天然来源农药活性氨基酸的研究进展
2022-05-25吴兆圆柯少勇王开梅
吴兆圆,柯少勇,方 伟,王开梅
湖北省生物农药工程研究中心,武汉 430064
天然产物一直是药物和药物先导化合物的重要源泉[1]。根据2014年全球农药销售统计,在过去60多年里,基于天然产物的农药销售额约占总额的21%,如果将以天然产物为先导所开发的农药包括在内,该比例将达到53%[2],还有研究者认为,这一比例甚至达到70%[3]。
蛋白质是由氨基酸组成的具有生理活性多样性的高聚物,在生命活动中起着重要作用[4],而构成蛋白质的22种基本氨基酸(包括硒代半胱氨酸和吡咯赖氨酸)称之为蛋白氨基酸[5]。非蛋白氨基酸是指除蛋白氨基酸以外的含有氨基和羧基的化合物[6],自然界存在上千个非蛋白氨基酸,其中有250多个来源于植物,其它的来源于微生物、藻类及动物[7]。从这些天然氨基酸中发现了一些具有农药活性的化合物,许多农药品种的开发也将天然氨基酸作为构造单元[8],其中一种重要的氨基酸——膦丝菌素(phosphinothricin),已经开发用作除草剂[9]。因此,从自然界中寻找具有农药活性的氨基酸具有重要意义。本文主要总结近30年文献报道的具有农药活性的天然来源氨基酸,包括结构类型、来源、农药活性及部分作用机理,希望能为新农药的研究开发提供参考。
1 具有农药活性的天然蛋白氨基酸
除脯氨酸和高脯氨酸外,其它天然蛋白氨基酸均为α-氨基酸,且除甘氨酸外,它们的α-碳原子都为L-构型。列当(OrobancheandPhelipanchespp.)是一种世界范围内危害严重的根寄生杂草,室内试验表明天然甲硫氨酸(1,L-methionine)、赖氨酸(2,L-lysine)以及色氨酸(3,L-tryptophane)(见图1)能显著抑制分枝列当(O.ramosa)种子的萌发和早期发育,田间试验进一步确认了其抑制活性,且1的活性最强,抑制率可达67%[10],此外,2 mM的1可显著降低番茄根系上列当小块茎的数量[11]。天然亮氨酸(4,L-leucine)和酪氨酸(5,L-tyrosine)对另一种寄生杂草埃及独脚金(Strigahermonthica)具有植物毒性,盆栽试验表明二者能显著降低侵染玉米的埃及独脚金的生物量[12]。这些天然蛋白氨基酸是通过代谢途径的反馈抑制来抑制植物的生长[10]。

图1 具有农药活性的天然蛋白氨基酸
2 具有农药活性的天然非蛋白氨基酸
在动植物体内广泛存在的一些非蛋白氨基酸是它们的初级代谢中间产物或终产物,而绝大多数存在于植物和微生物体内的非蛋白氨基酸为它们的次级代谢产物[13],可以作为化学防御物质,对竞争者和捕食者产生毒害作用[14]。
天然非蛋白氨基酸不仅分布广泛,且结构多变,包括L型氨基酸和D型氨基酸,除α-氨基酸外,还有β-氨基酸、γ-氨基酸以及δ-氨基酸,除链状的脂肪族氨基酸外,还存在杂环氨基酸[4]。
2.1 具有农药活性的天然非蛋白α-氨基酸
大部分具有农药活性的非蛋白氨基酸为α-氨基酸,其结构类型包括脂肪族α-氨基酸、含环己二烯或苯环的α-氨基酸、含杂环的α-氨基酸以及α-亚氨基酸(见图2)。
2.1.1 脂肪族α-氨基酸及其农药活性
合欢氨酸(6,albizzin)(见图2)是从合欢(Albizziajulibrissin)的种子中得到,结构与谷氨酰胺类似,能抑制马铃薯、番茄晚疫病菌(Phytophthorainfestans)和小麦叶枯病菌(Zymoseptoriatritici)的生长[15]。6还具有化感作用,能抑制莴苣(Lactucasativa)种子的萌发和浮萍(Lemnaminor)的生长[16]。4-氯苏氨酸(7,4-chlorothreonine)由Yoshida等[17]从菌株Streptomycessp.OH-5093的代谢产物中分离得到,并发现该化合物能明显抑制萝卜和高粱幼苗的生长,其活性与双丙胺膦相当,除草机理是抑制氨基酸代谢,此外,7还表现出抗真菌活性,对白假丝酵母菌(Candidaalbicans)具有抑制作用。Yoshimura等[18]从高等真菌Amanitacastanopsidis中分离得到2-氨基-3-环丙基丁酸(8)和2-氨基-5-氯-4-戊烯酸(9),0.1 mM时均可抑制生菜幼苗根的生长,但不影响下胚轴的生长,其机制可能是降低细胞壁的机械延展性[19]。L-膦丝菌素(10,商品名为草铵膦)可由多种放线菌产生,具有广谱除草活性,同时也是苗前除草剂双丙氨膦(bialaphos)的N末端氨基酸,后者在植物中水解成10后才发挥除草作用[9,20],除草机理是抑制杂草体内谷氨酰胺合成酶的活性,造成氨的积累,从而抑制光合作用导致杂草死亡。δ-羟基正亮氨酸(11,δ-hydroxynorleucine)存在于菽麻(Crotalariajuncea)种子和根系分泌物中,具有一定的植物毒性,对生菜胚根的生长具有抑制作用[21]。4-甲酰氨基-L-乙烯基甘氨酸(12)由小麦根际细菌PseudomonasfluorescensWH6产生,能不可逆地抑制某些杂草种子的萌发,如一年生早熟禾(Poaannua),但对成熟植物的生长以及阔叶植物种子的萌发没有明显的影响[22]。此外,12还对梨火疫病菌(Erwiniaamylovora)等植物病原细菌具有抑菌活性[23]。L-2-氨基-4-甲氧基-反式-3-丁烯酸(13)由植物根际细菌铜绿假单胞菌P.aeruginosaPAO1产生,研究表明13与12具有类似的农药活性,能显著抑制梨火疫病菌,并对早熟禾种子的萌发具有弱的抑制作用[24]。L-刀豆氨酸(14,L-canavanine)广泛存在于豆科(Leguminosae)蝶形花亚科(Papilionoideae)植物及其种子中,与L-精氨酸相比较,L-刀豆氨酸δ位亚甲基上的碳原子被替换为氧原子,14可用于防治烟草天蛾(Manducasexta)、斜纹夜蛾(Spodopteralitura)以及亚洲玉米螟(Ostriniafurnacalis)[25]。14还具有抑制植物生长的作用,可抑制水稻幼苗第二叶鞘伸长[26],Hamako等[27]发现14具有化感作用,10 μM的14能显著抑制生菜原生质体的分裂。由植物内生菌P.syringaepv.syringae产生的3-甲基精氨酸(15,3-methylarginine)可显著抑制植物病原细菌P.syringaepv.glycinea的生长,其作用机理可能是抑制精氨酸的合成[28]。Ascaulitoxin(16)由杂草藜(Chenopodiumalbum)的致病真菌Ascochytacaulina产生,为双氨基酸N-葡萄糖苷,可以用作藜的生物防治。此外,采用叶片穿刺法,30 μg/滴的16可使甜菜(Betavulgaris)、苦苣菜(Sonchusoleraceus)、一年蓬(Erigeronannuus)、西方苍耳(Xanthiumoccidentale)、臭椿(Ailanthusglandulosa)等杂草以及某些栽培作物(如豌豆、黄瓜)的叶片形成明显的坏死斑,而在蕃茄及反枝苋(Amaranthusretroflexus)叶片上则形成较小的坏死斑[29]。从大黍(Panicummaxinmum)叶中得到的黍灰斑病菌(Phaeoramulariafusimaculans)产生malonofungin(17),是一种罕见的天然氨基丙二酸,具有广谱抗真菌活性,对灰霉菌(Botrytis)、稻瘟菌(Pyricularia)及镰刀菌(Fusarium)等植物病原真菌表现出抑菌活性[30]。

图2 具有农药活性的天然α-氨基酸
2.1.2 含环己二烯或苯环的α-氨基酸及其农药活性
Gao等[31]从一株未鉴定的放线菌SPRI-511356中分离得到了L-1,4-二环己烯基-1-丙氨酸(18),并报道了其除草活性,83.4 mg/L时对6种供试植物的地下部分的生长抑制率均在50%以上,而在此浓度下对地上部分生长的抑制相对要弱。细叶羊茅(FestucarubraL.ssp.commutata)根系分泌物中含有大量的L-间酪氨酸(19,L-meta-tyrosine),结构与苯丙氨酸类似,是一种强有效的化感物质[32],对超过50种杂草和作物的幼苗具有生长抑制作用[33],可作为广谱除草剂应用。L-3-(3,4-dihydroxyphenyl)-alanine(20,L-DOPA)是藜豆(Mucunapruriens)根分泌的化感物质,与19相比在苯环上多了一个取代羟基,能显著抑制生菜胚根的生长[34]。20还具有抗真菌活性,在500 μg/mL时对胶孢炭疽菌(Colletotrichumgloeosporioides)、辣椒炭疽菌(C.capsici)及茄腐镰刀菌(F.solani)等多种植物病原真菌具有抑制作用[35],此外,20对根结线虫(Meloidogyneincognita)及大豆胞囊线虫(Heterroderaglycines)具有较强的活性[36]。
2.1.3 含杂环的α-氨基酸及其农药活性
化合物21~25结构中含五元杂环,26和27含六元杂环,28~30含嘌呤基团。从甜菜叶面分离的假单胞菌株P.fluorescensSBW25产生呋喃霉素(21,furanomycin),研究表明其对软腐病菌(Dickeyadadantii)、丁香假单胞菌(P.syringae)及梨火疫病菌等植物病原细菌具有抑菌活性[37]。阿西维辛(22,acivicin)是由链霉菌S.sviceus产生,温室试验表明其对卵菌纲真菌具有广谱且良好的防控作用[38]。此外,22具有抑制谷氨酸合成酶的活性,已被申请作为除草剂进行专利保护[39]。TU-185(23)是由链霉菌185菌株产生的鸟氨酸衍生物,该化合物在琼脂平板上未表现出任何抗菌活性,但盆栽试验中在200 μg/mL时对黄瓜白粉病表现出一定的预防作用,防效达到86%[40]。豌豆幼苗根的提取物及分泌物中含有β-(3-isoxazolin-5-on-2-yl)-alanine(24),具有化感作用,能抑制杂草和莴苣根的生长,并使根尖坏死,但对豆类作物的根影响较小。此外,24还具有抗真菌活性,对Saccharomycescerevisiae最小抑制浓度为0.5 μg/mL[41],在50 μg/mL可显著抑制灰霉菌(B.cinerea)菌丝的生长,当浓度为250~500 μg/mL时对终极腐霉(Pythiumultimum)具有一定的抑制活性[42]。由天竺葵(Pelargoniumhortorum)产生的L-quisqualic acid(25)对日本金龟子(Popilliajaponica)具有神经毒性,研究表明25是一种罕见的兴奋性氨基酸激动剂受体,最终导致神经元的降解和神经细胞的死亡[43]。含羞草素(26,mimosine)来源于含羞草亚科(Mimosoideae)植物[44],具有抑制海藻糖酶、蔗糖转化酶及淀粉酶活性,从而阻碍昆虫生长发育所需要的能量供应[45],对白蚁工蚁的LD50为54.1 μg/虫,100 μM时对秀丽小杆线虫(Caenorhabditiselegans)致死率约为80%[46]。此外,研究表明26是银合欢属(Leucaena)植物产生化感作用的物质基础,对多种植物和杂草具有明显的生长抑制作用,100 mg/L的26对油菜(Brassicarapa)、菜豆(Phaseolusvulgaris)、鬼针草(Bidenspilosa)及黑麦草(Loliummultiflorum)的胚根生长的抑制率分别为94.8%、72.4%、45.2%和89.5%,且对下胚轴生长的抑制率分别为92.8、44.2、40.6和31.1%,且26对油菜的生长抑制作用可以被铁离子恢复,因此可作为开发生物除草剂的先导化合物[47]。Pyridazocidin(27)是由一株土壤链霉菌Streptomycessp.(#620061)产生的除草抗生素,对多种双子叶及单子叶杂草均具有很好的苗期除草活性,其中对大狗尾草(Setariafaberi)活性最强[48]。粘细菌Dictyosteliumdiscoideum产生的discadenine(28)的C-4位有一个嘌呤基团,与细胞分裂素结构相似,研究表明其在烟草愈伤组织上表现出细胞分裂素的活性,而全合成的(+)-discadenine也具有植物生长调节活性[49]。西尼霉素(29,sinefungin)是由链霉菌S.griseolus和S.incarnatusNRRL 8089产生,50~400 μg/mL时对白粉菌(Erysiphepolygoni)、大豆锈菌(Uromycesphaseolivar.typico)、炭疽病菌(C.lagenarium)、根癌农杆菌(Agrobacteriumtumefaciens)以及大豆细菌性叶枯病菌(Xanthomonasphaseolivar.sojensis)等多种植物真菌或细菌引起的病害具有很好的防治效果[50]。Vothknecht等[51]发现,29可以抑制镁-原卟啉IX甲基转移酶的活性,导致镁-原卟啉IX的积累,使得经29处理的大麦叶片中叶绿素的合成受到抑制。Micromonosporasp. A87-16806所产生的脱氢西尼菌素(30,dehydrosinefungin)对朱砂叶螨表现出良好的杀螨活性,被申请专利保护,用作杀螨剂[52]。
2.1.4α-亚氨基酸及其农药活性
α-氨基酸的氨基上一个氢被取代后形成α-亚氨基酸。从多花玉竹(Polygonatumodoratumvar.pluriflorum)中分离得到的氮代环丁烷基-2-羧酸(31)对多种淡水藻类具有抑制活性,其中对蓝藻(Anabaenaaffinis和Microcystisaeruginosa)显著抑制活性,对赤潮微藻(Cochlodiniumpolykrikoides)亦具有较强抑制活性,而对绿藻(Chlorellavulgaris和Scenedesmusspp.)的抑制活性较弱[53]。反式-4-氨基脯氨酸(32,trans-4-aminoproline)由壳二孢属真菌A.caulina产生,具有植物毒性,对寄主植物白藜叶片的毒性最高,对其它的双子叶植物的叶片也有毒性,但对单子叶植物叶片没有毒性[54]。冠菌素(33,coronatine)由丁香假单胞菌产生,与脱落酸、茉莉酸结构性质相似,是一种植物生长调节剂,具有促进细胞分化、提高叶绿素含量、调控植物生长、抑制细胞衰老等生理功能[55]。Nectriasp.DA060097所产生的cinnacidin(34)是异亮氨酸的衍生物,4 000 g/ha苗后处理对向日葵(Helianthusannuus)和大狗尾草具有良好的除草活性[56]。Abenquines A、B2、C和D(35~38)是从智利阿塔卡马沙漠高地分离的一株链霉菌中分离得到,结构中包含氨基酸和N-乙酰氨基苯醌,其在1~100 μM浓度下可以抑制蓝藻SynechococcuselongatusPCC6301的生长[57,58]。赭曲霉毒素A(39,ochratoxin A)可由炭黑曲霉(Aspergilluscarbonarius)产生,结构由L-苯丙氨酸和二氢异香豆素组成,对酱曲露尾甲(Carpophilushemipteru)和美洲棉铃虫(Helicoverpazea)具有很好的杀虫活性[59]。Giganticine(40)是从牛角瓜(Calotropisgigantea)的根中分离到的α-氨基-苯乙酸衍生物,对沙漠蝗虫(Schistocercagregaria)5龄若虫表现出良好的拒食活性,1.6 μg/cm2处理,拒食率达到了55%,随着浓度的升高,拒食率上升[60]。
2.2 具有农药活性的天然β-氨基酸及其农药活性
氨基变位酶可以催化胺基和氢在相邻碳上的可逆交换,微生物和植物中的β-氨基酸可以通过此酶的作用由α-氨基酸转化而来[61]。
许多植物体内都含有β-氨基丁酸(41,见图3),与植物的抗逆性相关,可以诱导植物对多种生物逆境及非生物逆境产生防御反应,对细菌、真菌及病毒病害产生抗性[62]。41还可影响相应作物上害虫,包括同翅目害虫,如豆蚜、桃蚜及甘蓝蚜等及鳞翅目害虫,如粉纹夜蛾及小菜蛾的生长发育[63,64]。α-亚甲基-β-丙氨酸(42)由Streptomycessp. A12701菌株产生,可抑制拟南芥的生长[65]。Yan等[66]发现水稻品种日本晴的种子、叶、根及根分泌物中都含有β-酪氨酸(43,β-tyrosine),在液体培养基中加入10 μM的43可以抑制丁香假单胞菌的生长,43还可抑制双子叶植物根的生长,其中对拟南芥根的生长抑制作用比较明显,IC50为4.4 μM。

图3 具有农药活性的天然β-氨基酸
2.3 具有农药活性的天然γ-氨基酸及其农药活性
顺式-2-氨基-1-羟基环丁烷-1-乙酸(44,见图4)由链霉菌S.rocheiA13018产生,可诱导植物褪色,10 μg/ml的该化合物处理拟南芥可导致中等程度褪色,更高浓度处理可导致严重的白化及生长的轻微抑制,以11.2 kg/ha的44进行芽后处理,可使牵牛花及芥菜产生明显褪色,其作用机理为干扰含硫氨基酸的合成或利用[67]。γ-氨基丁酸(45,GABA)广泛存在于各种植物中,其合成是通过谷氨酸脱羧酶,该化合物可影响相应作物上害虫的发育,研究表明45可以抑制蔷薇卷叶蛾(Choristoneurarosaceana)的生长,并降低其存活率[68,69]。45对害虫的作用机理是使氯离子通道受到抑制,而害虫对作物的取食可能会使作物内45快速积累[70]。45还可以通过促进光合作用及提升抗氧化酶的活性来提高小麦对盐的耐受性[71]。
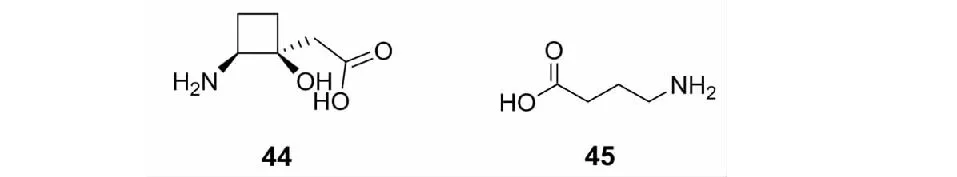
图4 具有农药活性的天然γ-氨基酸
2.4 具有农药活性的天然δ-氨基酸及其农药活性
5-氨基酮戊酸(46,5-ALA)(见图5)广泛存在于动物、植物、藻类及细菌中,是四氢吡咯的前体,双子叶植物比单子叶植物对该化合物更敏感,这种特性使其可作为除草剂开发[72]。46还可通过调节油菜幼苗内四氢吡咯及酪氨酸代谢来提高油菜对盐的耐受能力[73]。
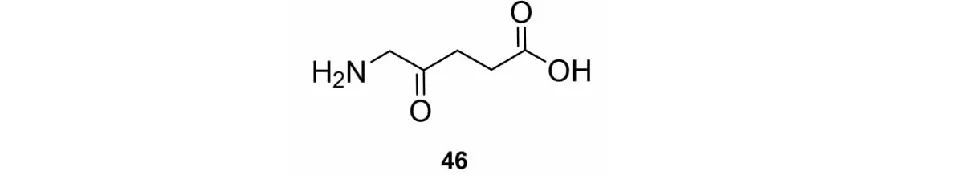
图5 具有农药活性的天然δ-氨基酸
2.5 其他其它具有农药活性的天然源氨基酸及其农药活性
甜菜等植物来源的甘氨酸甜菜碱(47,glycine betain)(见图6)对多种双子叶杂草,如欧活血丹(Glechomahederacea)、龙葵(Solanumnigrum)、苍耳(Xanthiumsibiricum)、反枝苋等具有芽前和苗后除草活性,此外,对马唐(Digitariasanguinalis)、偃麦草(Elymusrepens)等具有芽前除草活性,而且对这两种杂草三叶期前具有苗后除草活性,含有该化合物的制剂被申请专利[74]。

图6 其它具有农药活性的天然源氨基酸
3 结语
当前,由于人们对环境问题及食品安全问题的关注,高效安全及环保的农药新品种的开发越来越受到重视。许多天然来源氨基酸对杂草、有害真菌、细菌以及害虫具有较强的防治作用,有些已经被开发为商业化的农业产品,例如草铵膦是基于L-膦丝菌素(10)通过化学合成开发的[22],是近年来以微生物天然产物为先导开发的最成功的除草剂之一。由丁香假单胞产生的冠菌素(33)在低浓度下具有调控植物生长、提高叶绿素含量和植物抗逆性等生理功能,中国农业大学等单位已成功将其开发为植物生长调节剂。还有一些天然来源氨基酸在农业生产中具有良好的应用前景,如天然蛋白氨基酸甲硫氨酸(1)、赖氨酸(2)、色氨酸(3)、亮氨酸(4)以及酪氨酸(5)通代谢途径的反馈抑制产生除草作用,可以用于寄生杂草列当和埃及独脚金的生物防控。一些植物产生的非蛋白氨基酸可作为化感物质对其周边的竞争者产生抑制作用[75],如由豆科植物产生的合欢氨酸(6)、δ-羟基正亮氨酸(11)、L-刀豆氨酸(14)、L-DOPA(20)、β-(3-isoxazolin-5-on-2-yl)-alanine(24)、含羞草素(26)等均为氨基酸类化感物质,能选择性地抑制其它植物生长而对豆科和禾本科植物影响较小,因此可将其开发为除草剂应用于农业生产中。由细叶羊茅根分泌的间酪氨酸(19)也是一种强有效的化感物质,在μmol/L级浓度下对多种植物具有生长抑制作用,可作为广谱除草剂应用于农业生产。5-ALA(46)参与植物的生长调节过程,具有类似植物雌激素的生理活性,可作为植物生长调节剂在农业生产中使用。天然氨基酸来源丰富,具有广泛的农药活性,但对于其中的大多数化合物的作用机制有待于更深入的研究,以拓展其在农药领域中的应用。
