植物源生物活性成分控制肉及肉制品中酪胺含量的研究进展
2022-05-13包璐莹包天雨夏秀芳
包璐莹,石 硕,包天雨,夏秀芳
(东北农业大学食品学院,黑龙江 哈尔滨 150030)
酪胺(tyramine,Tyr)又称4-羟基苯乙胺,分子式为C8H11NO,相对分子质量为137.2,是一种低分子质量的单胺[1]。酪胺含量可反映食品中细菌指标或生物胺指数,可用来评估食品新鲜程度[2-3]。酪胺对人体的作用与其浓度相关,摄入低浓度(含量小于800 mg/kg)的酪胺不会对身体产生危害[4],而且具有增强呼吸作用[5]、使外周血管收缩[6]和作为神经递质[7]等生理功能,但摄入由于微生物作用或贮藏条件不当导致的含有高浓度酪胺的肉制品尤其是发酵肉制品或腌制、熏制的鱼肉[8],会引发高血压、偏头痛、呕吐、腹泻等疾病[9-10]。
鉴于酪胺对人的毒理作用,且酪胺具有热稳定性(85 ℃),无法通过热处理消除[11],人们利用物理法、化学法和生物法控制食品中酪胺含量。物理法主要包括气调/抗菌包装等。气调包装是通过改变食品包装中气体的组成及含量从而抑制产酪胺菌的生长,并达到控制酪胺含量的目的;抗菌包装是利用聚合物中具有抑菌性的金属或其氧化物纳米粒子释放到食品中,以抑制食品在贮藏期间产酪胺菌生长从而控制酪胺含量。Carbone等[12]将银纳米颗粒(AgNPs)与不可降解、可食用的聚合物结合制成活性食品包装,具有显著的抗菌性,但此方法不环保且无法确定该颗粒的毒性。化学法是指在产品贮藏、加工和销售期间添加化学防腐剂等以抑制酪胺形成,如添加5%山梨酸钾可有效抑制马鲛鱼在贮藏过程中酪胺的生成[13],但此方法在应用时会残留部分化学试剂。
消费者关于合成添加剂的担忧促进了生物法的应用,生物法是从动植物/微生物中提取天然且对人体无害的生物试剂从而控制酪胺含量,其中植物提取物如精油,作为由植物自身合成的可抑制细菌生长的次级代谢产物所组成的天然混合物,不仅参与植物抵抗外界伤害的防御机制[14],还可抑制肉及肉制品中产酪胺菌的生长并控制酪胺含量[15-16],被美国食品药品监督管理局(Food and Drug Administration,FDA)归类为公认的安全物质(generally recognized as safe,GRAS)[17],且抑制效果甚至优于合成添加剂[18],因成分天然、毒性及副作用小、生物可降解性好以及可与其他植物化学物质和非植物化学物质结合使用起到协同抑制作用等优点应用居多。
本文拟综述肉及肉制品中酪胺的形成机制、植物提取物活性成分的特性及在酪胺形成过程中的抑制作用,为提高肉及肉制品安全性及为植物提取物控制食品中酪胺含量提供参考。
1 肉及肉制品中酪胺形成途径
酪胺在肉及肉品中的形成有两条途径(图1)。两条途径都以酪氨酸为底物,但条件与底物细微的差别导致酪胺形成机制不同。第一条途径的条件一是原料肉固有的或肉制品贮藏过程中产生的产酪氨酸脱羧酶(tyrosine decarboxylase,TDC)的微生物,如粪肠球菌和屎肠球菌等[19];条件二是适宜这些微生物生长并产生TDC,且有利于TDC发挥作用的条件,如粪肠球菌在25 ℃、pH 5.5条件下,TDC活性最强;屎肠球菌在25 ℃、pH 6.0条件下,TDC活性最强[20]。第二条途径的底物除酪氨酸外还有脂质氧化产物如十七烷酸等,条件只需pH<7即可[21]。
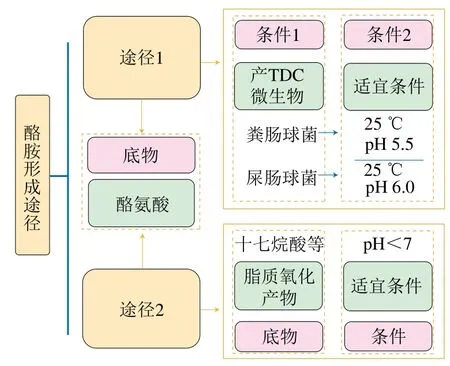
图1 肉及肉制品中酪胺形成途径[19-21]Fig. 1 Formation pathway of tyramine in meat and meat products[19-21]
两条途径中的底物酪氨酸分别通过酪氨酸脱羧作用或与羰基化合物发生羰基-氨基反应生成酪胺。第一条途径是酪胺形成的主要途径,在被氧化的脂质含量增多时,可能通过第二条途径促进酪胺形成[21-22],但该途径的具体反应机理有待进一步研究。图1中可看出底物和条件不同会引发不同的途径,从形成途径出发抑制肉及肉制品中酪胺形成可以从控制酪氨酸含量、抑制产TDC微生物生长、防止生成脂肪氧化产物3个角度考虑,而植物源活性成分具有的抑菌和抗氧化特性可以在酪胺形成途径中发挥抑制作用。
2 植物提取物活性成分及其结构
植物提取物活性成分的抑菌性和抗氧化活性在抑制肉品酪胺形成的过程中起着决定性作用,且各活性成分的结构对其特性有所影响,了解植物提取物活性成分的特性及其结构对植物提取物抑制酪胺形成具有重要意义。
2.1 抑菌活性成分及其结构
根据酪胺的形成途径及植物提取物活性成分的特性,植物提取物主要通过抑制包括肠杆菌(大肠杆菌)[23]、肠球菌(粪肠球菌)[17]以及不动杆菌[24]在内的产酪胺菌的生长来减少肉及肉制品中酪胺的形成,且植物提取物中的抑菌活性成分是决定植物提取物具有抑菌特性的重要因素[25]。植物提取物中抑菌活性成分多为酚类化合物,它是一类芳香环上有羟基取代基的化合物[26],按结构可分为酚酸类化合物、黄酮类化合物和挥发油等[27];按苯酚亚基数量可分为简单酚和多酚,简单酚由酚酸化合物组成,多酚可进一步细分为至少包含两个苯酚亚基的黄酮类化合物和具有3个及以上苯酚亚基的单宁[28-29]。
2.1.1 酚酸类化合物抑菌性及其结构
此类植物活性成分包括C6—C1型的没食子酸及其衍生物组成的羟基苯甲酸和C6—C3型的阿魏酸及其衍生物组成的羟基肉桂酸[30-31],羟基肉桂酸和羟基苯甲酸对大肠杆菌具有抑制作用,前者的侧链双键是负责其抑菌性的重要官能团,而对后者来说,苯环上—OH和—OCH3的数量对其抑菌活性具有一定影响,羟基苯甲酸苯环上—OH的数量与其抑菌活性呈负相关,且羟基苯甲酸中苯环上具有一个—OH和两个—OCH3的丁香酸抑制大肠杆菌的活性高于苯环上具有3个—OH的没食子酸的抑菌活性[32]。此外,有研究发现阿魏酸对粪肠球菌具有抑制作用[33],但其抑菌性与结构的构效关系有待进一步研究。
2.1.2 黄酮类化合物抑菌性及其结构
黄酮类化合物简称类黄酮[34],其基本碳架是由15个碳原子排列成C6-C3-C6 3个环(A、B和C环)的黄烷核[35],根据C环的饱和度不同及A环和B环上取代基不同可分为黄酮、异黄酮、黄酮醇等[36],其中黄酮醇化合物中的槲皮素可抑制产酪胺菌的生长[37]。红花黄酮提取物的主要活性成分为—OH数量较多的黄酮类化合物,具有广谱抑菌性,不仅可以抑制大肠杆菌的生长,还能抑制革兰氏阳性菌的生长[38]。有研究表明,虽然—OH数目增多会削弱黄酮类化合物的疏水性,但会增加C环上与电子特性相关的6 位碳原子的净电荷从而使其具有良好的抑菌活性[39]。
2.1.3 挥发油抑菌性及其结构
挥发油也称为精油[40],根据其化学结构可分为萜烯类化合物、萜类化合物、苯丙烷类化合物等。挥发油中能抑制酪胺形成的萜烯类化合物包括桧烯和蒎烯等[41];萜类化合物包括百里香酚和香芹酚等,它们由于结构中—OH基团的酸性和亲脂性而具有较强的抑菌活性[42]。百里香酚中单萜核第一个碳原子的—OH是负责其抑菌活性的官能团,有研究表明对该官能团以直链酯基进行结构修饰后,百里香酚衍生物对大肠杆菌的生长抑制作用与百里香酚相比有所降低[43];苯丙烷类化合物包括丁香酚和反式肉桂醛等[17]。反式肉桂醛中α,β-不饱和羰基部分即丙烯醛基团是负责其抑菌活性的官能团,以反式肉桂醛为主要成分的肉桂皮精油可抑制大肠杆菌的生长[15]。
石榴多酚和茶多酚可抑制产酪胺菌不动杆菌的生长,Zhuang Shuai等[44]研究发现以石榴多酚和鞣花酸等酚类化合物为主要活性成分的石榴皮提取物可有效抑制鳙鱼片在4 ℃贮存过程中不动杆菌的生长;茶多酚也具有抑制这种产酪胺菌生长的作用[45]。抑制产酪胺菌生长的植物提取物中活性成分的提取方法、结构及其在抑制酪胺含量过程中发挥作用的总结如表1所示。
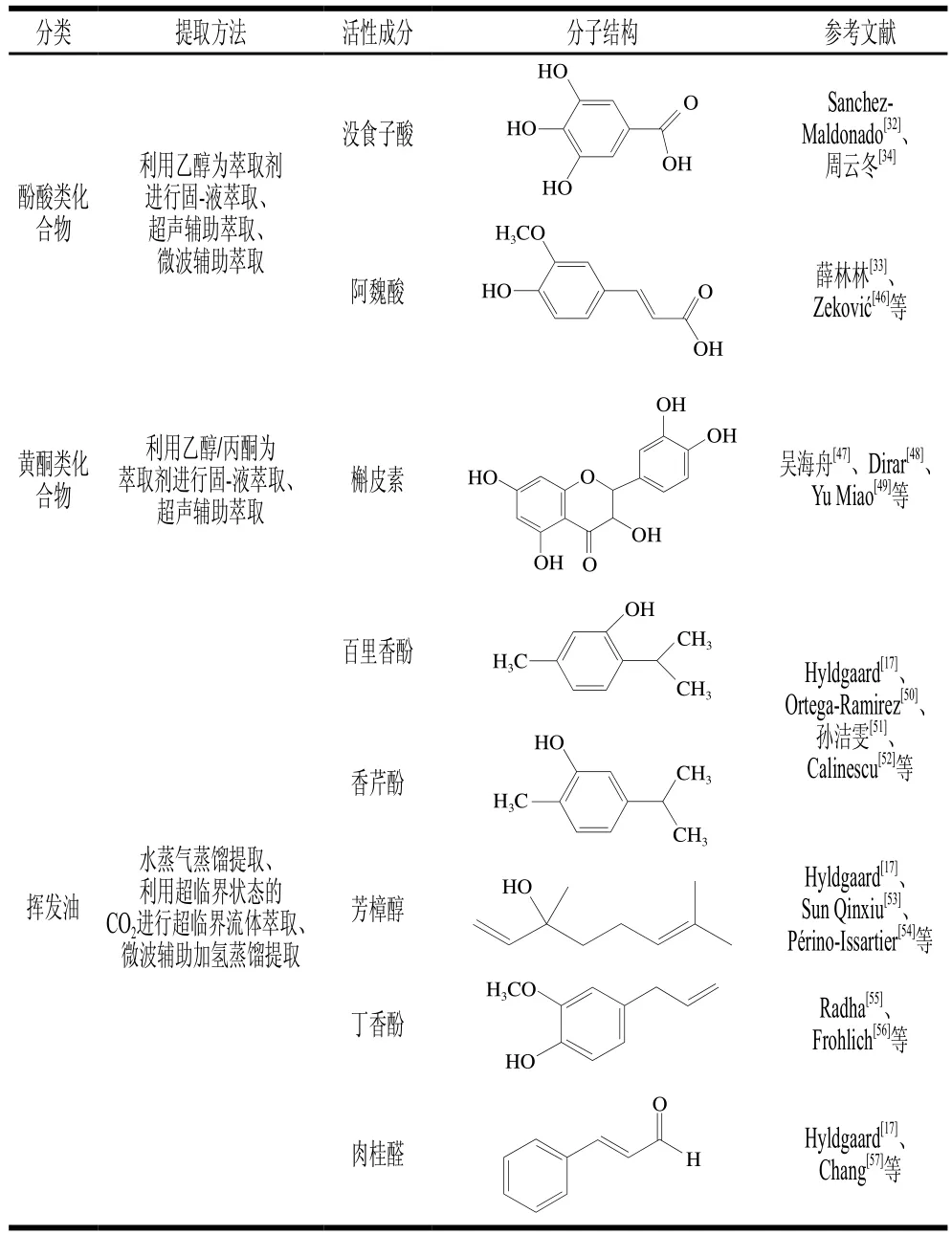
表1 抑制产酪胺菌生长的植物提取物活性成分的作用、提取方法及其结构Table 1 Functions, extraction methods and structures of active components from plant extracts
2.2 抗氧化活性成分及其结构
植物活性成分具有功能多样性,如酚酸类化合物与黄酮类化合物在肉及肉制品中不仅可作为抑菌剂,也具有抑制脂肪氧化的作用,且活性成分的结构对其抗氧化性具有一定影响。
2.2.1 酚酸类化合物抗氧化性及其结构
此类化合物的抗氧化活性取决于结构中—OH和甲氧基取代基的数量和位置。碳骨架为C6—C3型的咖啡酸可通过捕获自由基发挥抗氧化作用[58]。有研究表明没食子酸由于苯环上含3个邻位—OH,故具有较强的抗氧化性,咖啡酸苯环上的一个—OH被甲氧基取代后形成的阿魏酸的抗氧化活性更强[59];—OH数目多且具有邻-3,4-二羟基结构的原儿茶酸的抗氧化活性比没食子酸的抗氧化活性强[60]。
2.2.2 黄酮类化合物抗氧化性及其结构
此类化合物骨架上—OH的位置影响其抗氧化性,槲皮素因其结构含有3—OH、B环和C环而具有稳定共轭体系,且B环具有3’,4’-邻二羟基结构而具有抗氧化活性[61],根皮素中由A环的3个—OH和羰基构成的2,6-二羟基苯乙酮基团是赋予其抗氧化活性的基团[62]。茶儿茶素B环中的邻苯二酚基团、没食子酸酯基团、C环3位酯化的酯基与其抗氧化活性密不可分[63-64]。
3 活性成分控制酪胺含量形成的机制及抑制效果
植物提取物中含各种具有抑菌及抗氧化作用的活性成分,它们可能对不同作用靶标发挥不同作用,了解植物提取物及其所含活性成分的作用靶标以及作用机制可以更好地将植物提取物应用于控制肉及肉品中酪胺的形成。
3.1 活性成分的抑菌机制
3.1.1 酚酸类化合物的抑菌机制
在产酪胺菌如肠球菌的TDC基因簇中,与酪胺合成密切相关的酪氨酸脱羧酶基因(tyrosine decarboxylase,tyrDC)、控制细胞膜内外酪氨酸与酪胺交换的酪氨酸/酪胺透性酶基因(tyrosine/tyramine permease,tyrP)[65]、酪氨酰-tRNA合成酶基因(tyrosyl-tRNA synthetase,tyrS)和nhaC基因共同调控产酪胺菌生物合成酪胺,其中tyrS基因表达量会因酪氨酸含量的减少而提高,该过程是阻止酪氨酸大量脱羧的负调节系统[33]。tyrP基因从膜外吸收酪氨酸,再将生成的酪胺排放到膜外同时吸收膜外的酪氨酸[66]。
这些基因的表达量会受到植物提取物中某种活性成分如阿魏酸的影响而降低或升高。有研究表明,培养基内不含酪氨酸底物时,tyrS基因的表达量会被酚酸类化合物阿魏酸所促进,而含有酪氨酸底物时,tyrS基因的表达受阿魏酸的影响较小,但TDC和Na+/H+转运蛋白受到阿魏酸的影响而对酪氨酸的感应能力减弱,导致tyrDC和tyrP基因的表达明显受到抑制[33]。
3.1.2 黄酮类化合物的抑菌机制
脂质和蛋白质是组成细胞膜的两个主要成分,细胞膜基本特性之一的细胞膜流动性就是指这两种主要成分的运动性。细胞膜流动性在维系细菌正常功能(细胞分裂、扩散等)中具有重要作用,它会随着膜脂成分(脂肪酸结构和饱和度、脂肪酸链长度以及顺式脂肪酸与反式脂肪酸的比例)的变化而发生变化[34],若细胞膜不饱和脂肪酸含量减少,会降低膜流动性,从而导致细胞死亡。如黄酮类化合物柚皮素可以通过改变大肠杆菌细胞膜内不饱和脂肪酸比例而改变膜的流动性[39]。
3.1.3 挥发油的抑菌机制
精油的抑菌作用机理如图2所示,主要包括以下3种方式:1)精油活性成分可通过减少脂多糖含量抑制产酪胺菌生物膜形成;2)可在细菌细胞膜内积累并破坏其完整性,使胞内物质如核酸、Na+、K+、Ca+流出;3)进入细胞内抑制关键酶活性。此外,精油中多酚类活性成分还可抑制产酪胺菌鞭毛介导的运动以阻止生物膜的形成。具体抑菌机制表述如下。
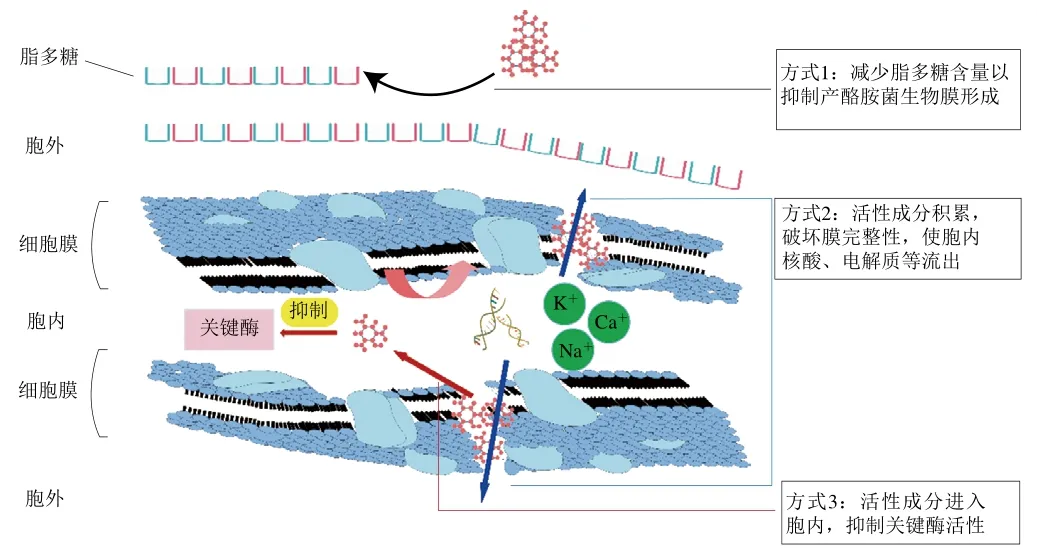
图2 精油抑菌作用机理[67-75]Fig. 2 Antibacterial mechanism of essential oil[67-75]
3.1.3.1 抑制产酪胺菌生物膜形成
从广义上生物膜定义为由细胞分泌的胞外聚合基质所包裹的一种表面附着细菌群落,这是细菌的自然状态和主要状态[67-68]。挥发油中活性成分可以通过影响生物膜形成过程中的决定性因素,即细胞外聚合物的合成来抑制产酪胺菌生物膜的形成[67]。研究发现,粪肠球菌经128 μg/mL香芹酚处理后培养12 h,随着培养时间的延长,处理组生物膜胞外多糖含量均显著低于对照组,说明香芹酚可以通过抑制甚至减少维持生物膜三维结构不可或缺的胞外多糖的合成或分泌从而抑制生物膜的形成[69]。
精油活性成分对菌体聚集后生物膜成熟期的微观形态也有影响,靳盼盼[69]通过扫描电子显微镜和激光共聚焦显微镜观察了粪肠球菌的生物膜成熟度对膜内细菌的影响,发现处理组的粪肠球菌生物膜与同时期对照组相比个体细胞无明显形态变化,但细胞之间的连接不致密,细胞层数较薄。
3.1.3.2 改变产酪胺菌细胞膜通透性
精油可通过改变产酪胺菌细胞膜通透性、破坏其完整性从而使胞内物质如蛋白质、核酸和电解质流出,进而抑制产酪胺菌生长[70-71]。精油对产酪胺菌细胞膜的破坏作用与其复杂的化学成分密不可分,其中具有代表性的化学成分包括百里香酚(百里香精油的有效成分)[72]、香芹酚(牛至精油的有效成分)[50]和丁香酚(罗勒精油的有效成分)[73]。
百里香酚是一种单萜[17],具有亲脂性和水溶性,可以迁移穿过胞外水性介质,到达、渗透并保留在细菌细胞质膜中,通过扰动质膜脂质组分引起细胞膜渗透性改变和胞内物质外泄发挥抑菌作用[74-75]。香芹酚的抑菌作用方式是增加细胞膜渗透性或作用于细胞质膜时使离子在膜上发生被动转运,前者是其主要作用方式。丁香酚能渗透细胞膜并与蛋白质相互作用,在致死浓度下破坏细胞质膜,从而增加其非特异通透性以达到抑菌的目的[17]。
3.1.3.3 降低产酪胺菌内重要酶的活性
产酪胺菌如大肠杆菌中与生长及复制密切相关的酶包括ATP酶和DNA拓扑异构酶。前者酶活性降低是导致细菌死亡的主要因素之一[76],而后者是通过催化DNA链断裂和拓扑变化从而影响DNA代谢的酶,拓扑异构酶I是一种普遍存在于细菌细胞内且不依赖ATP的单体酶,在DNA复制中起主要作用[71]。Cui Haiying等[71]研究发现用滁州菊花精油处理后的大肠杆菌,其ATP酶活力较对照组下降了86.34%,且通过DNA拓扑异构酶活性抑制实验发现该精油也能抑制在DNA复制中起主要作用的拓扑异构酶I的活性。
3.1.3.4 抑制产酪胺菌的丛动运动
在生物膜形成早期,由细菌鞭毛所介导的运动在初始附着阶段十分重要[68],产酪胺菌大肠杆菌鞭毛所驱动的运动包括泳动和丛动两种类型。前者代表单一菌体运动,后者代表群体菌体运动[69]。细菌的丛动能力决定了生物膜形态,丛动能力强的菌株能形成平坦、均匀的生物膜,而丛动能力弱的菌株会形成含有细胞聚集物的生物膜[77]。玫瑰多酚可通过减弱大肠杆菌的丛动能力进而抑制其生物膜的形成[78]。
3.2 活性成分的抗氧化机制
脂肪氧化是由引发、增殖和终止3个阶段组成的自由基链式反应,肉及肉制品中脂质氢过氧化物的形成、蛋白质尤其是包括肌红蛋白和血红蛋白在内的血红素蛋白氧化所释放的金属离子会促进脂肪氧化。植物提取物的活性成分可通过与自由基反应以中断氧化链式反应并清除自由基、阻止氢过氧化物的形成以及螯合金属离子来发挥抗氧化作用。
3.2.1 酚酸类化合物的抗氧化机制
酚酸类化合物的抗氧化机制包括清除自由基和抑制血红素蛋白氧化诱导的脂质氧化两个方面,前者反应机理主要为氢原子转移(hydrogen atom transfer,HAT),即通过代替脂质被自由基夺取氢原子发挥对脂质的保护作用[79]。但也有研究表明当酚酸化合物处于可电离状态时,其清除1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基的反应机理是HAT和连续的质子损失电子转移同时进行[80]。代表性化合物咖啡酸具有较高的自由基清除活性,有研究表明咖啡酸可清除DPPH自由基和羟自由基[58]。
肌红蛋白氧化后生成的高铁肌红蛋白在脂质过氧化物存在下会生成促进脂肪氧化的强氧化剂铁酰肌红蛋白[81],有文献指出阻止该过程的有效方法是抑制脂质氢过氧化物的形成[82],也有研究表明咖啡酸和没食子酸可有效抑制碎肉在贮存过程中氢过氧化物的形成,并相应地抑制了肌红蛋白诱导的脂质氧化[83]。
3.2.2 黄酮类化合物的抗氧化机制
此类化合物通过自由基清除机制和金属螯合抑制脂肪氧化,其中最具代表性的物质为槲皮素、根皮素和茶儿茶素。肌红蛋白自氧化过程中会破坏卟啉环结构从而释放出游离二价铁离子,该离子可通过Fenton反应促进脂肪氧化[84-85],槲皮素可与二价铁离子形成物质的量比为3∶2和2∶1的两种络合物以减少铁离子数量[86]。血红蛋白自氧化后生成的高铁血红蛋白更易释放含三价铁离子的卟啉环Hemin,Hemin可高效降解脂肪氧化引发阶段的产物氢过氧化物从而继续延伸脂肪氧化链反应[87],槲皮素可与高铁血红蛋白形成与Hemin亲和力更强的加合物。此外,槲皮素还具有羟自由基清除活性[47]。
根皮素可通过清除氧自由基发挥抗氧化作用[88],由于其药效基团的互变异构作用,可在氢原子被夺取后形成稳定的自由基,具体反应机理见图3。2,6-二羟基苯乙酮苯环上—OH之一的氢被夺取后一方面导致了羰基的酮-烯醇转化,另一方面使羰基的α-氢原子向氧自由基转移,同时未成对电子转移到羰基上,进一步的氢转移将未配对电子转移到另一个芳环—OH上,再经氢原子转移从而形成稳定的自由基[62]。
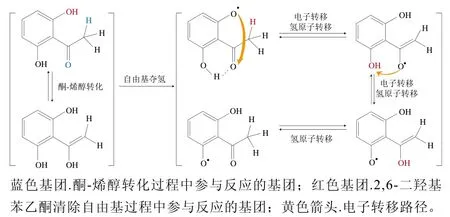
图3 2,6-二羟基苯乙酮清除自由基反应[62]Fig. 3 Free radical scavenging by 2,6-dihydroxyacetophenone[62]
茶儿茶素能成为茶多酚的主要抗氧化活性成分与其含有表没食子儿茶素没食子酸酯(epigallocatechin gallate,EGCG)密不可分[89],其抗氧化机制为B环的邻二羟基基团与自由基反应,生成不活泼的半醌从而清除自由基[64]。
3.3 抑制酪氨酸形成的效果
不同的活性成分抑制肉及肉品中酪胺形成的效果不同,了解各活性成分的抑制效果对降低各类肉及肉制品酪胺含量具有重要意义。
3.3.1 酚酸类化合物的抑制效果
酚酸类化合物阿魏酸可抑制粪肠球菌TDC基因簇中基因的表达,有研究表明,空白对照组粪肠球菌中tyrDC基因及tyrP基因表达量分别是阿魏酸添加组的7.6 倍和10.3 倍,阿魏酸的添加使酪胺产量降低了27%[69]。
另一种酚酸化合物咖啡酸,抗氧化性较强,浓度为1 mmol/L时的DPPH自由基和过氧化氢自由基清除活性分别可达到84.7%和99.8%[58],但其抗氧化活性对抑制肉及肉制品中酪胺含量的作用还有待进一步研究。
3.3.2 黄酮类化合物的抑制效果
可控制肉及肉制品中酪胺含量的代表性黄酮类化合物是根皮素,主要通过抑制产酪胺菌的生长以达到控制目的,有研究表明将鲑鱼浸入质量浓度4 mg/mL的根皮素浸泡液中,在4 ℃下保存3 d后,处理组中产酪胺菌不动杆菌的相对丰度下降了1.06%,且处理组酪胺含量较对照组酪胺含量低17.12 mg/kg,并由此得到根皮素抑制酪胺含量是由于抑制了产酪胺菌生长的结论[90]。
根皮素也具有抗氧化活性,可使浸泡于4 mg/mL根皮素溶液中的鲑鱼片在贮存过程中的MDA含量减少2.9 g/kg[90]。黄酮类化合物能否通过抑制酪胺形成的补充途径达到控制酪胺含量的目的还有待进一步研究。
3.3.3 挥发油的抑制效果
精油会对产酪胺菌造成不可逆损伤,同时损伤程度具有浓度依赖性。Guo Na等[70]发现大肠杆菌经质量浓度3.13 mg/mL的砂仁精油处理2 h后形态发生改变,细胞边界起皱且不规则,悬浮液中核酸和蛋白质浓度分别增加了6.2 倍和10.8 倍;当精油质量浓度升至6.25 mg/mL时,细胞被破坏,裂解成碎片,处理2 h后悬浮液中核酸和蛋白质浓度分别增加了9.1 倍和20.5 倍。砂仁精油也会使大肠杆菌胞内电解质如K+、Ca2+和Na+等泄漏[91]。
由芳香/刺激性挥发油、有机酸和辛辣成分组成的香辛料提取物可抑制肉制品中产酪胺菌的生长并减少酪胺含量。Sun Qinxiu等[53]研究发现以丁香酚为主要成分的丁香醇提取物能够抑制哈尔滨干香肠中肠杆菌的生长,使发酵过程中干香肠内的肠杆菌细菌数降至1.57(lg(CFU/g))。该植物提取物还可阻止干发酵羊肉香肠在发酵过程中酪胺的积累,处理组酪胺浓度与对照组酪胺浓度相比降低了18.7%[41]。
精油通过抑菌从而抑制肉及肉制品中酪胺含量,同时对肉及肉制品的感官特性具有积极影响。有研究表明留兰香精油处理后的红鼓鱼片在4 ℃贮藏20 d后,酪胺含量仅为0.06 mg/kg,且与丁香精油和小茴香精油处理的鱼片相比感官得分更高[16]。以百里香酚和香芹酚为主要成分的百里香精油和牛至精油可使真空包装鲤鱼鱼片在3.5 ℃保存21 d时,酪胺含量较对照组分别减少了90.6%和94.0%,且能有效抑制鲤鱼鱼片在贮藏过程中感官品质的劣变[72]。
茶多酚主要通过抑制诸如不动杆菌和粪肠球菌的产酪胺菌的生长控制酪胺含量。Jia Shiliang等[45]研究发现鳙鱼鱼片浸没在质量浓度5 mg/mL和10 mg/mL的茶多酚溶液中,在4 ℃贮藏6 d后其不动杆菌的相对丰度分别下降至4.7%和6.9%,酪胺含量与对照组酪胺含量相比分别减少了15.8%和24.7%;Lu Shiling等[92]研究发现将茶多酚质量分数为0.19%的植物提取物加入熏制马肉香肠中可抑制粪肠球菌的生长甚至致其死亡,在香肠贮藏第28天时,处理组中酪胺浓度较对照组减少了23.8%。
4 结 语
植物提取物可通过抑制产酪胺菌生长来影响酪胺形成的主要途径以控制肉及肉制品中酪胺含量,酚类化合物是最具代表性的抑菌活性成分,通过影响产酪胺基因表达、破坏产酪胺菌细胞膜完整性致使胞内内容物流出以致细菌死亡、降低产酪胺菌内关键酶活性和抑制产酪胺菌生物膜的形成以发挥抑菌作用,但对肉及肉制品中主要产酪胺菌粪肠球菌细胞膜及其形态变化的影响研究较少。植物提取物也具有通过氢原子转移机制清除自由基、螯合金属离子以及抑制脂质氢过氧化物形成以发挥抗氧化作用的抗氧化活性成分且酪胺形成的补充途径与脂肪氧化产物增多有关,植物提取物是否能通过影响酪胺形成的补充途径从而控制酪胺含量是未来的研究方向。
