茉莉花JsMYB305 转录因子的原核表达及蛋白纯化
2022-04-28伍炳华
万 超,张 月,胡 莉,伍炳华,袁 媛
(福建农林大学园艺学院,福建 福州 350002)
0 引言
【研究意义】茉莉Jasminum sambac (L).Ait 为木犀科(Oleaceae)素馨属(Jasminum)常绿灌木,茉莉花香气馥郁,广泛应用于“茉莉花茶”窨制、香料香精、化妆品药品、食品和园林应用等方面,是著名的芳香植物[1]。香气是茉莉花价值的重要体现方式。研究表明茉莉于夜间开放,主要香气成分包括芳樟醇、法尼烯、乙酸苄酯[2]等,芳樟醇和法尼烯分别占单萜和倍半萜成分的92%和96%。前人通过对茉莉花中一些香气相关酶活性的测定表明,其香气释放是有一定节律的,且其释放是受严格调控的[3],但相应的调控因子及调控机理还少有探究。【前人研究进展】MYB 类转录因子调控植物香气合成在多种植物中均有报道,如矮牵牛的ODO1(ODORANT1)、EOBI(EMISSION OF BENZENOIDS I)、 EOBII(EMISSION OF BENZENOIDS II)交错调控苯丙烷类香气物质代谢[4−6];草莓FaEOBII 调控花瓣中丁香酚生物合成[7];番茄SlMYB75 通过激活脂氧合酶途径和萜类代谢途径中的结构基因启动子,从而调控果实中的香气物质合成[8]。茉莉香气合成途径中的一些重要的结构基因或相关启动子序列已经得到克隆和分离[9−16],启动子区域存在激素响应、光响应、MYB结合位点等多种顺式作用元件,表明有些结构基因的表达可能受MYB 转录因子调控。【本研究切入点】茉莉JsMYB305 与矮牵牛、拟南芥中调控香气合成的转录因子ODO1 及AtMYB21 高度同源,表达量与花朵香气释放同步,且与4 个茉莉花萜类合成酶基因 JsTPS(Terpene synthase)表达特征一致,前期研究中发现JsMYB305 可显著提高茉莉愈伤组织中4 个 JsTPS 基因的表达[17],并激活其启动子的活性,推测JsMYB305 可能通过调控JsTPS 的表达从而调控香气合成。但JsMYB305 是如何调控JsTPS 基因表达的机制却并不明确。【拟解决的关键问题】为进一步研究JsMYB305 调控茉莉香气合成的机理,本研究通过原核表达的方式,在体外获得JsMYB305 重组蛋白,研究结果可为将来研究JsMYB305 与JsTPSs 基因启动子的体外结合及筛选JsMYB305 互作蛋白提供依据。
1 材料与方法
1.1 植物材料
以双瓣茉莉(Jasminum sambac)三年生植物花瓣为材料。取当天21:00 开放的茉莉花朵,用剪刀剪 下后迅速用铝薄纸包好投入液氮中备用。
1.2 原核表达载体的构建
以茉莉花瓣为材料,参照EasyPure®Plant RNA Kit(北京全式金生物技术有限公司)的方法提取花瓣RNA。按 照One-Step gDNA Removal and cDNA Synthesis SuperMix(北京全式金生物技术有限公司)说明书进行第一链cDNA 的合成。以cDNA 为模板,设计 含 有Bam H I 和Sal I 酶 切 位 点 的 特 异 性 引 物JsMYB305-F 和JsMYB305-R(表1)扩 增JsMYB305的编码序列。扩增体系为:上游引物1 μL,下游引物1 μL, FastPfu PCR Super Mix(北京全式金生物技术有限公司)12.5 μL , H2O 8.5 μL, cDNA 2 μL。PCR扩增程序:95 ℃ 5 min;以下程序35 个循环 :95 ℃30 s,63 ℃ 30 s,72 ℃ 1 min;接着72 ℃ 10 min。
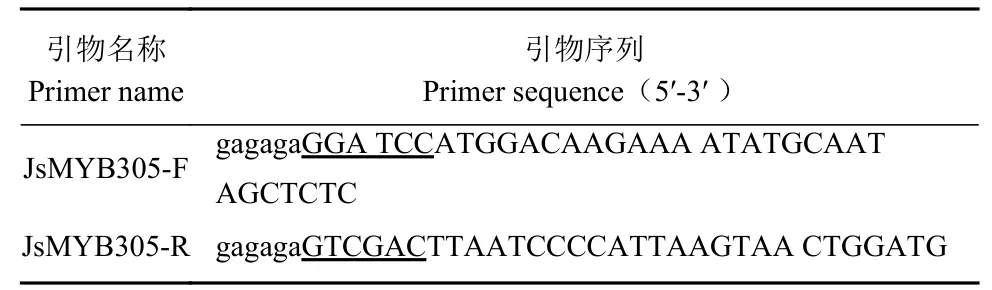
表 1 构建原核表达载体时扩增JsMYB305 编码序列的引物序列Table 1 Primer sequences for amplifying JsMYB305 coding sequence in prokaryotic expression vector construction
使用Bam H I 和Sal I 同时酶切pGEX-4T-1 载体和JsMYB305 扩增产物,酶切体系为:Bam H I 0.5 μL,Sal I 0.5 μL,PCR 产 物200 ng/载 体1 μg,快 切 酶Buffer 2 μL,无菌水补足至10 μL ,37 ℃酶切2 h。分别回收目的基因片段和酶切后的原核表达载体,使用T4 连接酶16 ℃连接过夜。连接体系为:T4 连接酶1 μL,连接酶Buffer 1 μL,载体酶切回收产物1.5 μL,目的基因酶切回收产物3.5 μL,无菌水3 μL。将连接产物取10 μL 转化大肠杆菌DH5α 感受态,涂布于含50 mg·L−1氨苄的LB 平板,37 ℃过夜培养后,挑取单克隆使用载体上的测序引物(F: 5′-GGGCTG GCAAGCCACGTTTGGT-3′,R: 5′-CCGGGAGCT GCATGTGTCAGAGG-3′)进行PCR 鉴定,将阳性菌 液送福州铂尚生物技术有限公司测序。
1.3 重组蛋白的诱导表达
将测序正确的重组质粒通过热激法转化大肠杆菌BL21(DE3)感受态,涂布于含50 mg·L−1的氨苄LB 平板,37 ℃过夜培养。从平板上挑取一个单克隆至5 mL 含氨苄的LB 液体培养基中,37 ℃、200 r·min−1,过夜培养。
取1 mL 过夜培养的大肠杆菌菌液,加入到10 mL含氨苄的LB 液体培养基,37 ℃、200 r·min−1培养至OD600 nm约0.5 时,加入0.2 mmol·L−1异丙基-ß-D-硫代半乳糖苷(IPTG),不加IPTG 的菌液为对照,在37 ℃进 行诱导培养4 h。
1.4 SDS-PAGE 检测
将诱导表达后的菌液10 000 r·min−1离心1 min 收集菌体,用200 μL PBS(NaCl 8 g·L−1,KCl 0.2 g·L−1,KH2PO40.24 g·L−1, Na2HPO41.44 g·L−1)缓冲液重悬菌体。取50 μL 悬菌液加入10 μL 6×Protein loading buffer(北京全式金生物技术有限公司)混匀,沸水浴10 min,12000 r·min−1离心5 min。取10 μL 上清点样至10% SDS-PAGE 胶,参照SDS-PAGE 凝胶制备试剂盒(北京索莱宝科技有限公司)制备,InstaBlue protein stain solution(APE BIO,美国)染色,凝胶成像 分析仪(BIO RAD,美国)观察结果。
1.5 重组蛋白的可溶性检测
确定JsMYB305 可成功诱导表达后,摸索诱导表达可溶性蛋白的条件。设置37 、28 ℃两个诱导温度,诱导时间为4 h。诱导表达结束后用2 mL PBS缓冲液重悬菌体,取50 μL 悬菌液加入10 μL 6×Protein loading buffer,剩余菌液使用冰浴超声破碎2 min,12 000 r·min−1离心5 min,收集上清,沉淀用1.95 mL PBS 重悬,分别取50 μL 上清和沉淀加入10μL 6×Protein loading buffer。将以上样品沸水浴10 min,12 000r·min−1离心5 min,取10 μL 上清点样至10%SDS-PAGE胶,染色,观察结果。
1.6 JsMYB305 重组蛋白的大量诱导及纯化
取5 mL 过夜培养的菌液加入到500 mL 含有氨苄的LB 液体培养基中,37 ℃摇床震荡培养至OD600 nm值约为0.5 后,加入IPTG 至终浓度为0.2 mmol·L−1,28 ℃诱导4 h 后收集全部菌体。使用15 mL Binding buffer(1×PBS,1% Triton x-100)重悬菌体,加入100 mmol·L−1PMSF(Phenylmethanesulfonyl fluoride,苯甲基磺酰氟)至终浓度为1 mmol·L−1,冰浴超声破碎10 min,功率50%,工作10 s,休息10 s。超声结束后在4 ℃,12 000 r·min−1离心20 min。
JsMYB305 重 组 蛋 白 纯 化:取5 mL proteinIso GST Resin(北京全式金生物技术有限公司)加入到层析柱,静置;用5 mL Binding buffer(1×PBS,1% Triton x-100)平衡层析柱;加入离心后的上清,收集流出液;用5 mL Binding buffer(1×PBS,1% Triton x-100)洗涤层析柱,收集流出液;用Elute buffer(50 mmol·L−1Tris,5 mmol·L−1GSH)配 制 浓 度 分 别 为10、20、30 mmol·L−1的还原型谷胱甘肽(GSH)溶液用于洗脱目的蛋白。每个GSH 浓度洗脱体积为5 mL,收集洗脱液,用于SDS-PAGE 检测。
1.7 Western Blot 检测重组蛋白
将 纯 化 后 的 重 组 蛋 白 取50 μL 加 入10 μL 6×Protein loading buffer,沸水浴10 min,12 000 r min−1
离心5 min,取10 μL 上清点样至10%SDS-PAGE 胶,电泳结束将凝胶于Trans blot SD 半干转膜仪(BIO RAD,美国)转移至NC 膜(Ge.Healthcare)上,5%脱脂牛奶室温封闭3 h,GST 单克隆抗体(北京全式金生物技术有限公司)4 ℃孵育过夜,PBST(1×PBS,1‰ tween 20)洗膜30 min,HRP 标记GST 抗体室温孵育1 h,PBST(1×PBS,1‰ tween 20)洗膜30 min,于膜上滴加2 mL eECL 显色液(北京康为世纪生物科技有限公司),凝胶成像分析仪(BIOR AD,美国)观察结果。
2 结果与分析
2.1 pGEX-4T-1-JsMYB305 原核表达载体的构建
以茉莉花cDNA 为模板,JsMYB305-F 和 JsMYB305-R 为引物,扩增获得约600 bp 的特异性条带(图1-A),经测序显示长度为588 bp,与JsMYB305 编码序列相符。回收扩增产物进行双酶切,酶切后进行回收,结果显示回收成功(图1-B)。连接产物转化大肠杆菌后,经载体上的测序引物鉴定,PCR 扩增获得约750 bp 的片段(图1-C),测序结果显示为746 bp,表 明pGEX-4T-1-JsMYB305 重组质粒构建成功。

图1 茉莉JsMYB305 基因编码序列的扩增、酶切回收及菌液PCR 鉴定Fig. 1 Amplification of JsMYB305 coding sequence, recovery of PCR product after enzyme digestion, and PCR identification of bacteria注:M 为DL2000 Marker ;A 为茉莉JsMYB305 基因编码序列的扩增,其中2 为JsMYB305 基因;B 为JsMYB305 基因扩增产物酶切回收后的产物,其中5 为目的基因,1、3、4 为其他基因条带;C 为重组载体pGEX-4T-1-JsMYB305 的菌液PCR 鉴定,1~5 为阳性条带。Note: M: DL 2000 marker; A: amplification of JsMYB305 encoding sequence, 2: JsMYB305; B: amplification product of JsMYB305 after enzyme digestion and recovery; 5: JsMYB305; 1, 3, and 4: bands of other genes; C: identification of recombinant vector pGEX-4T-1-JsMYB305; 1-5: positive bands.
2.2 重组蛋白的诱导表达及可溶性检测
将转化了pGEX-4T-1-JsMYB305 的大肠杆菌于 37 ℃诱导4 h。结果显示,与对照相比,诱导后的菌液在55 kDa 附近出现一条差异条带,大小约为49 kDa(图2-A),与预期的重组蛋白大小相符,表明JsMYB 305 能在大肠杆菌中成功表达。
分别于37 ℃和28 ℃诱导pGEX-4T-1-JsMYB305的表达,将诱导表达后所得的菌液超声破碎后分别取上清和沉淀部分进行SDS-PAGE 电泳检测,发现37 ℃诱导的重组蛋白主要存在于沉淀中(图2-B),28 ℃诱导下,重组蛋白在上清中的含量比37 ℃更高(图2-C),因此28 ℃诱导重组蛋白适合后续大量诱导和纯化。
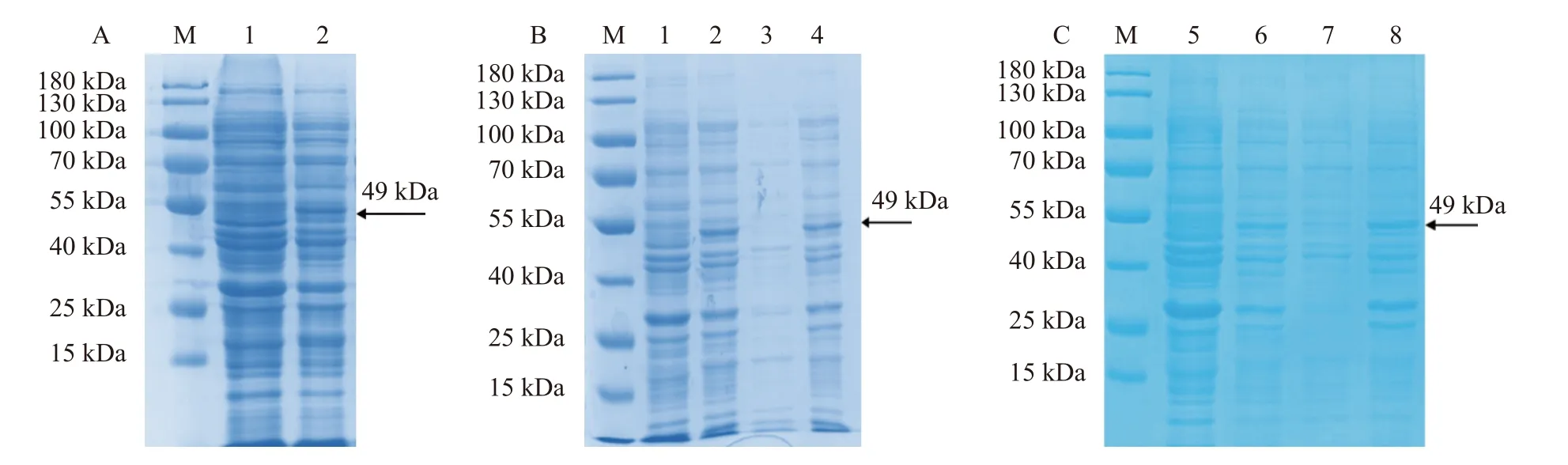
图2 JsMYB305 重组蛋白的诱导表达及可溶性检测Fig. 2 Inducible expression and soluble detection of JsMYB305 recombinant protein注:M:Protein ladder。A:JsMYB305 重组蛋白的诱导表达,1:未诱导的菌体蛋白,2:0.2 mmol·L−1IPTG 诱导后的菌体蛋白。B:37 ℃诱导重组蛋白表达,1:未诱导的菌体蛋白;2、3、4 分别为37 ℃ 0.2 mmol·L−1IPTG 诱导的菌体蛋白、上清、沉淀。C:28 ℃诱导重组蛋白表达,5:未诱导的菌体蛋白,6、7、8 分别为28 ℃ 0.2 mmol·L−1IPTG 诱导的菌体蛋白、上清、沉淀。Note: M: protein ladder; A: induced expression of recombinant protein of JsMYB305; 1: uninduced bacterial protein; 2: bacterial protein induced by 0.2 mmol·L−1IPTG; B: recombinant protein expression induced at 37 ℃; 1: uninduced bacterial protein; 2, 3 and 4: bacterial protein, supernatant, and precipitate, respectively, induced by 0.2 mmol·L−1IPTG at 37 ℃; C: recombinant protein expression induced at 28 ℃; 5: uninduced bacterial protein; 6, 7,and 8: bacterial protein, supernatant, and precipitate, respectively, induced by 0.2 mmol·L−1IPTG at 28 ℃.
2.3 重组蛋白的纯化及检测
基于以上诱导温度的选择,在28 ℃对重组蛋白进行大量的诱导,取上清进行蛋白纯化。上清中的蛋白经过层析柱吸附后,依次使用10、20、30 mmol·L−1的GSH 进行洗脱。不同浓度GSH 洗脱后的洗脱液SDS-PAGE 电泳分析显示,蛋白上清的流出液和洗涤液中不含目的蛋白(图3-A),表明目的蛋白与GST树脂的结合性较好。在20 mmol·L−1的GSH 洗脱下,重组蛋白能被成功洗脱,条带较为单一,表明重组蛋白成功纯化。
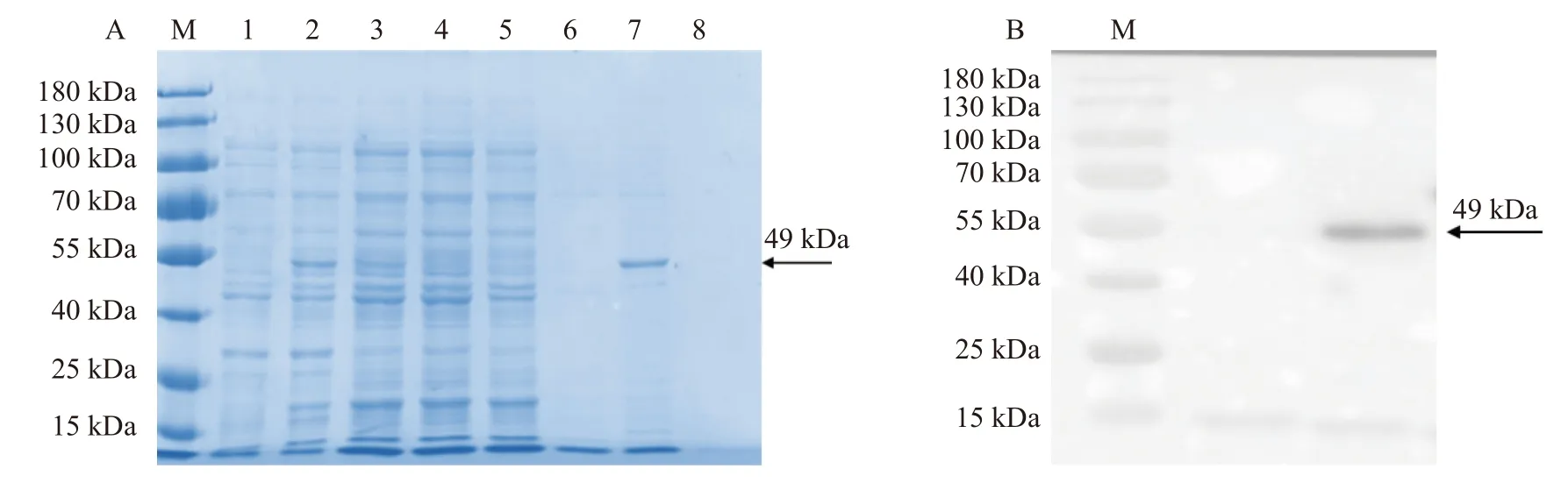
图3 重组蛋白的纯化及Western Blot 检测Fig. 3 Purification and western blot identification of recombinant protein注:M:protein ladder 。A:重组蛋白的纯化 ,1:未诱导的菌体蛋白;2:0.2 mmol·L−1IPTG 诱导后的菌体蛋白;3:超声破碎后的菌体蛋白上清;4:流出液;5:洗涤液;6:10 mmol·L−1GSH 洗脱液;7:20 mmol·L−1GSH 洗脱液;8:30 mmol·L−1GSH 洗脱液;箭头标注的是纯化的重组蛋白。B:纯化蛋白的Western Blot 检测,箭头标注的是JsMYB305 重组蛋白与GST 单克隆抗体杂交后的条带。Note: M: 15-180 kDa protein ladder; A: purification of recombinant protein; 1: uninduced bacterial protein; 2: bacterial protein induced by 0.2 mmol·L−1IPTG; 3: supernatant of bacterial protein after ultrasonic crushing; 4: effluent; 5: washing liquid; 6: eluent of 10 mmol·L−1GSH; 7: eluent of 20 mmol·L−1GSH; 8: eluent of 30 mmol·L−1GSH; arrow points at purified recombinant protein; B: detection of purified protein by western blot; arrow points at band after hybridization of JsMYB305 recombinant protein with GST monoclonal antibody.
将纯化后的JsMYB305 重组蛋白,采用GST 单克隆抗体进行Western blotting 检测,结果表明,纯化后的重组JsMYB305 蛋白,与GST 单克隆抗体有杂交条带,且条带大小与实际相符合(图3-B)。结果 说明纯化后的重组蛋白为JsMYB305 重组蛋白。
3 讨论
外源基因能否在原核表达系统中成功表达受到多种因素影响,如目的基因本身特性、诱导温度和时间等[18−19]。温度会显著影响原核蛋白的表达,在高温诱导时,大肠杆菌的生长速度较快,蛋白表达的速度会随之增快,导致表达的蛋白不能正确折叠,易形成包涵体[20−21]。本研究中,相比37 ℃诱导温度,28 ℃能诱导JsMYB305 蛋白更多地在上清中存在,与前人的研究结论一致。表达载体和表达菌株的选择同样对蛋白的诱导表达起十分重要的作用,如AtSTK 基因利用大肠杆菌Rosetta 菌和pET-32a 载体可以更好地表达出目的蛋白[22]。孙伟等[23]利用大肠杆菌Transetta(DE3)菌株表达ZmAGO18b 时发现,pET-28a 能够诱导出目的蛋白的表达,而pGEX-6p 基本没有检测到目的蛋白。本研究也发现类似的现象,将JsMYB305 编码序列构建到pQE30 载体并在BL21 株系诱导表达时基因无法成功诱导表达,换用pGEX-4T-1 表达载体后能成功诱导JsMYB305重组蛋白的表达和纯化。这可能是谷胱甘肽S-转移酶(Glutathione S-transferase,GST)与目的蛋白融合后,能提高目的蛋白的可溶性[24],从而成功表达目的蛋白。
在研究蛋白质的相互作用及筛选已知蛋白的未知互作蛋白方面,GST 标签有着广泛的应用。为验证两个蛋白间是否具有相互作用,朱炳森[25]和王意程[26]将目的蛋白分别构建到带HIS 和GST 标签的原核表达载体,诱导表达并纯化后,利用pull down 试验验证两个蛋白间的相互作用。赵杰等[27]和窦万福等[28]分别利用 GST-pull down 技术联合LC-MS/MS,鉴定出128 个与毛白杨天冬氨酸蛋白酶PtoAED3 和53个与柑橘抗溃疡病转录因子CsBZIP40 特异结合的互作蛋白。本研究中也选择了GST 标签,为后续利用pull down 技术筛选JsMYB305 的互作蛋白提供基础。
本研究首次将茉莉花中调控香气的转录因子JsMYB305 在大肠杆菌中诱导表达,并能通过GST 柱层析纯化获得,可为后续利用凝胶阻滞(EMSA)试验分析JsMYB305 对TPS 基因启动子的作用,以及利用GST-pull down 联合LC-MS/MS 及免疫共沉淀(coip)筛选JsMYB305 的互作蛋白奠定坚实基础,为深入分析JsMYB305 蛋白功能,并基于此基因功能探索茉莉花香气分子育种提供借鉴。
