油柰PsWRKY33 基因启动子的克隆与表达分析
2022-04-28陈永萍何水林刘志钦陈桂信蒋际谋
陈永萍,何水林,刘志钦,陈桂信 ,蒋际谋
(1. 福建省农业科学院果树研究所/福建省龙眼枇杷育种工程技术研究中心,福建 福州 350013;2. 福建农林大学农学院,福建 福州 350002;3. 福建农林大学园艺学院,福建 福州 350002)
0 引言
【研究意义】油柰(Prunus salicina lindley)是蔷薇科(Rosaceae)李属(Prunus)核果类落叶果树,是福建省特色水果和稀优李资源,具有较高的经济价值[1],但逆境胁迫(如极端温度[2]、干旱[3]等)均会影响油柰的生长发育,严重时甚至造成减产[4]。植物对各种逆境胁迫的响应需通过一系列信号网络的感知、传导和传递[5],故研究植物在逆境胁迫下的信号感知传递过程及其分子机制,对研究植物抗逆机理,提高植物抗性具有重要意义。【前人研究进展】在植物应答各种非生物胁迫的过程中,作为植物体内最大转录因子家族之一的WRKY转录因子在调节植物应答逆境胁迫的防御反应中发挥着至关重要的作用[5−8]。研究表明,在拟南芥中,ABA、干旱、盐处理能诱导增强AtWRKY33 和AtWRKY25的表达,通过表达可增强拟南芥对盐的耐受性[9]。分别在拟南芥中过表达TaWRKY33[10]和CkWRKY33[11]可提高转基因株系在高温胁迫和干旱胁迫下的存活率。在紫花苜蓿中过表达MsWRKY33 可以提高转基因材料的耐盐性[12]。在葡萄中VaERF092 通过结合VaWRKY33 启动子GCC-box 调控VaWRKY33,从而增强植物的耐寒性[13]。在油柰上,外源SA 处理下能显著抑制PsWRKY22启动子在拟南芥中的表达,表明SA 能调控PsWRKY22的表达[14]。【本研究切入点】基于前人相关研究发现有关油柰的抗性基因研究相对较少,本研究前期通过对油柰cDNA 文库的筛选,获得一个能响应SA 的WRKY 基因并命名为PsWRKY33,经克隆PsWRKY33启动子全长以及缺失序列构建含有GUS报告基因的载体,通过转基因到拟南芥来分析PsWRKY33 启动子不同片段在不同处理下的表达模式。【拟解决的关键问题】利用模拟各种非生物胁迫,探测转基因PsWRKY33 启动子不同片段在不同胁迫处理下的应答模式,以期为进一步研究油柰WRKY基因生物学功能 及其调控机制提供科学数据。
1 材料与方法
1.1 植物材料和生长条件
以福建古田地区标准室外嫁接育苗的一年生翠屏晚柰品种作为研究对象。取叶片样品,迅速冷冻于液氮中用于提取油柰基因组DNA。
将野生型(Columbia)拟南芥以及转基因的种子播种于装有经高压灭菌后的基质中[V(泥炭苔):V(蛭石)=2∶1]。置于(25±1) ℃、光照强度为100 μmol·ms−1(12 h 光 / 12 h 暗循环)、相对湿度为60%的条件下培养。转基因拟南芥种子在4 ℃暗处理3 d 后用灭菌水洗1 次,后用75 %酒精洗3~5 min,最后用无菌水洗5 次。转基因T0 和T1 代种子播种在含50 mg·L−1潮霉素的½MS 固体培养基中进行筛选。将培养皿转移到培养箱中,在(23±1) ℃、相对湿度8 0%的条件(12 h 光/12 h 暗循环)下培养。
1.2 生物信息学分析
在NCBI 数据库blastp 比对获得与PsWRKY 氨基酸序列同源性较高的5 种植物的蛋白序列,利用MEGA 6.06 软件将以上6 种植物氨基酸序列进行同源比对,构建系统进化树。
利用PlantCARE 数据库(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析预测PsWRKY33 5 ′端上游启动子区域存在的顺式作用调控元件。
1.3 载体构建、植物转化、胁迫处理
利用染色体步移法从油柰基因组DNA 中克隆出PsWRKY33 启动子片段,在此基础上利用PCR 技术扩增出3 个不同缺失片段(表1),载体构建过程、转 基因过程、胁迫处理等过程参照陈永萍等[14]的方法。

表 1 引物序列Table 1 Sequences of primers
1.4 组织化学染色和荧光法检测GUS 活性
采集不同胁迫处理下的7 日龄T3 转基因株系的叶片,以及T3 代转基因株系的叶片、花序和角果进行GUS 染色并拍照,具体操作方法过程参照陈永萍等[14]的方法。
参照陈永萍等[14]的方法,采用荧光法定量测定胁迫处理下的转基因植株GUS 相对活性。数据整理后用SigmaPlot 12.5 软件作图。所有试验重复3 次。采 用t 检验进行统计学分析。
2 结果与分析
2.1 PsWRKY33 系统发育分析
为确定PsWRKY 基因与其他物种之间的系统发育关系,将获得的PsWRKY 氨基酸序列与梅花PmWRKY33(accession no. XP_008231402.2)、欧 洲甜樱桃PaWRKY24L(accession no. XP_021810565.1)、东京樱花PyWRKY33(accession no. PQM41056.1)、巴旦木PpWRKY24(accession no. XP_007208290.1)和拟南芥AtWRKY33蛋白序列(accession no. NP_181381.2)进行系统进化树分析。结果表明,PsWRKY 与拟南芥AtWRKY33 具有较高的序列相似性。因此,我们将该基因命名为PsWRKY33(图1)。

图1 系统进化树Fig. 1 Phylogenetic tree
2.2 PsWRKY33 启动子序列的克隆及顺式元件分析
利用染色体步移法获得PsWRKY33 的ATG 起始密码子上游1 872 bp 的片段,并利用PlantCARE 网站对其启动子序列进行预测分析。结果发现(图2),PsWRKY33 启动子含有大量参与激素诱导响应、逆境胁迫响应等顺式作用元件,如分别响应低温、生长素、SA、ABA、乙烯等的LTR、AuxRR-core、TCA、ABRE、ERE 等顺式元件。在此基础上,通过对不同的响应元件进行切除获得不同缺失片段,并分别构建到pMDC163 表达载体上(图3)。经浸花法分别转化大野生型拟南芥中,经潮霉素筛选与PCR 检测转基因阳性植株(图4),最终获得含有pPsWRKY33、P1、P2、P3 的转基因株系,分别命名为pPsWRKY33::GUS、 pPsWRKY33-P1::GUS、 pPsWRKY33-P2::GUS和 pPsWRKY33-P3::GUS。

图2 PsWRKY33 基因启动子序列Fig. 2 Cis-elements of PsWRKY33 promoter注:-60 表示ATG 上游60 个核苷酸。红色粗体字母加框表示预测的顺式作用元件序列。pPsWRKY33、P1、P2、P3 加箭头加黑色下划线表示为4 个片段(−1 872、−852、−357 和−271 bp)的特异性引物,箭头表示它们的方向。Note: -60 represented the 60 nucleotides upstream of the ATG. Bold red letters with boxes indicated the predicted sequence of cis-acting elements. pPsWRKY33, P1, P2 and P3 add arrows and black underlines represented the specific primers of four fragments (−1 872, −852, −357 and −271 bp), the arrows indicated their direction.

图3 PsWRKY33 启动子各片段的GUS 载体Fig. 3 Schematic diagram of GUS vector of individual PsWRKY33 promoter fragments注:不同启动子区域的数目表示PsWRKY33 翻译起始ATG 密码子上游的核苷酸位置。 attR1 和attR2 限制性位点: 将pDMC163 载体中的CaMV35S 启动子替换为PsWRKY33 启动子片段。LB 和RB:代表T-DNA 的左右边缘;Hyg:潮霉素标记基因; GUSA: GUS 基因编码区域。 黄色的条形图代表PsWRKY33::GUS、 PsWRKY33-P1::GUS、 PsWRKY33-P2::GUS 和PsWRKY33-P3::GUS 结构的-1 872 bp、-852 bp、-357 bp 和-271 bpPsWRKY33 启动子片段。Note: The number of different promoter regions represents the nucleotide position upstream of the PsWRKY33 translation initiation ATG codon.The CaMV35S promoter in pDMC163 vector was replaced by PsWRKY33 promoter fragment by attR1 and attR2 restriction sites. LB and RB represent the left and right edges of T-DNA; Hyg, hygromycin B marker gene; GUS is the region that codes for the GUS gene. The yellow bars represent the -1 872 bp, -852 bp, -357 bp and -271 bp PsWRKY33 promoter fragments of PsWRKY33::GUS, PsWRKY33-P1::GUS,PsWRKY33-P2::GUS and PsWRKY33-P3::GUS, respectively.
2.3 PsWRKY33 启动子在不同组织的活性分析
利用GUS 组织化学方法分析4 个启动子片段在转基因拟南芥T3 幼苗中的表达特征。结果表明,在转基因拟南芥T3 代苗的叶片、花序和角果中,PsWRKY33 启动子的4 个缺失片段存在不同的表达模式。pPsWRKY33::GUS在叶片、花序和花序梗中均检测到较强的GUS 活性,且随着启动子片段缩短,叶片和花序梗中的GUS 活性越弱。4 个缺失片段在角果上均未发现明显的GUS 活性,其中在pPsWRKY33::GUS、pPsWRKY33-P1::GUS 和pPsWRKY33-P2::GUS 的角果荚 底部观察到轻微的GUS 活性(图5)。
2.4 4个PsWRKY33启动子片段对不同胁迫的响应特征
为确定PsWRKY33 启动子是否能响应不同的逆境胁迫,故对7 日龄T3 转基因拟南芥植株进行ABA、SA、MeJA、ACC(ETH)、NaCl、MAN 和低温(4 ℃)处理12 h,检测其GUS 染色(图6)以及相关GUS酶活(图7)。经GUS 染色结果发现PsWRKY33 启动子4 个缺失片段在不同胁迫处理下的GUS 活性强弱不同。不同PsWRKY33 启动子片段在4 ℃低温处理下,各片段GUS 活性均强于对照组,而在SA 处理下4 个缺失片段的GUS 活性最弱,其中在全长、P1 和P2 缺失片段的植株发育真叶中检测到微弱的GUS 活性,而在P3 缺失片段的转基因植株中未能明显观察到GUS 活性。同时发现4 个PsWRKY33 启动子片段转基因植株在对照组及NaCl 和ABA 处理下随着片段缺失长度的增加,其GUS 活性逐渐减弱;而在MAN 和ETH 处理下,随着越长的片段缺失,其GUS 活性反而明显增强。而在MeJA 处理下,全长和P3 缺失片段植株的GUS 活性强于P1 和P2 缺失片段植物的GUS 活性(图6)。
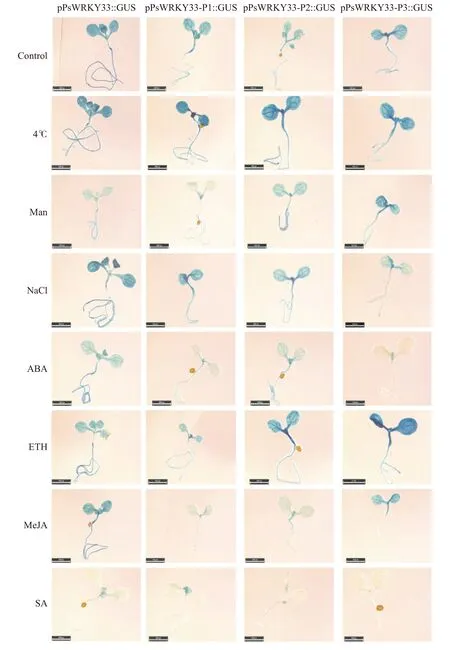
图6 不同胁迫处理下转基因拟南芥GUS 组织染色Fig. 6 GUS staining of transgenic Arabidopsis under different stresses
为进一步定量分析各片段GUS 活性在各胁迫下的变化趋势,同时对各胁迫下的4 个PsWRKY33 启动子缺失片段转基因拟南芥幼苗进行GUS 相对酶活测定分析。结果图7 可见,不同长度的GUS 活性在不同处理下呈现出不同的变化趋势。在对照组、4 ℃低温、盐、ABA、SA 处理下,各片段GUS 活性表现出片段越短,其GUS 活性越低的变化模式,并且发现在除了乙烯处理外的各个处理下,全长GUS 活性均不同程度上高于3 个缺失片段。在乙烯和MeJA处理下,全长和P3 的GUS 活性高于P1 和P2 的GUS活性。相比于对照组,4 ℃低温处理下各片段GUS活性表现出不同程度上地提高,而ABA 和SA 处理下,4 个片段GUS 活性显著降低。总体上GUS 活性变化趋势与GUS 染色结果相似。
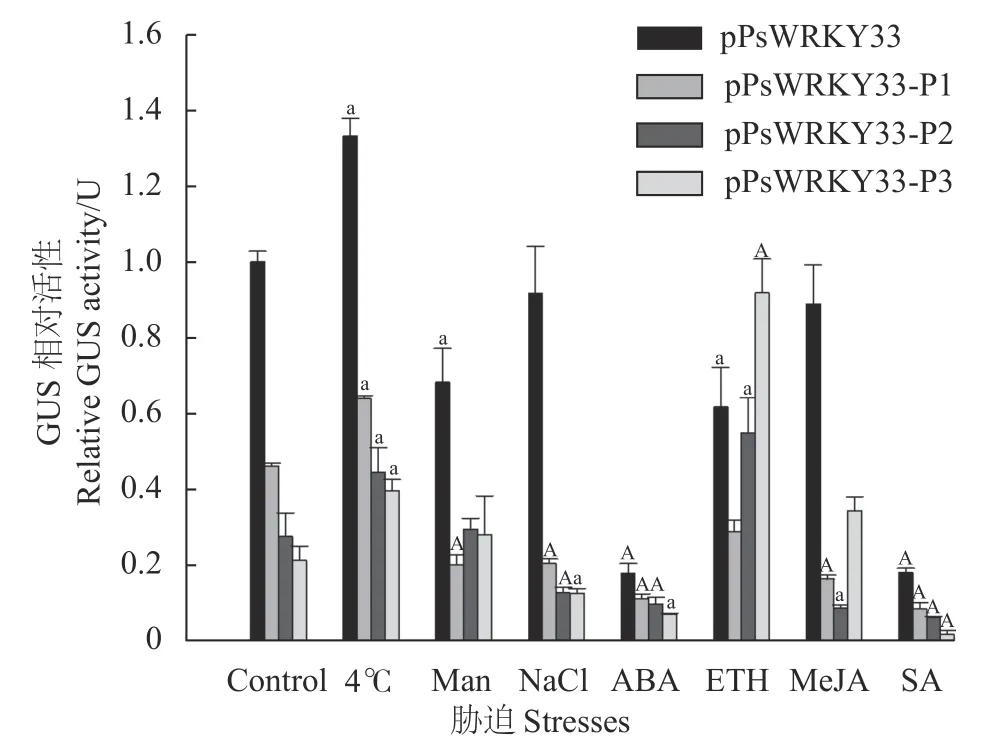
图7 转基因拟南芥在不同胁迫处理下的GUS 相对活性Fig. 7 GUS relative activity in transgenic Arabidopsis under different stresses注: 图中不同大、 小写字母表示同一处理达极显著(P<0.01)、显著(P<0.05)差异。Note: Different uppercase and lowercase letters indicate highly significant (P<0.01) and significant (P<0.05) differences in the same treatment .
综上所述,在对照组及4 ℃、盐、ABA 和SA 的胁迫下,随着缺失的片段越长,其GUS 活性下降越明显。这表明,PsWRKY33 启动子的3 个缺失片段失去 对4 ℃、ABA、SA 和NaCl 胁迫响应的特性。
3 讨论与结论
WRKY 转录因子是植物体内最大的转录因子家族之一,广泛参与植物生长发育过程,以及对各逆境胁迫(生物、非生物及激素胁迫)的响应[15]。WRKY转录因子通过激活相关信号通路[16],与各种胁迫应答基因启动子区的顺式作用元件相互作用,正向或负向调节激素相关基因或各种胁迫防御应答基因的表达,参与防御反应,促使植物在分子、生理生化等水平做出调节以适应各种环境胁迫[17]。启动子是基因转录起始位点前的一段重要序列,对基因的转录调控起着重要影响[18]。油柰上发现PsWRKY22 基因能被外源激素SA 处理诱导差异表达[19],在拟南芥中过表达PsWRKY22 基因启动子[14]发现该基因启动子在SA 处理下表达被显著抑制,为研究同样能被外源激素SA 处理诱导差异表达的PsWRKY33 基因[19]转录调控机制,本研究借鉴PsWRKY22 基因启动子[14]应答不同胁迫处理的研究方法,以期探讨PsWRKY33启动子的功能和表达调控的分子机制。
启动子序列上包含核心元件及一些响应逆境胁迫的顺式元件,研究顺式作用元件对启动子以及基因功能的作用,对分析目的基因的功能具有重要意义[18]。PsWRKY22 基因启动子上含有响应多个响应激素、逆境相关的顺式元件[14],尤其含有SA 响应元件TCA-element,经试验发现PsWRKY22 基因启动子在SA 处理下活性被抑制表达与该元件有关。本研究经网页预测也发现PsWRKY33 基因启动子区域含有多种响应激素、逆境相关的顺式作用元件,如ABA响应元件ABRE、生长素响应元件AuxRR-core、ETH响应元件ERE、GA 响应元件P-box 和TATC-box、SA 响应元件TCA、低温响应元件LTR、响应干旱和ABA 元件MYB、响应干旱、ABA 和低温胁迫元件MYC、响应机械损伤元件WUN-motif,推测该基因的表达可能会受ABA、ETH、低温等的诱导或抑制。
高永峰等[20]分析显示SlWRKY31 基因启动子上含有分别响应热胁迫、干旱、ABA 和SA 的HSE、MBS、ABRE 和TCA-element 元件,通过不同胁迫处理后发现该启动子显著受到盐、甘露醇、SA、ABA和42 ℃高温的诱导表达,说明SlWRKY31 基因启动子是一个可以响应多种逆境胁迫的诱导型启动子[14]。刘志钦等[21]通过构建CaWRKYK5 启动子5′端不同缺失片段的载体,经青枯病菌、激素等处理分析后得出CaWRKYK5 启动子应答青枯菌、MeJA 等的顺式元件区域。本研究经多种胁迫处理,结果发现在SA 处理下,PsWRKY33 启动子GUS 染色和活性均显著低于对照组,说明PsWRKY33 可能参与油柰对过量SA 胁迫的响应,进而通过调节SA 去响应逆境胁迫。而在极端温度(4 ℃低温)胁迫下,GUS 染色和活性均高于对照,说明该基因可能是受低温诱导的。虽然在本研究没有预测到应答JA 的顺式作用元件,但是在JA 的处理下,PsWRKY33 启动子驱动的GUS 报告基因可以被激活,说明可能存在其他的未知的调控机制。
PsWRKY33 基因启动子能被外源ABA、SA 抑制转录表达活性,而受到低温诱导表达,可能与其含有的ABRE、LTR 和TCA 元件有关。在ETH 处理后,pPsWRKY33 和pPsWRKY33-P1 的GUS 活性受到抑制,而pPsWRKY33-P2/P3 的GUS 活性被诱导上调表达,推测−1 871 ~ −852 bp 区域可能含有对ETH 应答起着负调控的顺式元件,而−357 bp ~ −1 (ATG)区域里可能存在应答ETH 的核心元件。
本研究检测不同胁迫处理下PsWRKY33 基因启动子及其缺失片段的GUS 活性,但还未确定具体发挥作用的顺式作用元件,以及对下游基因表达起促进或抑制作用的是由于单个顺式作用元件调控引起的,还是由多个顺式元件协同作用引起的结果[22],已有文献表明,通过对启动子上的元件进行缺失处理分析的方法,是作为鉴定启动子顺式作用元件功能的重要手段之一[23]。Rushton 等[24]通过对PR1 基因启动子进行缺失及功能分析后发现,对启动子Wbox 元件核心序列进行突变后启动子会丧失响应病原菌诱导的能力,故日后可通过对PsWRKY33 基因启动子上的顺式元件碱基进行点突变或其他方法来确定其启动子应答各逆境胁迫的核心元件位置,为后续对PsWRKY33 基因及其启动子的功能和表达调控的分子机制研究奠定坚实的基础。
