冻结及冻藏温度对小龙虾蛋白质理化性质的影响
2022-05-25杨海琦陈季旺田宏伟王海滨
杨海琦,陈季旺,3,4*,徐 言,田宏伟,廖 鄂,3,4,王海滨,3,4
(1 武汉轻工大学食品科学与工程学院 武汉 430023 2 湖北周黑鸭食品工业园有限公司 武汉 430000 3 湖北省农产品加工与转化重点实验室 武汉 430023 4 国家小龙虾加工技术研发分中心(潜江) 湖北 潜江 433100)
据农业农村部渔业渔政管理局联合全国水产技术推广总站、中国水产学会发布的《2020 中国小龙虾产业发展报告》[1]测算,2019年,我国养殖小龙虾达208.96 万t,养殖总面积达128.5 万hm2,养殖小龙虾的省份有23 个,然而产量分布不均。我国小龙虾养殖业高度集中在长江中、下游地区,说明小龙虾养殖存在地域性。同时,小龙虾养殖也有季节性。一年中,夏季是小龙虾大量上市时期,滋味鲜甜,产量较高;从秋季至第2年春季,小龙虾的产量明显下降,且个头较小,肉质较差。解决地域性和季节性问题,是小龙虾产业发展的重要课题。在长期的冻藏过程中,小龙虾肉蛋白会发生变性和氧化、聚集及降解,使蛋白质的功能特性降低。例如,表面疏水性的上升和溶解性的下降,导致品质劣变[2-4]。
国内外关于蛋白质冷冻变性机制的假设,可以归纳为“水和水合水的相互作用学说”“细胞液的浓缩学说”以及“结合水的分离学说”,然而尚未有定论[5]。Cheng 等[6]比较了分别经压力辅助冷冻、变压冷冻和浸渍冷冻的天然肌动球蛋白的构象和功能变化,探究高压冷冻影响对虾中提取的天然肌动球蛋白变性的机理。结果显示,变压冷冻能显著降低天然肌动球蛋白的变性程度,在变压冷冻过程中,冰晶的形成对天然肌动球蛋白的二级和三级结构影响显著。Liu 等[7]为了探究浸渍冷冻温度对斑鱼肉中冰晶形成和蛋白质特性的影响,将鱼肉分别在-20,-30,-40 ℃共3 个温度下浸渍冻结和-20 ℃下空气冻结,测定鱼肉中的冰晶大小及蛋白质的二级结构和热稳定性。结果显示,较低浸渍冷冻温度可以更好地保持蛋白质的二级结构,并提供更高的蛋白质热稳定性。Shi 等[8]为了探究冻结温度 (-18,-30,-80 ℃) 和时间(1,4,12,24周)对小龙虾品质的影响,测定了冻藏小龙虾的三磷酸腺苷酶(ATPase)活性、肌原纤维蛋白含量、盐溶性蛋白质含量、总巯基含量、质构、持水性和水分含量,结果显示,ATPase 活性和肌原纤维蛋白含量显著降低,持水率显著上升,冻结温度显著影响小龙虾的品质。目前已知冻结速度、冻藏温度和添加剂等冷冻加工方式显著影响冷冻过程中蛋白质的理化性质,且不同种类水产品的品质受冷冻加工条件影响的差异明显[9-10]。
目前,工业化生产中常采用液氮冻结或者鼓风冻结小龙虾原料及其制品,而冻结温度通常依靠经验确定,有关冻结温度、冻藏温度对冻藏小龙虾品质及蛋白质理化性质的影响研究尚未见报道。本课题组前期试验发现,冻结及冻藏温度显著影响小龙虾冻藏过程中的品质。较低的冻结温度对肌纤维的破坏程度较轻;较低冻藏温度的小龙虾肉中结合水和不易流动水的比例较高,小龙虾肉的汁液流失率、剪切力、pH 值、TVB-N 值较低,持水率较高。本研究探究经不同冻结温度处理和冻藏温度贮藏的小龙虾肉的肌原纤维蛋白含量、表面疏水性、内源荧光强度、巯基和二硫键含量、二级结构的变化,结合冰晶的显微结构观察结果,探讨冻结、冻藏温度对小龙虾肉蛋白质理化性质的影响,为小龙虾加工原料的周年供应提供理论指导。
1 材料与方法
1.1 材料与试剂
鲜活小龙虾(20~30 g/只),湖北周黑鸭食品工业园有限公司;氯化钾(分析纯)、氯化钠(分析纯)、盐酸(分析纯)、2,4-二硝基苯肼(DNPH)、尿素(分析纯)等,国药集团化学试剂有限公司;乙二胺四乙酸(EDTA)、十二烷基磺酸钠(SDS)、5,5'-二硫代双(2-硝基苯甲酸)(DTNB)、三羟甲基氨基甲烷(Tris)、磷酸缓冲盐溶液(PBS,0.2 mol/L)、四甲基乙二胺 (TEMED)、1-苯胺基-8 奈基磺酸盐(ANS)等,德国Biofroxx 公司。
1.2 仪器与设备
DW-60W388 超低温冰柜,青岛海尔生物医疗有限公司;SSN-61 热电偶温度采集仪,深圳宇问温度系统传感有限公司;冷冻干燥机,北京博医康实验仪器有限公司;XHF-DY 高速分散器,宁波新芝生物科技有限公司;7200 型可见光分光光度计,上海尤尼柯仪器有限公司;GL-20-Ⅱ高速冷冻离心机,上海安亭科学仪器厂;NEXUS670 傅里叶红外光谱仪,美国尼高力仪器公司;F-4600 荧光分光光度计,日本Hitachi 公司。
1.3 试验方法
1.3.1 冻藏小龙虾的制备 将挑选好的鲜活小龙虾用自来水刷洗,超声波清洗20 min(水的添加量以覆盖虾体为准),放入沸水中热烫1 min,装盒(每盒1 kg 小龙虾),盒中灌入清水(水的添加量以覆盖虾体为准),将盒抽真空。盒装小龙虾分别置于-20,-40,-55 ℃的冰柜中冻结至中心温度达到-15 ℃。将装盒虾随机分为2 组,分别置于-20℃和-40 ℃冰柜中冻藏,冻藏周期为24 周,每隔6周取样用于检测。每次随机取约1 kg 小龙虾,使用流动水解冻。取虾肉时去除头、壳和肠线,冷冻干燥后粉碎备用。
1.3.2 肌原纤维蛋白含量的测定 参考Pazos 等[11]的方法。准确称取3 g 小龙虾肉粉于50 mL 离心管中,加入30 mL(10 倍体积)Tris-HCl 缓冲液(10 mmol/L,pH 7.5),用高速分散器匀浆,溶液经冷冻离心(4 ℃,10 000 r/min)20 min 后,取沉淀,加 入30 mL Tris-HCl 缓冲液(10 mmol/L,pH 7.0),匀浆后离心(4 ℃,10 000 r/min)20 min 后取上清液,即为肌原纤维蛋白溶液。
用0.6 mol/L KCl 溶液将肌原纤维蛋白溶液稀释10 倍,取1 mL 上清液,用双缩脲法测定其浓度。
1.3.3 肌原纤维蛋白表面疏水性的测定 参照Haskard 等[12]的方法,使用荧光探针ANS 确定表面疏水性。取已知浓度的肌原纤维蛋白溶液,采用Tris-HCl 缓冲溶液 (0.6 mol KCl-20 mmol Tris-HCl,pH 7.0) 稀释至质量浓度为0.1,0.2,0.3,0.5 mg/mL 的溶液。分别取上述稀释液2 mL 置于5 mL PCR 离心管,加入10 μL ANS 溶液(8 mmol/L,溶解在0.1 mol/L pH 7.0 PBS 溶液中),涡旋混匀。采用荧光光谱仪测定,操作条件为:激发波长385 nm,扫描范围400~700 nm,扫描速度1 200 nm/min,激发狭缝和发射狭缝宽均为2.5 nm,响应时间0.1 s,记录470 nm 波长处的荧光发射强度,以蛋白质浓度为横坐标,荧光发射强度为纵坐标做图,曲线初始段的斜率(通过线性回归分析计算)即为蛋白质分子的表面疏水性指数(H0)。
1.3.4 肌原纤维蛋白内源荧光强度的测定 参考Cao 等[13]的方法。用0.6 mol/L KCl 溶液将肌原纤维蛋白溶液稀释至浓度为0.05 mol/L,采用荧光分光光度计进行波长扫描,设定参数:激发波长295 nm,狭缝宽度2.5 nm,波长扫描范围280~400 nm,扫描速度12 000 nm/min。
1.3.5 肌原纤维蛋白总巯基和二硫键含量的测定 参照Beveridge 等[14]、Oliver 等[15]的方法,稍作修改。
1)总巯基含量 总巯基A 液:分别称取尿素480.48 g、EDTA 2.9224 g、SDS 20 g、Tris 24.23 g溶于400 mL 水中,用6 mol/L 盐酸调节pH 值至7.0,定容至1 L。
总巯基B 液:分别称取DTNB 1.0 g、Tris 24.23 g 溶于800 mL 水中,用6 mol/L 盐酸调节pH 值至8.0,定容至1 L。
测量:取1 mL 肌原纤维蛋白提取液,采用Tris-HCl 缓冲溶液 (0.6 mol KCl-20 mmol Tris-HCl,pH 7.0)稀释至质量浓度为0.2 mg/mL,加入9 mL 的总巯基A 液,涡旋混匀后在室温下放置30 min,取4 mL 混合液,加入0.4 mL 总巯基B 液;在40 ℃水浴反应25 min 后,测定波长412 nm 处的吸光值,以0.2 mol/L PBS 为空白对照。
2)二硫键含量 二硫键A 液:分别称取480.48 g 尿素、2.9224 g EDTA、20 g SDS、15.826 g K2SO3溶于400 mL 水中,用6 mol/L 盐酸调节pH值至8.0,定容至1 L。
二硫键B 液:分别称取1 g DTNB、15.826 g K2SO3溶于800 mL 水中,用6 mol/L 盐酸调节pH值至9.5,定容至1 L。
测量:取1 mL 肌原纤维蛋白提取液,采用Tris-HCl 缓冲溶液 (0.6 mol KCl-20 mmol Tris-HCl,pH 7.0)稀释至质量浓度为0.2 mg/mL,加入9 mL 的二硫键A 液,涡旋混匀后,在室温下放置30 min。取4 mL 混合液,加入0.4 mL 二硫键B 液,在40 ℃反应25 min 后,测定波长412 nm 处的吸光值,以0.2 mol/L PBS 为空白对照。
1.3.6 肌原纤维蛋白二级结构的测定 参照Sun等[16]的方法,稍作修改。小龙虾肉粉过100 目筛后,取1 g 装于10 mL PCR 离心管中备用。将虾肉粉与无水溴化钾按质量比1∶100 混合压片,做全波段扫描测定,空白对照为溴化钾。分辨率为4 cm-1,扫描累加32 次,光谱观察范围为400~4 000 cm-1。利用Peakfit 软件进行分析,首先进行基线校正,然后用Deconvolve Gaussian 曲线去卷积,进行二阶导数拟合,直至拟合相关系数稳定不小于0.99 为止。确定各子峰与各二级结构的对应关系,计算各子峰面积的相对百分含量。
1.3.7 冰晶的显微结构观察 参照Yang 等[17]的方法,分别用冷却至-20 ℃和-40 ℃的刀片,将-20℃和-40 ℃的冻藏小龙虾肉切成1 mm×1 mm×1 mm 左右的正方体(沿肌肉纤维纹理切割以便观察)。将-20 ℃和-40 ℃冻藏小龙虾肉置于由30 mL 60%无水乙醇、30%三氯甲烷和10%冰醋酸组成的混合溶液中 (混合溶液已分别降温至-20 ℃和-40 ℃),分别在-20,-40 ℃的冰柜中贮藏18 h后,从冰柜中取出,自然升温至25 ℃,用无水乙醇脱水2 h,然后在正丁醇溶液中脱水2 次,每次2 h,最后浸泡在正丁醇溶液中8 h。洗脱的虾肉块再用甲苯冲洗3 次,每次30 min,然后浸入57 ℃液体石蜡中3 次,每次1 h,最后用模具将每个虾肉块包埋在石蜡中,并用切片机沿肌肉纤维纹理切割成10 μm 厚的切片。将切片和水一起放置在玻片上,置于56 ℃温板上熔化石蜡。将切片浸入甲苯中2 次去除石蜡,每次10 min,然后浸入无水乙醇中2 次,每次10 min,将虾肉片密封在玻片上。用光学显微镜放大100 倍观察并拍照。
1.4 数据处理
每组数据重复测定3 次,结果用“平均值±标准差(SD)”表示。应用SPSS 软件对数据进行统计分析,采用ANOVA 进行方差分析、Duncan 多重极差检验比较平均值在显著性水平上的差异,P>0.05 判定为差异不显著,P<0.05 判定为差异显著。
2 结果与分析
2.1 小龙虾冻藏过程中肌原纤维蛋白含量的变化
由表1可以看出,冻藏温度相同时,随着时间的延长,6 组小龙虾肉的肌原纤维蛋白含量均呈现先显著减小(P<0.05)后稳定的趋势,这与Li等[18]的研究结果类似。在冻藏过程中,冰晶不断增大,肌肉组织中细胞液不断被浓缩,使离子浓度上升、pH 值发生显著变化(P<0.05),导致肌原纤维蛋白变性;在冻藏过程中,脂质自动氧化等化学反应中可能会产生氧自由基,氧自由基易攻击肌肉细胞和膜蛋白,使蛋白质发生氧化反应而变性,部分变性后的蛋白质由于结构发生改变,更容易被酶水解[19-20]。同时,蛋白质可能受到微生物的作用被分解[21]。

表1 小龙虾冻藏过程中肌原纤维蛋白含量的变化(mg/mL)Table 1 Change in myofibrillar protein content of red swamp crayfish during freezing storage (mg/mL)
冻藏温度相同时,在整个冻藏周期内,-20 ℃冻结的小龙虾肉肌原纤维蛋白的含量显著低于-40 ℃和-55 ℃冻结的小龙虾(P<0.05),而-40℃和-55 ℃无显著差异(P>0.05)。这是由于-20 ℃冻结的速度较慢(见图1),使更多水分被冻结,更多与蛋白质结合的水分被迫迁移,导致蛋白质脱水程度更深(水分状态的结果验证了这一点,见图2),同时,更多水分被冻结使得蛋白质周围环境的溶质被浓缩,使得pH 值和离子浓度变化更大,加快了蛋白质的变性[22]。
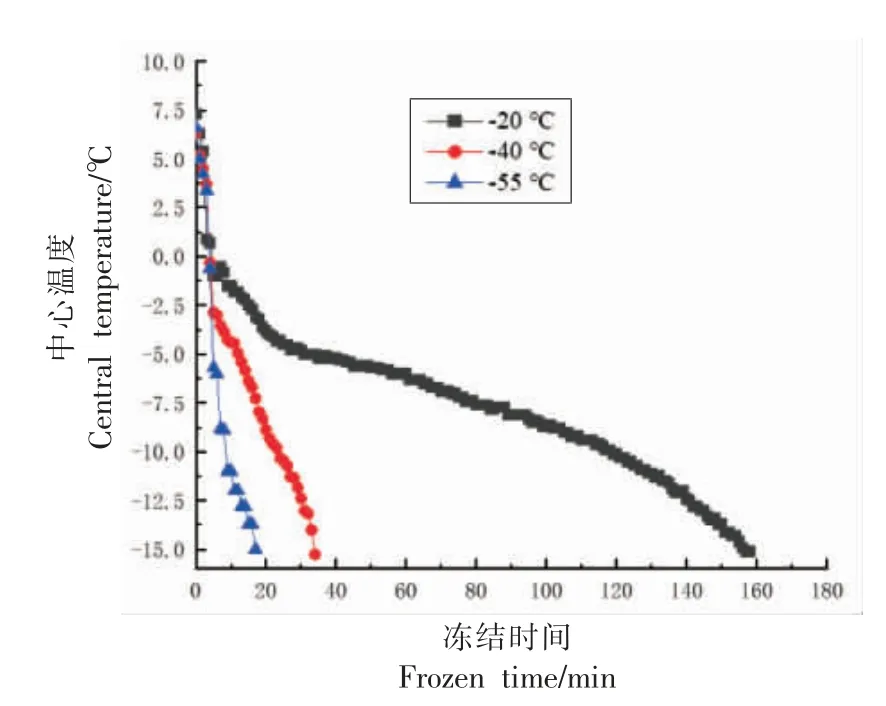
图1 小龙虾冻结过程中心温度的变化Fig.1 Change in central temperature of red swamp crawfish during frozen
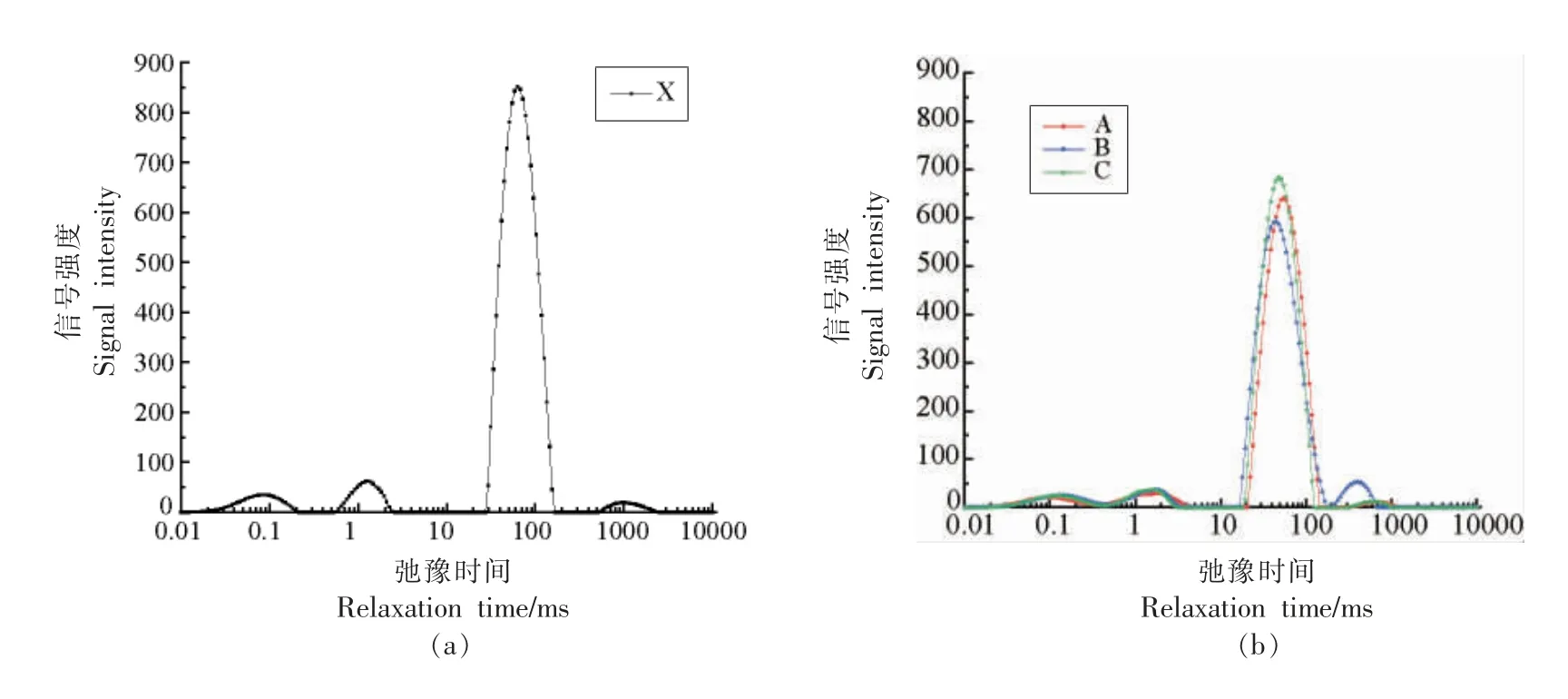

图2 小龙虾冻藏过程中水分状态的变化Fig.3 Change in moisture state of red swamp crawfish during freezing storage
冻结温度相同时,在整个冻藏周期内,-20 ℃冻藏的小龙虾肉的肌原纤维蛋白含量显著低于-40 ℃冻藏的小龙虾(P<0.05)。这是由两方面原因造成:一是-20 ℃冻藏的小龙虾的脂质自动氧化程度较重,产生更多的氧自由基,导致蛋白质的氧化变性更明显(脂质氧化的试验可验证,结果未显示);二是-20 ℃冻藏条件下抑制微生物生长繁殖能力较弱,导致微生物分解了更多的蛋白质[21]。
2.2 小龙虾冻藏过程中肌原纤维蛋白表面疏水性的变化
由表2可以看出,随着冻藏时间的延长,6 组小龙虾肉的肌原纤维蛋白的表面疏水性均呈先显著增大(P<0.05)后稳定的趋势。因为,新鲜的小虾肉疏水基团多埋在蛋白质分子内部,表面疏水性较低;冻藏开始后,肌原纤维蛋白分子逐渐展开,空间构象发生改变(内源荧光强度的结果验证了这一点),使得疏水基团暴露,表面疏水性上升;外露的疏水基团越来越多,疏水相互作用力增大,部分蛋白质可能发生聚集,导致疏水性轻微下降(P>0.05),最后趋于平稳[23]。
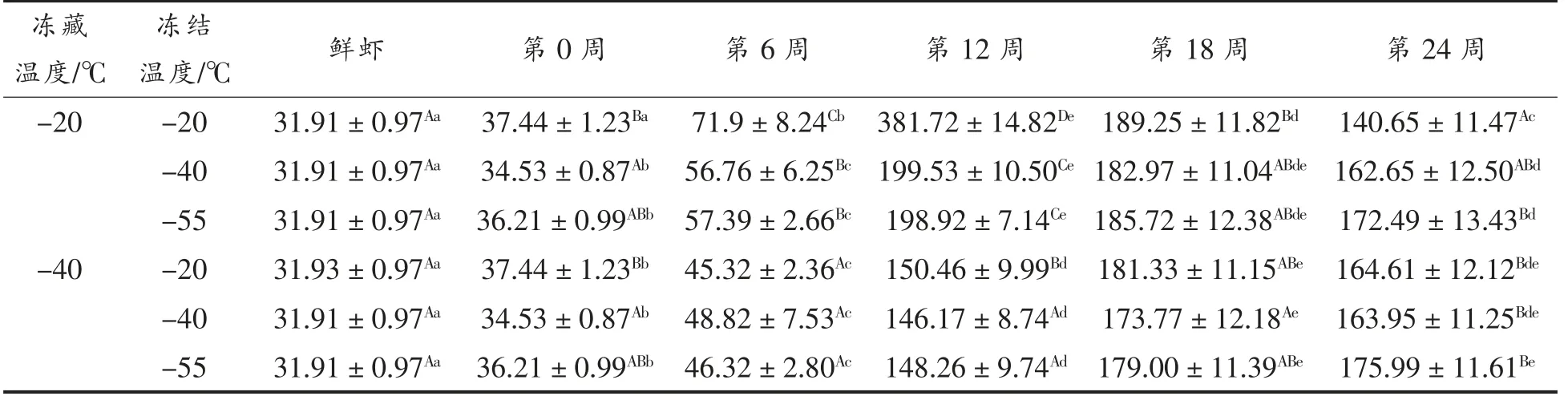
表2 小龙虾冻藏过程中表面疏水性指数(H0)的变化Table 2 Change in surface hydrophobicity index of red swamp crayfish during freezing storage (H0)
冻藏温度相同时,冻结温度对小龙虾肉肌原纤维蛋白的表面疏水性影响不显著(P>0.05)。冻结温度相同时,冻藏温度显著影响了小龙虾肌原纤维蛋白的表面疏水性(P<0.05):-20 ℃冻藏的小龙虾表面疏水性在第12 周升至最高,然后趋于稳定;而-40 ℃冻藏的小龙虾肌原纤维蛋白的表面疏水性在第18 周才达到最高值,然后趋于稳定。因为,较低的冻藏温度抑制了虾肉中水分的重结晶,减少了结合水向自由水迁移的比例(水分状态的结果证明了这一点,见图2),从而降低了蛋白质周围pH 值和离子强度的变化程度,使蛋白质空间构象更稳定。
2.3 小龙虾冻藏过程中肌原纤维蛋白内源荧光强度的变化
由图3可以看出,冻藏温度相同时,6 组小龙虾肉在冻藏第6~12 周时,荧光强度逐渐增大,在冻藏第12~24 周时,荧光强度逐渐减小。这表明在冻藏期间,色氨酸的微环境在不断变化。因为在冻藏早期,冰晶增大,损伤肌纤维结构,使得肌原纤维蛋白的天然排列方式被破坏,肌原纤维蛋白展开使得埋在内部的色氨酸残基暴露,荧光强度增大;冻藏中后期,随着冰晶的增大,在加重了对肌纤维机械损伤的同时,也使得离子浓度和pH 值上升,越来越多的疏水基团和巯基氧化形成的二硫键暴露,使得分子间作用增强,蛋白质发生再聚集,导致色氨酸残基重新被包裹在内部,使得荧光强度下降[21]。这也验证了表面疏水性的结果。

图3 小龙虾冻藏过程中内源荧光强度的变化Fig.3 Change in intrinsic fluorescence intensity of red swamp crayfish during freezing storage
冻藏温度相同时,3 种温度冻结的小龙虾肉的内源荧光强度无明显差别(P>0.05);冻结温度相同时,-20 ℃冻藏温度下小龙虾的荧光强度在整个冻藏过程中的变化大于-40 ℃,这与Ren 等[24]的研究结果类似。内源荧光强度的变化与表面疏水性一致,可能的原因类似。
2.4 小龙虾冻藏过程中肌原纤维蛋白总巯基和二硫键含量的变化
由图4、图5可以看出,随着冻藏时间的延长,6 组小龙虾肉肌原纤维蛋白的总巯基含量均呈先显著下降(P<0.05)后逐渐稳定的趋势,而二硫键含量呈先显著增大(P<0.05)后趋于稳定的趋势,二硫键含量的变化趋势与总巯基含量基本相反,跟李志鹏等[25]的研究结果类似。这可能是冰晶产生的机械损伤作用,使肌原纤维蛋白的结构被破坏,内部巯基暴露出来,进一步被氧化形成二硫键。
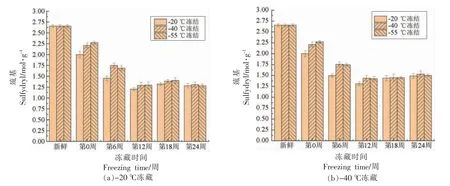
图4 小龙虾冻藏过程中总巯基含量的变化Fig.4 Change in total sulfhydryl content of red swamp crayfish during freezing storage
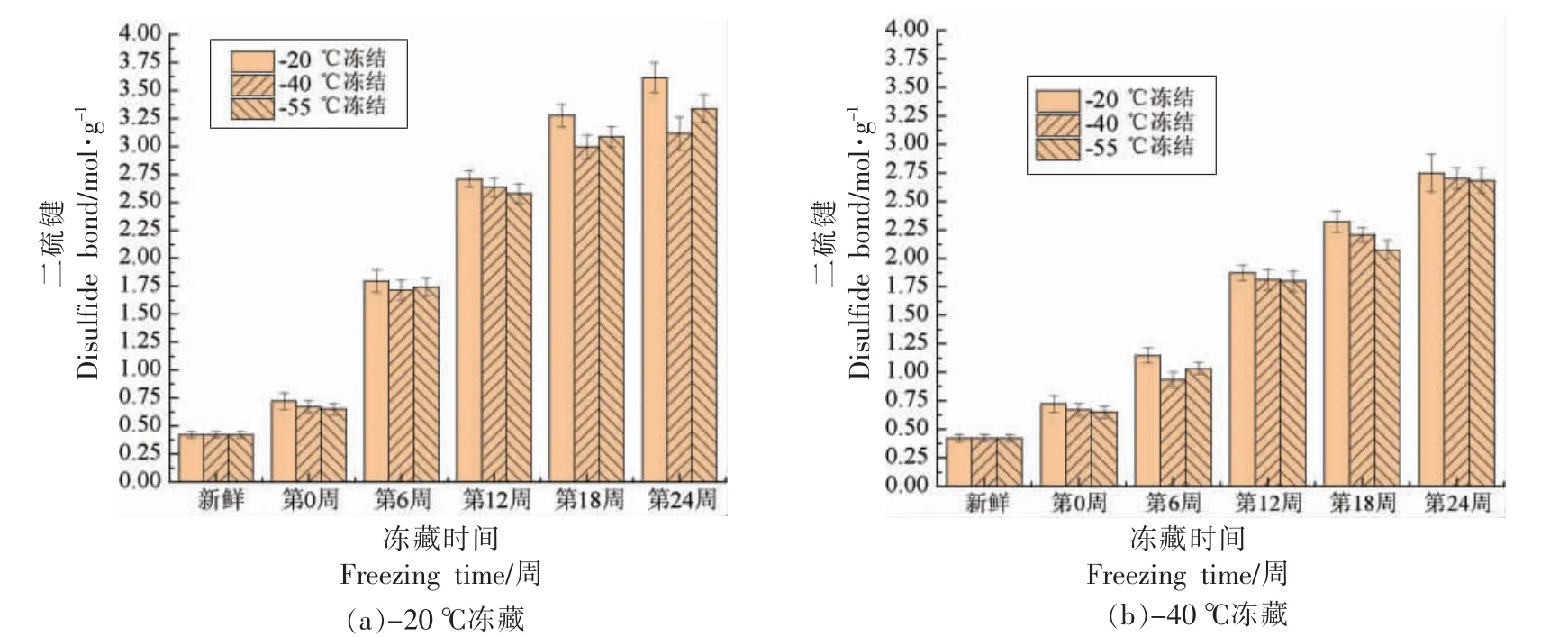
图5 小龙虾冻藏过程中二硫键含量的变化Fig.5 Change in disulfide bond content of red swamp crayfish during freezing storage
冻藏温度相同时,在第0~12 周,-20 ℃冻结的小龙虾的巯基含量显著低于-40,-55 ℃冻结,而二硫键含量显著高于-40,-55 ℃冻结(P<0.05);在第12~24 周,小龙虾的巯基和二硫键含量变化不显著(P>0.05)。因为,在冻藏中前期,-20 ℃冻结的小龙虾形成了更大的冰晶,对肌原纤维蛋白的结构破坏更严重,导致更多的巯基暴露,与氧接触而被氧化;而在冻藏后期,越来越多巯基氧化形成二硫键,增强了分子间作用,使得蛋白质发生再聚集,减轻了巯基被进一步氧化的程度,导致巯基和二硫键含量变化不显著。
冻结温度相同时,冻藏温度显著影响了小龙虾的巯基和二硫键含量(P<0.05):-20 ℃冻藏巯基含量显著低于-40 ℃冻藏,二硫键含量显著高于-40 ℃冻藏(P<0.05)。这是由于-40 ℃抑制了水的重结晶,降低了蛋白质空间结构被破坏程度,减少了巯基的暴露。同时,-40 ℃冻藏的小龙虾肉脂质的自动氧化速度较低,产生的氧自由基较少,减轻了肌原纤维蛋白被氧自由基氧化的程度[26-27]。
2.5 小龙虾冻藏过程中的肌原纤维蛋白二级结构的变化
由表3可以看出,随着冻藏时间的延长,6 组肌原纤维蛋白的α-螺旋的相对含量逐渐下降(P<0.05),β-转角的相对含量逐渐上升(P<0.05);β-折叠的相对含量呈先上升后下降的趋势,无规则卷曲的相对含量呈先下降后上升的趋势。天然肌原纤维蛋白分子的α-螺旋多位于多肽链内部,是一种紧密且无空腔的稳定结构;β-折叠是比α-螺旋更稳定的伸展结构,β-转角和无规则卷曲结构的紧密程度和构象稳定性远低于α-螺旋[28-29]。肌原纤维蛋白的α-螺旋相对含量下降主要是由肌球蛋白尾部结构变化所致[30]。天然肌球蛋白的杆状尾部主要二级结构是α-螺旋,当其逐渐解旋而部分转化成β-转角和无规卷曲时,肌原纤维蛋白结构的紧密程度和构象稳定性大幅度降低,包埋于分子内部的疏水性残基暴露出来,增强了分子间的疏水相互作用力,使肌原纤维蛋白聚合(这也验证了表面疏水性的变化)[31]。

表3 冻结小龙虾冻藏过程中肌原纤维蛋白二级结构的变化(%)Table 3 Change in secondary structure of myofibrillar protein from red swamp crayfish frozenduring freezing storage (%)
在冻藏温度相同时,-20 ℃冻结的小龙虾肉肌原纤维蛋白的α-螺旋的相对含量低于-40 ℃和-55 ℃,β-折叠、β-转角和无规卷曲的相对含量无明显差别。这是因为-20 ℃冻结使小龙虾肉中形成的冰晶更大,对肌原纤维蛋白造成的机械损伤更严重。在冻结温度相同时,-40 ℃冻藏的小龙虾肉肌原纤维蛋白的α-螺旋的相对含量明显高于-20 ℃,而β-折叠、β-转角和无规卷曲的相对含量无明显差异,可能是-40 ℃抑制了重结晶和微生物的生长,减轻了肌纤维被破坏的程度,使-40 ℃冻藏的小龙虾肉的肌原纤维蛋白的二级结构具有更好的稳定性[21]。
2.6 小龙虾冻藏过程中冰晶结构的变化
由图6可以看出,未经冷冻的小龙虾肉组织排列有规则且紧密,组织边缘光滑,组织间形成有规律且清晰的间隙。冷冻后的小龙虾肉组织变化非常明显,6 组小龙虾肉组织均出现冰晶孔隙,对组织造成不同程度的损伤。随着冻藏时间的延长,冰晶孔隙不断扩大。在第12 周和24 周,冰晶孔隙变化最明显,甚至形成拉长的针形冰晶孔隙。

图6 小龙虾冻藏过程中冰晶微观结构的变化Fig.6 Changes in the ice crystal microstructure of red swamp crawfish during the freezing storage
在冻藏温度相同时,-20 ℃和-40,-55 ℃冻结的小龙虾肉的冰晶孔隙差异明显,-40,-55 ℃之间差异相对较小。在刚结束冻结后,-40 ℃和-55 ℃冻结的虾肉的冰晶孔隙分布均匀、细小、圆润且数量较多,-20 ℃冻结的数量较少而孔径较大且不均匀。这可能是由于-20 ℃冻结的降温速度较慢,通过冰晶生成区(-1~5 ℃)的时间较长,导致形成较大的冰晶。在整个冻藏周期中,-20 ℃冻结较其它两种冻结温度的冰晶孔隙增大更明显,这与向迎春等[32]的研究结果类似,可能是-20 ℃冻结形成的冰晶较大,对肌纤维的破坏更严重,使得细胞内电解质渗透进细胞膜,引起细胞膜收缩,增大了胞外空间,导致冻藏过程中,冰晶更容易相互结合而增大。
在冻结温度相同时,-40 ℃和-20 ℃冻藏的小龙虾肉的冰晶孔隙差异明显。-20 ℃冻藏的冰晶孔隙相对较大,冻藏至12 周时有针形冰晶的形成,导致组织间隔不均。这可能是在-20 ℃冻藏时,整个体系处于橡胶态,虾肉内的黏度较低,使得水分子移动能力增强,冰晶容易在冻藏过程中结合,产生重结晶现象,而-40 ℃的冻藏温度更接近玻璃态,水分子移动能力较弱,故而冰晶较小。这些结果进一步验证了冻结及冻藏温度对小龙虾肉肌原纤维蛋白的表面疏水性、内源荧光强度、二级结构等的影响。
3 结论
-40 ℃和-55 ℃冻结的小龙虾肉中形成的冰晶更小,使小龙虾肉的离子浓度更低,pH 值更稳定,减轻了小龙虾肉蛋白的变性;-40 ℃冻藏更好的抑制了重结晶,减轻了肌纤维被破坏的程度。同时,-40 ℃冻藏降低了脂质自动氧化的速度,减轻了蛋白质被自由基氧化的程度,使小龙虾肉蛋白的空间构象更稳定。这表明采用较低的温度冻结及冻藏小龙虾,可以更好的保持冻藏小龙虾的品质。该试验可为小龙虾的冷冻保鲜及加工原料的周年供应提供理论指导。
