TRIM59基因沉默对结直肠癌细胞增殖和细胞周期的影响
2022-06-06高晓斌武雪亮王胜杰孙光源王文静赵轶峰刘振显
高晓斌,武雪亮,王胜杰,孙光源,王文静,梁 峰,赵轶峰,刘振显
近年来,我国结直肠癌发病率及患病率呈逐年上升趋势[1-2],尽管近年来结直肠癌的诊断和治疗技术已经取得了较大进展,但由于在早期筛查、早期诊断上的诸多困难,部分患者失去了最佳治疗时期,五年生存率并未有明显提高[3]。因此,探究结直肠癌的发生发展机制非常迫切。三结构域蛋白59(tripartite motif-containing 59,TRIM59)是一个新型的三结构域蛋白家族(tripartite motif, trim)家族成员,其主要通过介导蛋白与蛋白间的互相作用,调控蛋白稳定性,进而促进肿瘤细胞的无序化增殖,促进肿瘤发生[4]。现阶段TRIM59已被证实在肝癌[5]、非小细胞肺癌[6]等肿瘤中高表达,而TRIM59在结直肠癌中的研究鲜有报道,该研究探讨沉默TRIM59对结直肠癌细胞HCT116增殖和细胞周期的影响,旨在为结直肠癌的诊断、分子治疗提供理论基础。
1 材料与方法
1.1 主要试剂RPMI-1640培养基、胰蛋白酶和胎牛血清购于美国Hyclone公司;细胞计数试剂盒(cell counting kit-8, CCK-8)、电化学发光试剂盒以及细胞周期试剂盒购自上海碧云天生物有限公司;磷酸盐缓冲液(phosphate buffered solution, PBS)、二甲基亚砜(dimethylsulfoxide, DMSO)细胞冻存液、蛋白质裂解缓冲液(RIPA)均购于北京索莱宝生物有限公司;RNA提取试剂盒购于北京天根生化科技有限公司;SDS-PAGE凝胶制备试剂盒购于武汉博士德生物工程有限公司;靶向TRIM59的干扰质粒RNA(si-TRIM59)和阴性对照质粒(si-NC)购自上海吉玛基因科技有限公司;兔抗人TRIM59单克隆抗体、CDK4、cyclinD1抗体以及辣根过氧化物酶标记的二抗购于美国Abcam生物公司;引物由武汉擎科创新生物科技有限公司合成。
1.2 细胞培养结直肠癌细胞HCT116、SW620、SW480及人正常结直肠黏膜细胞FHC均购于中国科学院典型培养物保藏委员会细胞库,用含有10%胎牛血清的RPMI-1640培养基培养,置于37 ℃、5% CO2的恒温培养箱中培养,每隔1 d更换新鲜培养液。次日倒置荧光显微镜下观察细胞生长状况,待细胞铺满培养瓶,细胞可进行细胞传代。细胞传代步骤:吸净旧的培养基,0.25%胰蛋白酶消化细胞,收集细胞悬液与离心管中,置于离心机中以1 000 r/min离心8 min;重新混悬细胞,并将悬液于新的培养瓶内继续培养。
1.3 细胞分组及细胞转染选择处于对数生长期的HCT116细胞用于后续实验,设置TRIM59干扰组(si-TRIM59组)和阴性对照组(si-NC组)。将1×105/孔细胞量接种6孔板,培养12~24 h后,待细胞融合密度达到60%~70%时开始进行细胞转染,根据转染操作说明书,每孔转染50 nmol/L si-TRIM59或si-NC。转染完成后采用实时荧光定量聚合酶链式反应(real time quantity polymerase chain reaction, RT-qPCR)和Western blot验证质粒转染效果。
1.4 RT-qPCR实验根据TRIzol操作说明书提取HCT116细胞总RNA。使用RNA提取试剂盒提取HCT116细胞中的总RNA;利用随机引物反转录成cDNA。根据荧光定量试剂盒操作说明书进行PCR定量反应。引物序列分别为:TRIM59正义链为5′-GGACATGCTGTGTCACATCCT-3′,反义链为5′- CTCCTGTTGGTCTCATTTGGT-3′;GAPDH正义链为5′- GGAGCGAGATCCCTCCAAAAT-3′,反义链为5′- GGCTGTTGTCATACTTCTCATGG -3′。PCR反应条件如下: PCR预变性5 min,95 ℃变性30 s,然后60 ℃退火30 s,65 ℃延长60 s,共35个循环。以GAPDH作为内参,用2-ΔΔCt方法计算目的基因TRIM59mRNA的相对表达量,重复3次实验。
1.5 Western blot实验收集转染后的HCT116细胞并用细胞裂解液裂解细胞,提取总蛋白并采用BCA法测定蛋白浓度。上样后在SDS-PAGE凝胶电泳1.5 h,将分离的蛋白转移到PVDF膜上,10%脱脂牛奶封闭1 h,分别加入TRIM59抗体(1 ∶6 000)、cyclinD1抗体(1 ∶8 000)、CDK1抗体(1 ∶4 000)、STAT3抗体(1 ∶4 000)、p-STAT3抗体(1 ∶2 000)、JAK2抗体(1 ∶4 000)、p- JAK2抗体(1 ∶2 000),在4 ℃下过夜。TBST漂洗,加入辣根过氧化物酶标记的羊抗兔二抗在孵育1 h,取出PVDF膜后再TBST漂洗,用电化学发光试剂盒显影,计算目标条带的相对表达量。
1.6 CCK-8法检测细胞增殖能力吸取100 μl密度为2×104个/ml细胞悬液接种于96孔板上,待细胞贴壁生长后加入CCK-8试剂,使用酶联免疫检测仪分别在0、24、48、72 h时450 nm波长处吸光度(A)值,实验重复3次,并绘制细胞生长曲线。
1.7 克隆形成试验收集转染si-TRIM59和si-NC转染后的HCT116细胞,胰酶消化制备单细胞悬液,以500个细胞数目/孔接种于6孔板中,并在完全培养基中培养2~3周。每4 d更换1次新鲜培养基。待细胞克隆形成后,采用4%多聚甲醛固定细胞,1%结晶紫染色10 min,PBS洗涤细胞3~5次,获取图像,计算细胞克隆形成数量。
1.8 流式细胞术检测细胞周期细胞量按照5×105/孔接种在培养板中;待细胞融合密度达到60%~70%时转染细胞,胰酶消化收集细胞;以1 000 r/min离心5 min,取沉淀细胞,PBS洗涤细胞2次;经预冷的70%乙醇固定并低温下孵育3 h,离心弃固定液,调整细胞浓度为1×106/ml,加入50 μg/ml碘化丙啶(PI)染液室温下避光反应30 min进行DNA染色,最后流式细胞仪检测细胞周期分布情况,实验重复3次。
2 结果
2.1 TCGA数据库分析TRIM59在结直肠癌组织中的表达及其与患者预后的相关性采用TCGA数据库的在线网站GEPIA(http://gepia.cancer-pku.cn/)数据表明,与癌旁组织相比,TRIM59在结直肠癌组织中明显高表达,差异有统计学意义(P<0.01) (图1A);进一步通过Kaplan-Meier分析TRIM59表达与结直肠癌患者总生存期和无病生存期之间的关系,结果表明,TRIM59低表达组患者的总生存期和无病生存期较TRIM59高表达患者无明显缩短,差异无统计学意义(log rankP=0.57,log rankP=0.54)(图1B、C)。上述结果表明TRIM59在结直肠癌中高表达,但与结直肠癌患者不良预后无关。
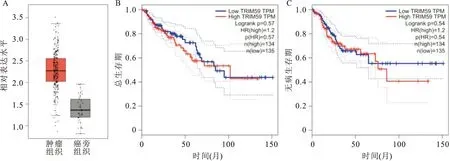
图1 TCGA数据库分析TRIM59在结直肠癌及正常结直肠黏膜组织中的表达水平及其与患者预后的相关性A:结直肠癌组织及癌旁组织中TRIM59的相对表达水平;B:Kaplan-Meier生存曲线分析TRIM59表达与结直肠癌患者总生存期的关系;C:Kaplan-Meier生存曲线分析TRIM59表达与结直肠癌患者无病生存期的关系
2.2TRIM59mRNA在结直肠癌细胞中的表达水平采用RT-qPCR检测结直肠癌细胞及正常结直肠黏膜细胞中TRIM59mRNA表达水平。结果显示,TRIM59mRNA在结直肠癌细胞HCT116、SW620、SW480中表达高于正常结直肠黏膜细胞FHC,尤其是在HCT116细胞表达最高。
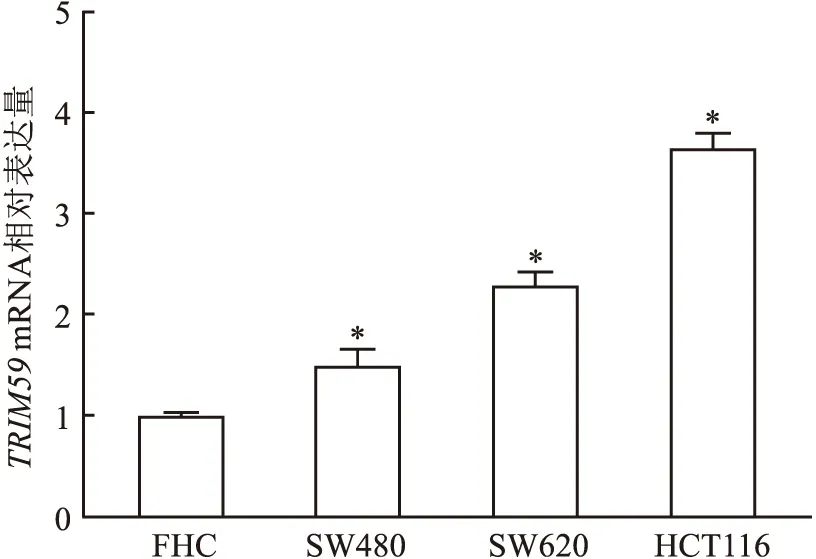
图2 qRT-PCR法检测人结直肠癌细胞中TRIM59 mRNA 的表达水平
2.3TRIM59siRNA对结直肠癌细胞中TRIM59mRNA及蛋白表达量的影响将si-TRIM59和si-NC转染HCT116细胞中,结果显示,si-TRIM59组的TRIM59mRNA表达水平和蛋白表达水平低于si-NC组(t=18.56,P<0.01)。si-TRIM59干扰组TRIM59蛋白相对表达水平低于si-NC组(t=16.93,P<0.01)。见图3。
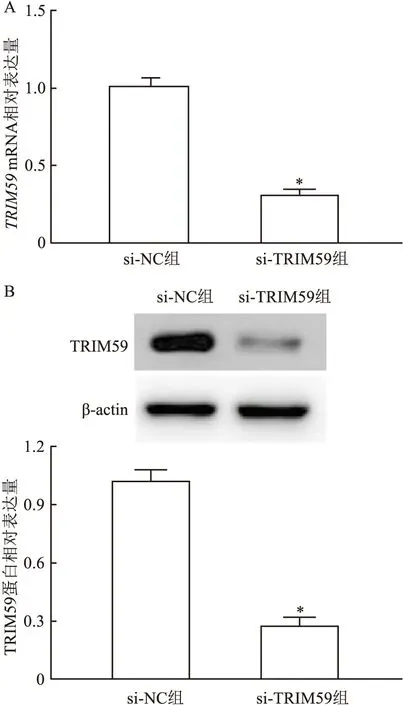
图3 qRT-PCR和Western blot检测转染TRIM59 siRNA后结直肠癌细胞HCT116中TRIM59的mRNA和蛋白表达水平A:qRT-PCR;B:Western blot;与si-NC组比较:*P<0.05
2.4 敲减TRIM59对结直肠癌细胞增殖及细胞克隆形成能力的影响CCK-8检测结果显示,在细胞转染48 h和72 h后,与si-NC组相比,敲减TRIM59表达后,HCT116细胞的增殖能力下降(48 h:t=3.939,P<0.05;72 h:t=8.828,P<0.01);细胞克隆形成实验结果表明,与si-NC组相比,敲减TRIM59后HCT116细胞集落形成数目减少(t=5.18,P<0.01),表明干扰TRIM59表达可显著抑制结直肠癌细胞的增殖能力。见图4。
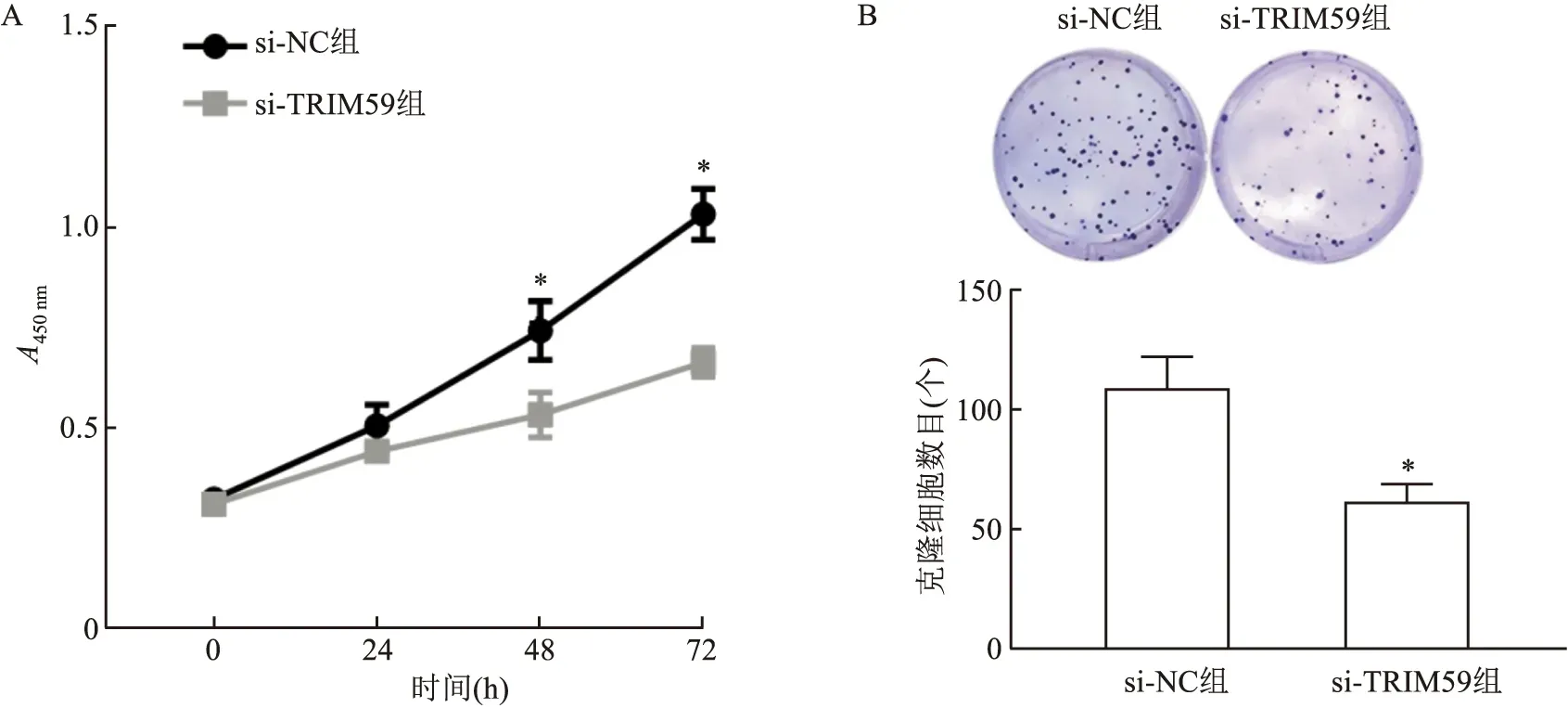
图4 CCK-8和细胞克隆形成实验检测敲减TRIM59对HCT116细胞增殖和克隆形成能力的影响A:CCK-8实验检测细胞增殖;B:细胞克隆形成实验;与si-NC组比较:*P<0.05
2.5 敲减TRIM59对人结直肠癌HCT116细胞周期的影响流式细胞术检测结果显示,与si-NC组相比,si-TRIM59组G0/G1期细胞百分比显著增加,S期比例下降,差异均有统计学意义(P<0.05)。见图5。
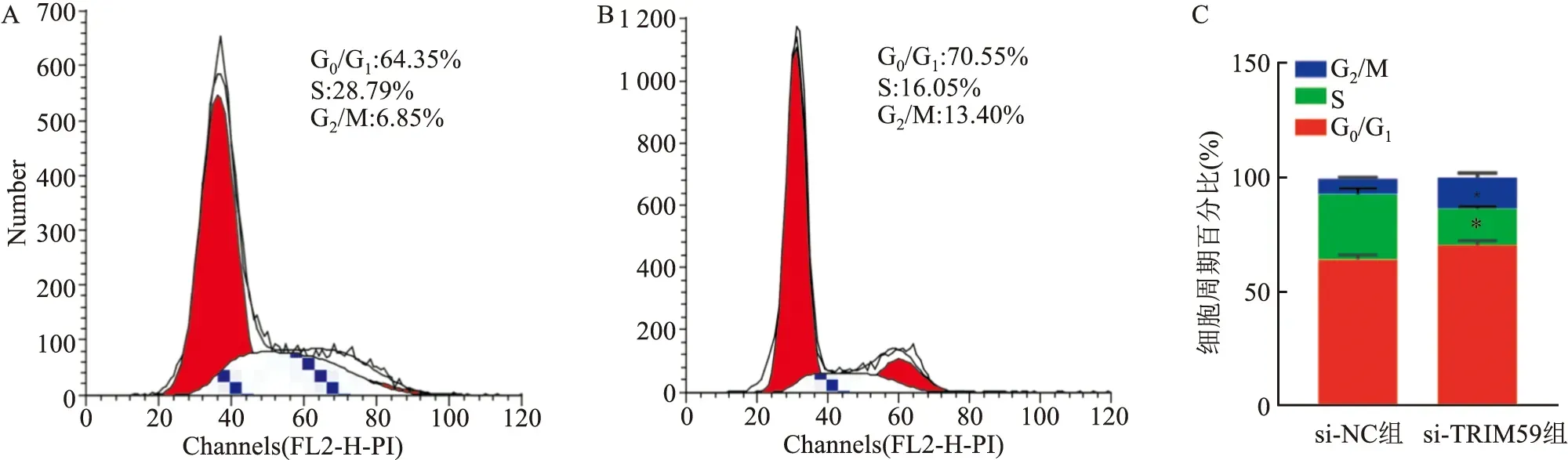
图5 流式细胞术检测敲减TRIM59对人结直肠癌HCT116细胞周期的影响A:si-NC组;B:si-TRIM59组;C:直方图;与si-NC组比较:*P<0.05
2.6 敲减TRIM59对细胞周期相关蛋白的影响采用Western blot检测敲减TRIM59对细胞周期蛋白cyclin D1和CDK4表达水平,结果显示,敲减TRIM59后HCT116中细胞周期相关蛋白CDK4和cyclin D1表达降低(t=38.41,P<0.01;t=11.65,P<0.01)。见图6。
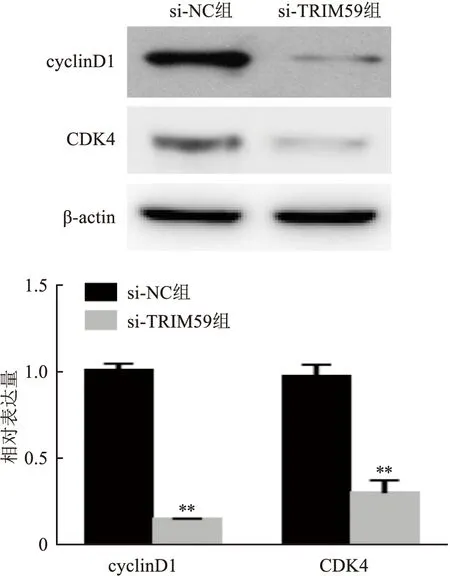
图6 Western blot检测干扰TRIM59对HCT116细胞周期蛋白的影响与si-NC组比较:**P<0.01
3 讨论
TRIM59,又称为Mrf1(mouse ring finger protein 1),属于TRIM蛋白超家族的成员之一,其主要位于人类第3号染色体上[7]。TRIM59结构包含高度保守的RBCC结构域和涉及蛋白跨膜定位的TM结构域,其主要功能是参与调控TRIM59细胞内定位[8]。TRIM59可作为原癌基因,在多种实体恶性肿瘤的侵袭、迁移、增殖和凋亡等过程中发挥重要作用[9]。Hao et al[10]研究表明,TRIM59在非小细胞肺癌(NSCLC)中的表达明显上调,其表达与NSCLC患者的年龄、性别、吸烟状态、肿瘤分期、淋巴结转移和病理分期密切性相关;此外,Cox回归分析结果表明TRIM59是NSCLC的独立预后因子。提示TRIM59可作为非小细胞肺癌预后的独立预测指标和潜在的治疗靶点。Zhou et al[11]采用qPCR和Western blot法检测50例胃癌组织和配对正常组织中TRIM59的mRNA和蛋白水平,结果表明,胃癌组织中TRIM59mRNA和蛋白水平较癌旁组织明显升高,且其高表达与胃癌患者生存时间缩短相关。在胰腺癌的研究中,Li et al[12]研究表明,TRIM59在胰腺癌组织中的表达显著增加,并且其高表达与胰腺癌患者不良预后呈正相关。Lin et al[13]研究表明TRIM59在人前列腺癌组织中高表达,干扰TRIM59表达可明显抑制前列腺癌细胞增殖和克隆形成能力,干扰TRIM59可增加S期细胞的比例,降低G2/M期细胞比例,且CDC25A、CDC2和cyclinB1三个细胞周期调节蛋白表达水平下降。
与上述结果相类似,本研究结果表明,TRIM59在结直肠癌组织及细胞中均高表达,提示TRIM59可能在结直肠癌的发生发展过程中发挥重要作用。
以结直肠癌细胞HCT116为研究对象,构建TRIM59基因siRNA载体,将此载体以瞬时转染方式抑制细胞中TRIM59mRNA表达;结果显示,敲减TRIM59的表达抑制了HCT116细胞的增殖能力和细胞集落形成能力,导致G1期比例增加,S期比例减低,提示G0/G1期细胞周期阻滞。以上研究表明,TRIM59在结直肠癌发展中发挥促进作用。
随后,本研究对TRIM59促进结直肠癌细胞增殖的相关分子机制进行初步探索。有研究[14-15]表明,CDK4作为细胞G1/S期转换的关键调控因子,它与细胞cyclin D1形成复合物后可使肿瘤细胞蛋白发生磷酸化失活,驱动细胞周期进展,从而影响肿瘤的恶性进程。本研究表明敲减TRIM59表达后可通过下调CDK4和cyclin D1表达来抑制细胞周期G1/S期转变,从而抑制细胞生长。癌基因和抑癌基因的异常表达参与细胞周期的调节,从而导致肿瘤细胞异常增殖。
