2012—2018年长春市手足口病非肠道病毒A组71型肠道病毒V P1基因特征分析
2022-05-13张智丽李晨光张兴国孙宇崔薇乔凤娟孙炳欣
张智丽,李晨光,张兴国,孙宇,崔薇,乔凤娟,孙炳欣
长春市疾病预防控制中心,吉林长春 130033
手足口病(hand,foot and mouth disease,HFMD)是一种由多种肠道病毒引起的、我国法定报告管理的丙类传染病[1],主要通过粪-口途径传播,也可通过呼吸道和接触传播,5岁以下婴幼儿为主要易感人群,好发于夏秋季[2]。自从我国将HFMD列入丙类传染病管理后,长春市HFMD病例报告始终位于丙类传染病前列[3]。2008—2012年,长春地区HFMD优势病原为肠道病毒71型(enteroviruses 71,EV-71)[4],而近年来病原构成逐渐发生变化,EV-71占比逐渐下降,柯萨奇病毒A组16型(coxsackievirus A16,CV-A16)及其他肠道病毒比重逐渐增加[5-6]。HFMD至今仍无有效疫苗及特异性治疗手段。因此,加强疾病监测,准确处置疫情,开展健康教育是控制该病流行的关键[7]。本研究分析了2012—2018年长春市HFMD病例的病原学特征,为今后开展本地区传染病的预防控制工作提供科学依据。
1 材料与方法
1.1样本 收集2012—2018年长春市哨点医院和长春地区各县(市)区疾控中心采集的HFMD病例的肛拭子、粪便、咽拭子样本(获得长春市疾病预防控制中心伦理委员会批准)。严格按照卫生部印发的《手足口病预防控制指南(2009版)》[8]要求采集、运输、保存。纳入标准:①临床诊断病例。在流行季节发病,常见于学龄前儿童,婴幼儿多见,发热伴手、足、口、臀部皮疹,部分病例可无发热;极少数重症病例皮疹不典型,临床诊断困难,需结合病原学或血清学检查进行诊断。②确诊病例。临床诊断病例具有下列之一者即可确诊:肠道病毒(CV-A16、EV-71等)特异性核酸检测阳性;分离出肠道病毒,并鉴定为CV-A16、EV-71或其他可引起HFMD的肠道病毒;急性期及恢复期血清CV-A16、EV-71或其他可引起HFMD的肠道病毒中和抗体有4倍以上升高。
1.2主要试剂及仪器 全自动核酸提取仪、肠道病毒荧光定量PCR检测试剂盒购自江苏硕世生物科技股份有限公司;RT-PCR试剂盒(210212)、毛细管电泳仪(卡夹QIAxcel,DNA Screening Cartridge)购自德国凯杰(QIAGEN)公司。
1.3病毒核酸提取及PCR检测 采用全自动核酸提取仪提取样本核酸后,再利用肠道病毒荧光定量PCR检测试剂盒对样本中总肠道病毒、肠道病毒A组71型(EV-A71)和CV-A16进行荧光定量PCR。其中非EV-A71阳性样本VP1区利用引物[CV-A6-VP1-F:5′-TGTTGGGCACG(A)CACGTCTGGGA-3′,CV-A6-VP1-R:5′-CCAGCATAATTTGGGTTGGCT(C)TTG-3′;CV-A16-VP1-F:5′-CAGTAATACACACTAC(T)AGGGCA-3′,CV-A16-VP1-R:5′-ACCCTATAGTTGCCT(C)ACATATA-3′;CV-A10-VP1-F:5′-ATGGCDACAGGCAAGATGCT-3′,CV-A10-VP1-R:5′-ATAYCTAGCAGGGTAATACTC-3′。引物由生工生物工程(上海)股份有限公司合成]进行RT-PCR扩增,扩增片段大小为1 000 bp。反应条件为:55℃30 min;94℃2 min;94℃15 s,55℃30 s,68℃40 s,共35个循环;72℃10 min。PCR产物用毛细管电泳仪(卡夹QIAxcel,DNA Screening Cartridge)进行检测,并送生工生物工程(上海)股份有限公司测序。
1.4生物信息学分析 用LaserGene软件包中Mega-Align软件对序列进行比对,截取VP1区进行后续分析。用于构建系统发生树的非EV-A71各基因型代表株的参比序列均来自美国国立生物技术信息中心GenBank(http://www.ncbi.nlm.nih.gov/)。采用NJ(neighbor-joiningmethod)法构建系统进化树,利用MEGA X软件,设置bootstrap值为1 000。
2 结果
2.1VP1区基因进化分析
2012—2018年长春市2 621例核酸检测阳性病例中,非EV-A71阳性样本VP1区PCR产物测序结果显示,共获得83条CV-A16序列、30条CV-A6序列和10条CV-A10序列。
2.1.1CV-A16分离株VP1区基因进化分析 基于VP1区(891 bp),对CV-A16序列及其各基因型和亚型参考株序列进行进化分析,结果显示,2012—2018年长春市HFMD病例中CV-A16毒株相对集中,83条序列均属于B1亚型,其中8条属于B1a亚型(2012年1条,2018年7条),其他均属于B1b亚型。见图1。
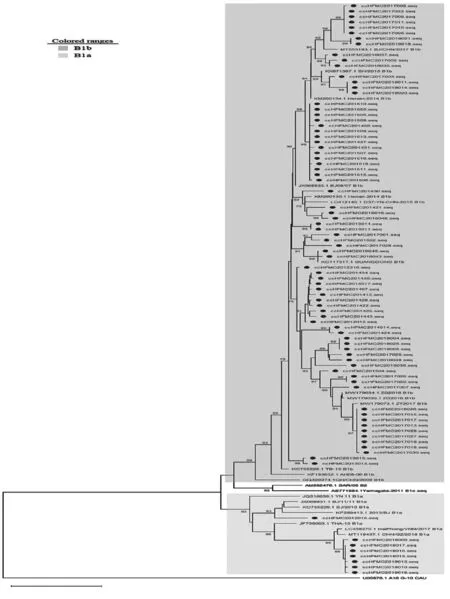
图1 2012—2018年长春市HFMD病例中CV-A16型VP1蛋白系统进化树Fig.1 Phylogenetic tree of VP1 protein of CV-A16 in Changchun City from 2012 to 2018
2.1.2CV-A6分离株VP1区基因进化分析 基于VP1区(915 bp),对CV-A6序列及其各基因型和亚型参考株序列进行进化分析,结果显示,2012—2018年长春市HFMD病例中CV-A6毒株相对集中,30条序列均属于D3亚型。见图2。
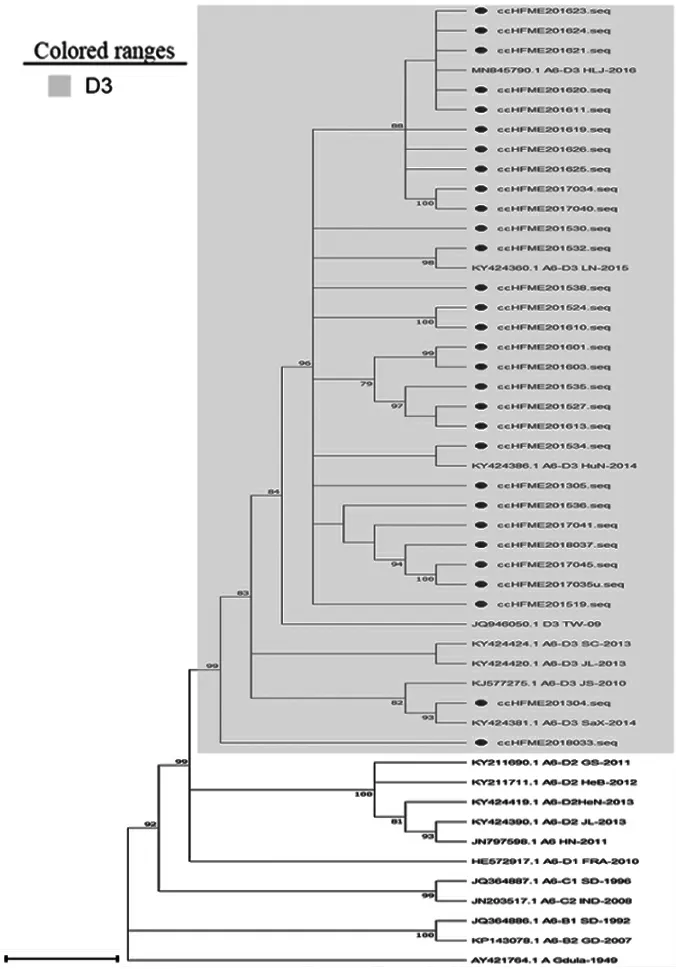
图2 2012—2018年长春市HFMD病例中CV-A6型VP1蛋白系统进化树Fig.2 Phylogenetic tree of VP1 protein of CV-A6 in Changchun City from 2012 to 2018
2.1.3CV-A10分离株VP1区基因进化分析 基于VP1区(894 bp),对CV-A10序列及其各基因型和亚型参考株序列进行进化分析,结果显示,2012—2018年长春市HFMD病例中CV-A10毒株相对集中,10条序列均属于D2亚型。见图3。
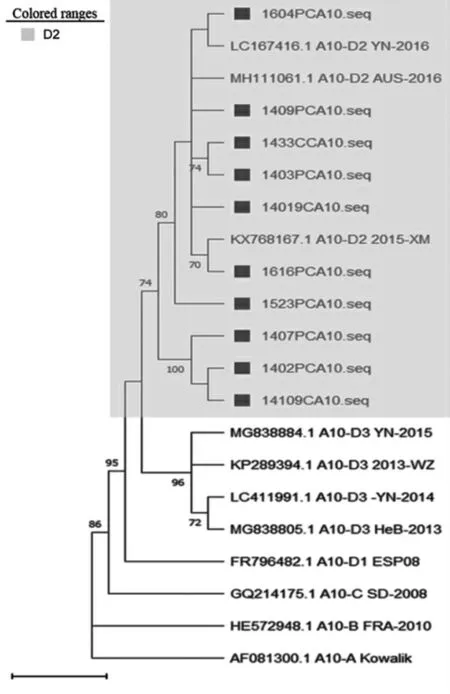
图3 2012—2018年长春市HFMD病例中CV-A10型VP1蛋白系统进化树Fig.3 Phylogenetic tree of VP1 protein of CV-A10 in Changchun City from 2012 to 2018
2.2VP1区核苷酸序列同源性分析
2.2.1CV-A16流行株VP1区同源性分析 结果显示,CV-A16各流行株与CV-A16原型株的同源性为75.0%~77.0%,B1a亚型分支与B1a亚型参考株的同源性为92.3%~98.5%,B1b亚型分支与B1b亚型参考株的同源性为94.5%~99.2%,与B1c亚型同源性为88.4%~91.7%,与B2亚型同源性为89.3%~93.0%,其中与B1b亚型同源性较高,各株间同源性为88.2%~100.0%。见图4。

图4 2012—2018年长春市HFMD病例中CV-A16型基于VP1区序列的同源性分析Fig.4 Homology of CV-A16 based on sequence of VP1 region in HFMD cases in Changchun City from 2012 to 2018
2.2.2CV-A6流行株VP1区同源性分析 结果显示,CV-A6各流行株与CV-A6原型株的同源性为82.7%~83.5%,与B1亚型参考株的同源性为81.9%~93.7%,与B2亚型参考株的同源性为82.1%~84.5%,与C1亚型参考株的同源性为85.1%~86.5%,与C2亚型参考株的同源性为83.8%~84.8%,与D1亚型参考株的同源性为89.7%~91.1%,与D2亚型参考株的同源性为89.2%~91.3%,与D3亚型参考株的同源性为92.8%~98.1%,其中与D3亚型同源性较高,各株间同源性为91.6%~99.8%。见图5。
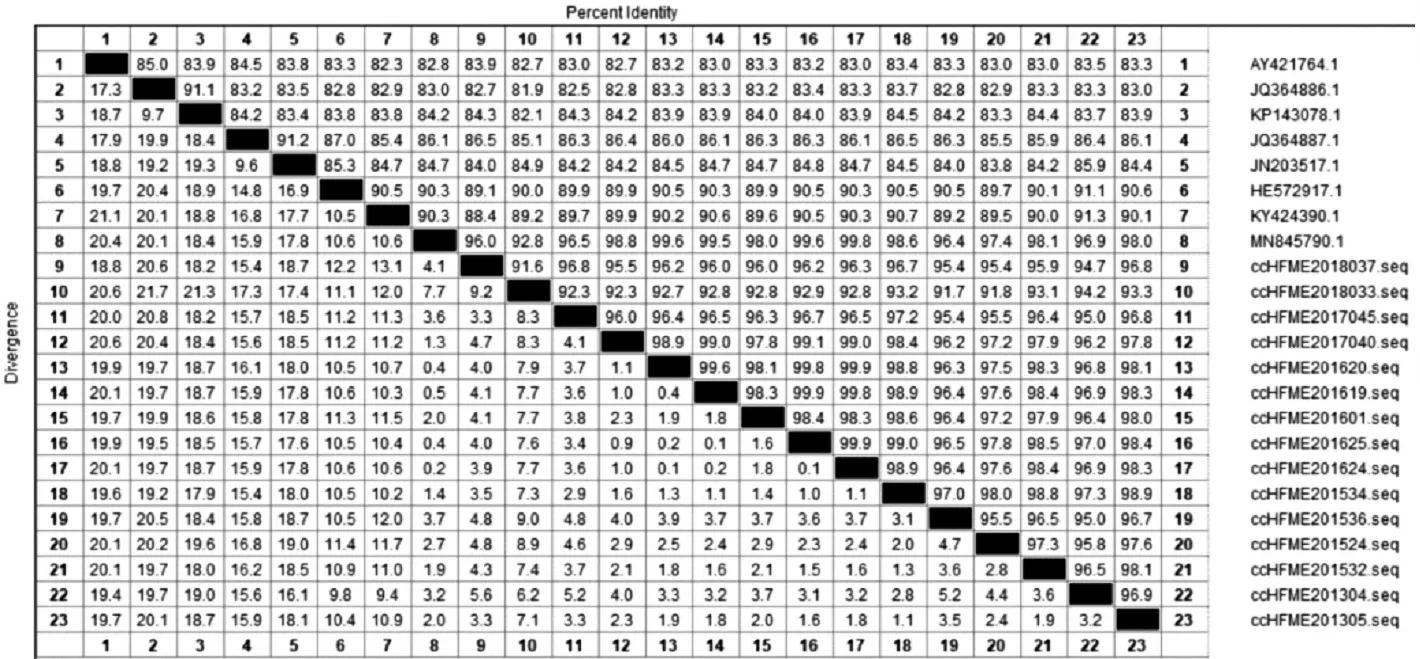
图5 2012—2018年长春市HFMD病例中CV-A6型基于VP1区序列的同源性分析Fig.5 Homology of CV-A6 based on sequence of VP1 region in HFMD cases in Changchun City from 2012 to 2018
2.2.3CV-A10流行株VP1区同源性分析 结果显示,CV-A10各流行株与CV-A10原型株的同源性为76.3%~77.0%,与B亚型参考株的同源性为78.9%~80.3%,与C亚型参考株的同源性为82.9%~83.8%,与D1亚型参考株的同源性为94.1%~95.4%,与D2亚型参考株得同源性为96.4%~98.4%,与D3亚型参考株的同源性为94.6%~96.1%,其中与D2亚型同源性较高,各株间同源性为96.4%~100.0%。见图6。
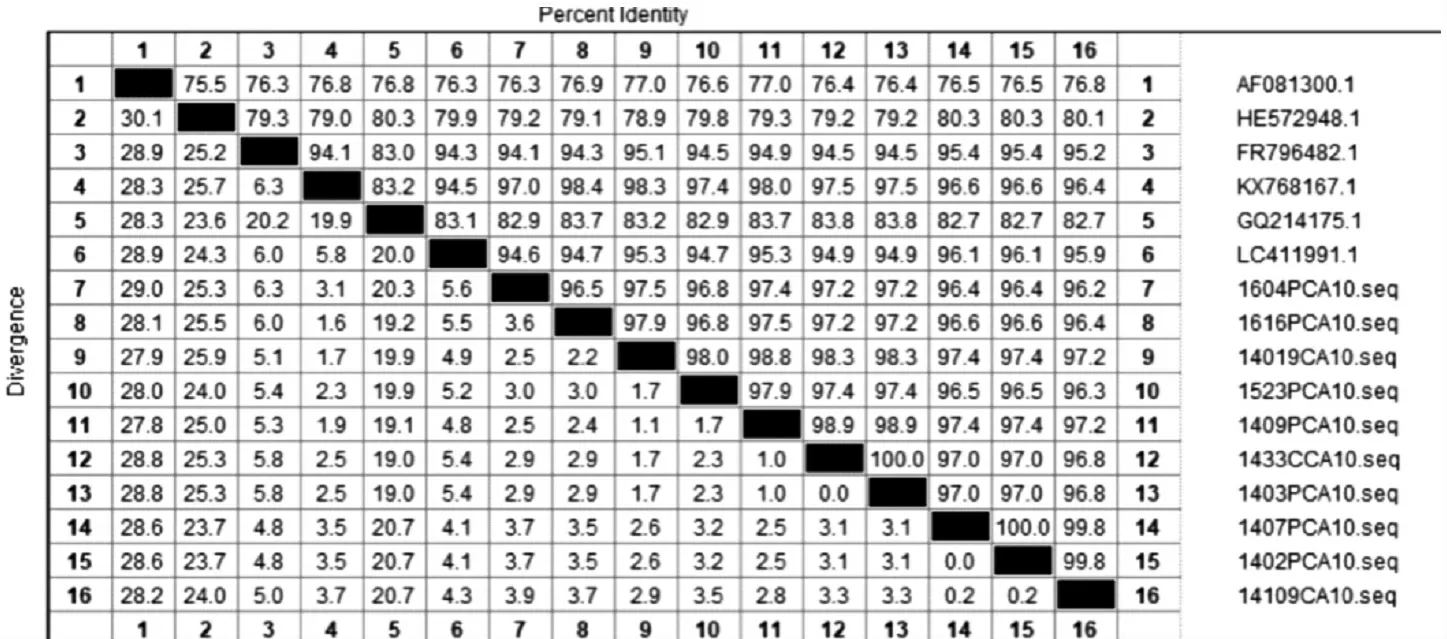
图6 2012—2018年长春市HFMD病例中CV-A10型基于VP1区序列的同源性分析Fig.6 Homology of CV-A10 based on sequence of VP1 region in HFMD cases in Changchun City from 2012 to 2018
2.3CV-A16流行株变异位点分析 CV-A6和CVA10流行株突变位点并未发现明显代表性,因此,选取CV-A16流行株进行变异位点分析。将所获序列中的代表序列经MEGA X软件翻译获得氨基酸序列后进行变异位点分析,以病毒流行株JX068833.1 BJ08/07 B1b为参考株,结果显示,CV-A16毒株中存在14个氨基酸位点变异,有12株在第14位点上发生N天冬酰胺→S丝氨酸(N14S)的变异,其中9株同时存在L215P变异,1株(ccHFMC2017030)同时存在N14S、L215P和F131S变异,另有1株(cc-HFMC-2017007)存在N14S、M23L和T94A 3个位点变异。有9株存在M23L变异,其中8株为B1a亚型,这8株除ccHFMC2012015外,均同时存在T164K和V251I变异,ccHFMC2012015则同时存在N218S变异。有2株同时存在T11A变异,其中ccHFMC2-016046同时存在I262V变异。在ccHFMC2017008、ccHFMC201430、ccHFMC201421这3株中则分别可见A22T、I235V和T289A变异。见表1。
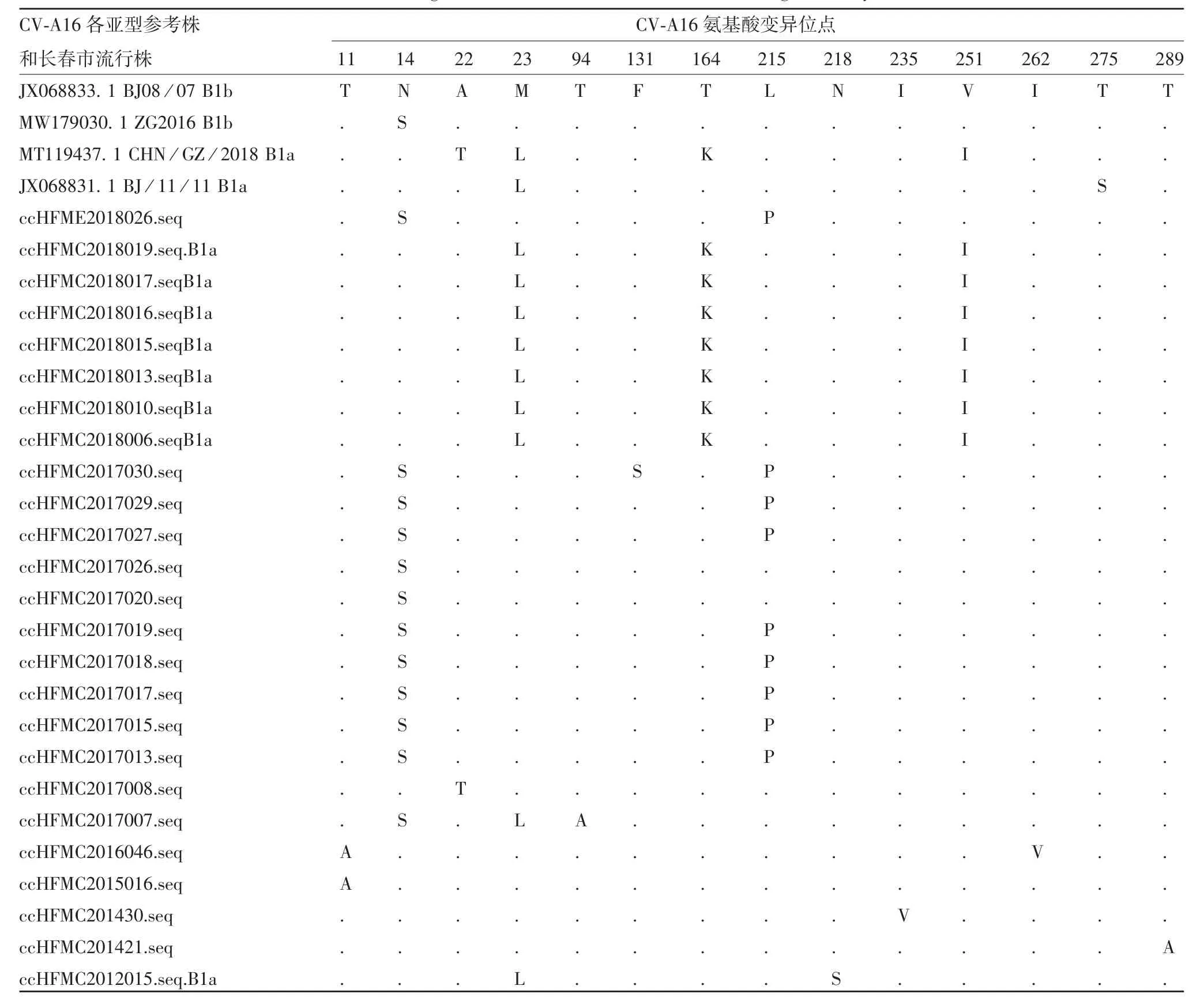
表1 2012—2018年长春市HFMD病例中CV-A16型VP1区氨基酸变异位点Tab.1 Variation sites of amino acids in VP1 region of CV-A16 in HFMD cases in Changchun City from 2012 to 2018
3 讨论
HFMD是一种常见于婴幼儿和儿童的急性肠道传染病,病程复杂、症状多样,多数可自愈,少数病例会出现重症,甚至危及生命[9]。由于HFMD的病原具有多样性,且病原之间经常发生重组变异,因此,随着每年优势病原株发生变迁,就会引起新一轮HFMD的暴发流行[10],在世界范围内造成了严重的疾病负担。
在国外,CV-A16常是大规模HFMD流行时的主要病原体[11]。以往研究多认为CV-A16仅引起轻症患者,但近年来,越来越多的报道显示,CV-A16感染也可引起心肌、脑部和肺部等疾病,甚至引起死亡[12-14]。我国自1981年首次报道CV-A16型HFMD病例以来,其他多个省份相继出现关于CV-A16型肠道病毒引起的HFMD暴发报道[15-16]。CV-A16被分为2个基因型:A和B基因型,而B基因亚型又分为B1和B2 2个亚型。对2012—2018年长春市83株CV-A16病原株VP1区的进化分析结果显示,近7年长春市CV-A16病原株均属于B1基因亚型,其中8条属于B1a亚型,其他均属于B1b亚型。从CV-A16 VP1区系统进化树上可以看出,B1b亚型上包括2012—2018年各年份的毒株,而B1a亚型的分支上仅有2012年的1条和2018年的7条流行株,表明2012—2018年长春市内B1a和B1b亚型属于共循环、共进化的过程,而B1b亚型的CV-A16一直是优势流行株,2018年B1a亚型与B1b亚型共同流行,这与其他省市的报道相同[17-19]。将B1a和B1b分流支分别进行同源性分析,发现B1b亚型分支与B1b亚型参考株同源性为94.5%~99.2%,B1a亚型分支与B1a亚型参考株同源性为92.3%~98.5%,各株间同源性为88.2%~100.0%,与广东、北京、河南等中国其他省份的CV-A16代表株来源相近,表明与其他病原体相比为区域性暴发。
变异位点分析发现,2012—2018年长春市HFMD病例CV-A16毒株中存在14个氨基酸位点变异,12株发现在第14位点上发生N天冬酰胺→S丝氨酸(N14S)的变异,这与2016年成都流行株(MW179030.1 ZG2016 B1b)相同,其中有9株同时存在L215P变异(2017年8株,2018年1株),提示该位点变异可能已演变为较稳定的基因特征被流行传递下来。有9株存在M23L变异,其中8株为B1a亚型,这8株除ccHFMC2012015外,均同时存在T164K和V251I变异,这与2018年广州流行株(MT119437.1 CHN/GZ/2018 B1a)相同。长春市有1株(ccHFMC2017-007)存在N14S、M23L和T94A 3个位点变异,VP1第94~108位氨基酸肽段是一个保守型线性中和抗体,可诱导产生抗CV-A16感染的中和抗体[20],这可能降低亲和力和免疫优势,影响中和抗体结合。
自2008年,芬兰暴发了1起由CV-A6引起的HFMD疫情后[21],全球各地均有CV-A6引起HFMD暴发流行的报道[22-23],长春市2013年也曾暴发过由CV-A6引起的HFMD疫情[24]。目前国内将CVA6主要划分为A~D 4个基因型别,CV-A6原型株为A分支,未在国内流行。对2012—2018年长春市30株CV-A6病原株VP1区的进化分析结果显示,近7年长春市CV-A6病原株均属于D3基因亚型。2012—2018年长春市CV-A6流行株与CV-A6原型株的同源性为82.7%~83.5%,与B1亚型参考株的同源性为81.9%~93.7%,与B2亚型参考株的同源性为82.1%~84.5%,与C1亚型的同源性为85.1%~86.5%,与C2亚型的同源性为83.8%~84.8%,与D1亚型的同源性为89.7%~91.1%,与D2亚型的同源性为89.2%~91.3%,与D3亚型的同源性为92.8%~98.1%,其中与D3亚型的同源性较高,各株间同源性为91.6%~99.8%。目前长春主要流行的CV-A6毒株与国内其他地区流行的型别相同[25],均为D3亚型,并未进化出新的型别。
虽然CV-A10并不是长春市HFMD的主要优势型别,但有研究表明,CV-A10与EV71相似,其感染导致重症和死亡的频率相对较高[26]。根据CV-A10的VP1基因序列,将CV-A10分为A~D 4个基因型,目前国内主要流行从B型和C型亚型逐渐转为D型亚型[26-27]。对2012—2018年长春市10株CVA10病原株VP1区的进化分析结果显示,与CV-A10原型株的同源性为76.3%~77.0%,与B亚型参考株的同源性为78.9%~80.3%,与C亚型参考株的同源性为82.9%~83.8%,与D1亚型参考株的同源性为94.1%~95.4%,与D2亚型参考株的同源性为96.4%~98.4%,与D3亚型参考株的同源性为94.6%~96.1%,其中与D2亚型同源性较高,各株间同源性为96.4%~100.0%。表明长春市2012—2018年CV-A10毒株为D2亚型,与国内其他地区主要流行的型别相同[28],较为稳定。
目前,非EV-A71型肠道病毒引起的HFMD疫情日益加重,而对其研究相对较少,疫苗研发基础薄弱,研发难度大[29]。只有通过加强对HFMD病毒株的进一步研究,掌握病毒变异及进化状态,才能控制HFMD的流行。本研究通过对2012—2018年长春市非EV-A71毒株进行分析,探讨其传播规律和进化情况,对疾病防控具有重要意义。
