2013)认为以下情况可能会发生同性交配:(1) 雄性在与雌性交配完后,身上携带有雌性的气味,从而向其他求偶的雄性发出混乱的信号;(2) 雄性和雌性形态非常相似以至于雄性难以区分潜在的交配对象是否是雌性,直到它骑在“她”身上并准备交配行为时才能“一辨雌雄”;(3) 利用同性交配以转移竞争对手对潜在雌性配偶的注意力;(4) 还有一种可能性,任何形式的交配都会给昆虫带来愉悦感,虽然这种可能性非常低。黄氏枝膜叶蜂属于何种同性性行为有待研究。
金龟子绿僵菌对黄氏枝膜叶蜂幼虫致病力测定结果表明,金龟子绿僵菌不同菌株对黄氏枝膜叶蜂幼虫均表现出一定的致病力,但不同菌株之间的致病力存在一定差异,其中1×107孢子/mL MaYTTR-04-05菌株感染11 d后其幼虫的校正死亡率为100%,僵虫率达50%,LT50为5.090 d,较其他菌株致病力强(表3),该菌株对黄氏枝膜叶蜂幼虫防治有较好的应用潜力。
昆虫疾病的暴发往往是许多环境因子相互结合而发生作用的结果,尤其是温度、湿度、光照等因素,同一菌株对不同寄主或同一寄主不同虫龄的致病力不同,昆虫的低龄虫态较高龄虫态易感病。针对黄氏枝膜叶蜂的防治,其幼虫在土中化蛹,将近10个月的时间预蛹都包裹在密闭的蛹室内,外界的绿僵菌对其影响有限,因此施药时机要选择幼虫下树高峰期之前,可考虑在幼虫低龄期喷粉,即在4月份中、下旬,此时永安气温通常在20~25℃,雨水也较多,林间温湿度适合绿僵菌侵染、定殖,掉落在枯枝落叶层或土壤表面的粉剂可在一定时间内保持较高致病力,黄氏枝膜叶蜂老熟幼虫在入土化蛹前在地面爬行进一步接触菌体,幼虫带菌入土,使其在土中高湿条件下继续感染致死,同时还可以让病原在土中延续繁育。在预蛹期可结合林木抚育,浅翻土壤,破坏蛹室,使预蛹死亡或被天敌取食、感染。
致病力(毒力)是菌株在一定条件下致病性大小的表现,生防真菌菌株的选择除了需考虑菌株对靶标害虫的致病力外,还需考虑其在林间的耐热、耐紫外线辐射(罗成等, 2011),以及延续情况等。有研究表明,不同菌株间抗逆性存在差异,同一菌株对不同条件的抗逆性也不相同(刘延辉等, 2013),绿僵菌在温、湿度及营养条件适合时,可继续营养生长并产孢,干燥及低温下可长期宿存于土壤,且不失去其侵染力(樊美珍等, 1991),在一定起始浓度下在土壤中能宿存几个月到一年不等(程子路等, 2003; 蔡守平等, 2016)。本研究只在室内条件下测定了不同菌株对黄氏枝膜叶蜂幼虫的致病力,菌株的抗逆性如抗紫外线、高温、干旱以及延续性和林间防治效果有待进一步研究。
