温度变化对休耕期冬水田天敌捕食猎物的影响及天敌耐冷性分析
2022-04-25孙佳琦王光华任应党白耀宇
孙佳琦,王光华,任应党,陈 琴,白耀宇,*
1 西南大学 植物保护学院,重庆 400716
2 河南省农业科学院 植物保护研究所,郑州 450002
3 重庆市沙坪坝区农业技术服务中心,重庆 400030
在我国南方稻作区,冬水田是指水稻收割后休耕的秋、冬及春季用来蓄水的稻田,既是一种耕作制度又是一种蓄水工程,广泛分布于四川、重庆等西南丘陵山区[1]。目前,重庆稻区的冬水田种植面积较大,占其水稻栽种面积的59%以上,约为1.1 × 106hm2;种植模式以年种一季中稻或中稻结合再生稻为主,水稻从9月收割后休耕至翌年5月移栽的时间长达6—8个月[2]。
由于水稻(特别是再生稻)收割后稻田休耕系统水稻主要害虫及植食性昆虫种类和数量急剧减少,节肢动物群落又存在重建的过程[3],而捕食性天敌需要在重建的稻田节肢动物群落中发现其主要猎物,这就使得长达半年之久的冬水田休耕期捕食性天敌的生存问题受到了关注[4—5]。我们前期调查指出,冬水田以天台刺齿虫兆Homidiatiantaiensis等弹尾虫为代表的主要腐食者功能群对拟水狼蛛Piratasubpiraticus和拟环纹豹蛛Pardosapseudoannulata及青翅蚁形隐翅甲Paederusfuscipes等优势捕食者种群发生起到关键作用[4—5]。尽管如此,这些捕食者的捕食作用也易受到冬水田休耕季节温度变化的影响。研究指出,温度能显著影响蜘蛛等天敌的种群数量[6]及捕食率和捕食量[7—10]。捕食功能反应是研究捕食者捕食作用有效性和评价其捕食潜力的经典方法[11],从捕食功能反应方程可以得出捕食者的瞬时攻击率、对猎物的处理时间和搜寻效应值及猎物密度对天敌捕食量的影响规律[12—13]。同时,捕食过程中捕食者自身密度的干扰效应及对猎物的捕食偏好性也能影响其捕食效率和捕食量[14]。另外,过冷却点(Supercooling point,SCP)是衡量变温动物耐冷性强弱的重要指标,也是其抵御低温环境的一种生理现象;捕食者SCP越低,其抗冷能力就越强[15]。而随季节温度变化捕食者的种群发生量和捕食作用也与其对低温的耐冷性有关[16]。总之,温度与捕食者的捕食作用密切相关[12—13],而目前研究稻田系统拟水狼蛛、拟环纹豹蛛和青翅蚁形隐翅甲等优势捕食者的捕食功能反应及猎物偏好性等主要以模拟作物生长季节的温度为主[17—19],罕有模拟冬水田休耕季节较低温度下捕食者的捕食作用报道,更无分析冬水田捕食者耐冷特性与其捕食作用关系的研究。
休耕季节冬水田中数量巨大的天台刺齿虫兆和以成虫越冬的水稻害虫白翅叶蝉Thaiarubiginosa往往在稻田生境混合发生[4],且其主要游猎型天敌青翅蚁形隐翅甲、拟水狼蛛和拟环纹豹蛛以成虫和成蛛、若蛛等越冬[4,20—22]。这些捕食者能在越冬期间白天较低温度下出洞觅食[5,21];而重庆丘陵低海拔地区休耕季节冬水田生境温度较高,从当年9月至翌年3月,稻田土壤温度为4.4—26.4 ℃,均温为13.1 ℃[23]。因此,我们以冬水田休耕季节前后期(当年9—11月和翌年2—3月)和中期(当年12月—翌年1月)白天较多频次波动范围中的两种温度为代表,在室内条件下研究温度变化对这3种天敌捕食天台刺齿虫兆和白翅叶蝉、自身密度干扰反应及猎物偏好性的影响,测定这些捕食者的SCP等耐冷性参数并分析与其捕食作用关系,以期为更好地保护冬水田休耕季节这些天敌的种群发生和翌年寻找更加有效的水稻害虫“绿色防控”策略提供基础理论依据。
1 材料与方法
1.1 试验时间和地点
于2020年12月至2021年3月在西南大学植物保护学院昆虫生态与转基因植物生物安全评价实验室进行捕食性天敌的捕食作用试验。于2021年3月在河南省农业科学院进行捕食性天敌的耐冷性测定试验。
1.2 试验材料
于2020年12月至2021年3月在重庆市璧山区(106°02′—106°20′E,29°17′—29°53′N)大路街道和河边镇休耕期冬水田采集青翅蚁形隐翅甲P.fuscipes成虫、拟水狼蛛P.subpiraticus和拟环纹豹蛛P.pseudoannulata成蛛和亚成蛛、天台刺齿虫兆H.tiantaiensis和白翅叶蝉T.rubiginosa成虫(分别简称隐翅甲、狼蛛成蛛或亚成蛛、豹蛛成蛛或亚成蛛、刺齿虫兆和叶蝉)。采回的捕食者均使用刺齿虫兆和叶蝉混合猎物在培养皿中饲喂24 h后再分别置于后续试验温度(22 ℃和12 ℃)中饥饿24 h供试。刺齿虫兆和叶蝉在室温下饲养供试。
1.3 试验条件
捕食者对猎物的捕食功能反应、捕食者自身密度对其捕食的干扰效应以及捕食者的捕食偏好性试验均在人工气候箱(RXZ型,江南仪器制造厂,中国)中进行。该气候箱温度分别为(22 ± 1)℃和(12 ± 1)℃,相对湿度和光周期均为(85 ± 10)%和12L:12D。
1.4 研究方法
1.4.1供试捕食者的体长测定
由于本试验中隐翅甲和蜘蛛个体均采自休耕季节冬水田,为减少这些捕食者个体的体型大小(特别是狼蛛和豹蛛亚成蛛)对试验结果的影响,我们对这些供试捕食者个体的体长做了测定分析。
1.4.2捕食者对刺齿虫兆和叶蝉的捕食功能反应和搜寻效应
在试验开始前,将粗细和长度基本一致的稻杆一端用湿润无菌脱脂棉包裹后竖立于塑料管(直径3 cm,高10 cm)底部,并在塑料管的管壁上刻划易于捕食者上下爬行的线条。在试验开始时,先按照5、10、15、20和25头/管密度梯度分别接入刺齿虫兆或叶蝉成虫,然后分别挑取1头饥饿后的捕食者放入不同猎物密度的塑料管中,最后将塑料管顶部用纱布封口后放置于人工气候箱内开始试验。每个猎物密度重复10次。24 h后统计每个塑料管中猎物的剩余数量,计算捕食者的日捕食量,日捕食量=(猎物初始数量-剩余数量)/1 d。死亡的叶蝉在体视镜下观察后,若有残缺、孔洞等情况,则记为被捕食。
单头捕食者对猎物的捕食功能反应类型通常符合Holling II型[12],圆盘方程为Na=aNTr/(1+aThN)[24];式中,Na为被捕食的猎物数,a为瞬时攻击率,N为猎物密度,Tr为总试验时间(本试验中为1 d),Th为处理单头猎物的时间。为了拟合回归曲线并计算参数a和Th,对该方程取倒数后变形,使其线性化,即1/Na=1/aNTr+Th/Tr;在该方程式中,1/Na和1/N分别看作该方程的因变量和自变量。在本试验中,除了计算上述方程中的主要参数a和Th外,也计算能综合反映捕食者对猎物的捕食能力参数a/Th值。利用上述反应模型参数,分别计算每种捕食者对刺齿虫兆和叶蝉的搜寻效应参数S,计算公式为:S=a/(1+aThN)。
1.4.3捕食者自身密度对其捕食的干扰效应
先在上述1.4.2处理后的塑料管中按照1、2、3、4和5头/管的天敌密度梯度接入捕食者,然后分别在每个塑料管中接入15头刺齿虫兆或叶蝉,最后将塑料管顶部用纱布封口后放置于人工气候箱内开始试验。每个捕食者密度重复5次。24 h后统计每个管中猎物的剩余数量,计算日捕食量。若捕食者个体间发生同类相残现象,则这一重复视为无效。
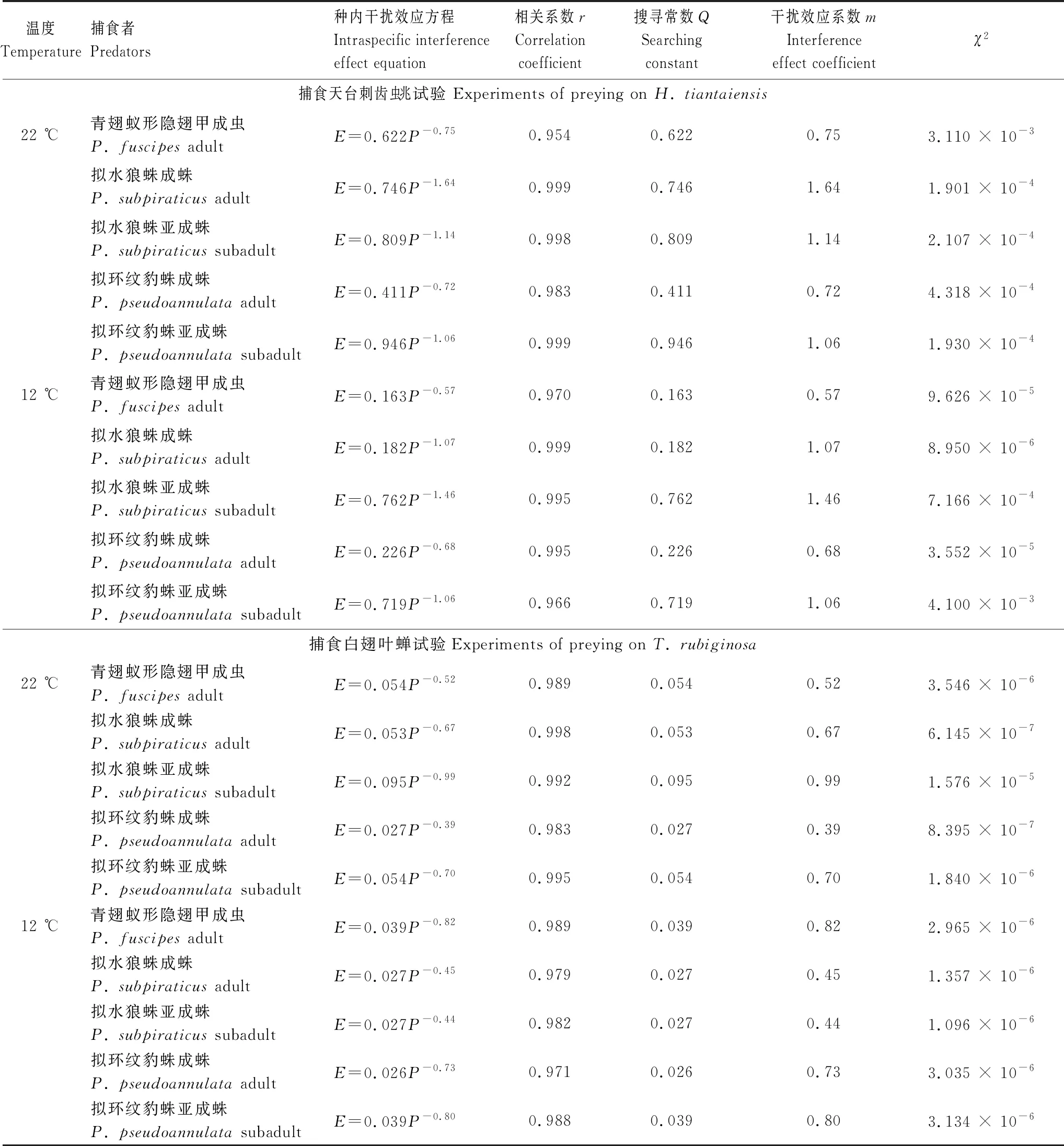
捕食者自身密度的干扰效应曲线采用Hassell和Varley[25]干扰反应模型E=QP-m拟合;式中,E为捕食作用率,Q为搜寻常数,P为捕食者密度,m为干扰系数。捕食作用率E的计算公式为:E=Na/NP;式中,Na和N分别为被捕食的猎物数和猎物初始密度。
1.4.4 捕食者的捕食偏好性分析
先在上述1.4.2处理后的每个塑料管中分别接入1头一种捕食者和20头不同组合的两种猎物,然后将每个塑料管顶部用纱布封口后放置于人工气候箱内开始试验。其中,猎物组合设置3个处理,分别为5头叶蝉和15头刺齿虫兆、10头叶蝉和10头刺齿虫兆、15头叶蝉和5头刺齿虫兆。每个处理重复5次。24 h后统计每个管中猎物剩余数量,计算日捕食量。猎物选择性指数(D)的计算公式为:D=Np1N2/(Np2N1)[26];式中,Np1和Np2分别为刺齿虫兆和叶蝉的被捕食量,N1和N2分别为刺齿虫兆和叶蝉的初始密度值。当D>1时,捕食者对猎物具有捕食偏好性。
1.4.5 不同温度快速冷驯化后捕食者过冷却点和结冰点的测定
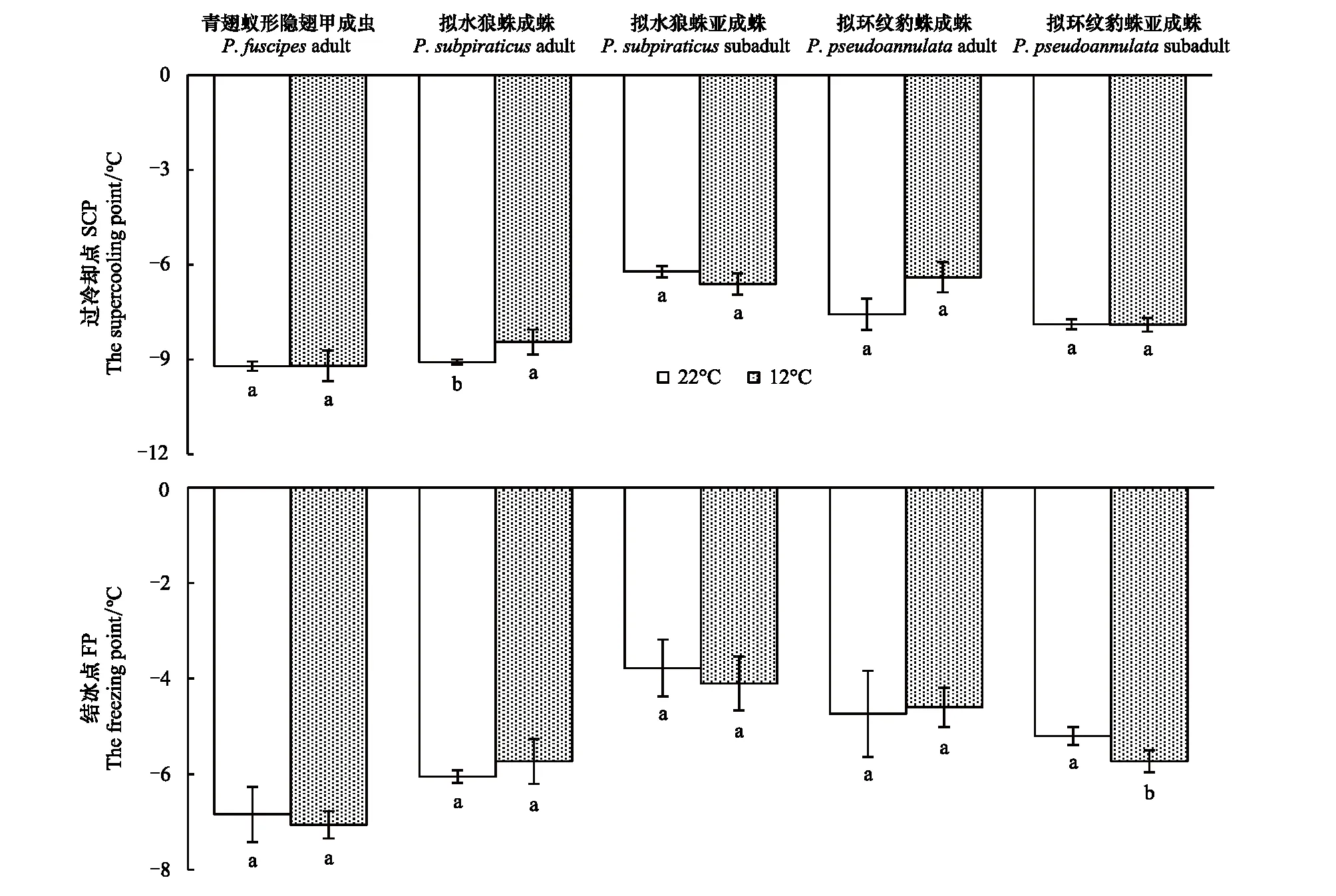
利用DW- 40L348型医用低温保存箱(海尔有限公司,中国)和SUN-V型智能昆虫过冷却点测定仪(北京鹏程电子公司,中国)及仪器配套软件测定捕食者的过冷却点(SCP)和结冰点(Freezing point,FP)。这些捕食者快速冷驯化方法参照刘佳等[15]的方法略有修改。在测试前,先将隐翅甲、狼蛛成蛛和亚成蛛、豹蛛成蛛和亚成蛛保存于1.5 mL离心管中,然后将这些离心管分别置于12 ℃和22 ℃的人工气候箱内对这些捕食者驯化2 h,最后将它们取出并放置于室温下恢复1 h后进行SCP和FP的测定。在测定时,先将测定电极与虫体充分接触并塞入棉花固定(注意不要使虫体受伤),然后将它们迅速放入冰箱内降温。在该降温过程中,计算机软件会记录虫体温度变化曲线,当该曲线跳跃式上升时的温度即为SCP,而上升结束后开始下降时的温度则为FP。每个温度处理测定的这3种不同虫态的捕食者均为30头,其中隐翅甲成虫及狼蛛和豹蛛成蛛为雌雄各15头。
1.5 统计分析
使用SPSS 19.0软件对试验数据进行统计分析。其中,对捕食者体长、捕食量、耐冷性参数SCP和FP先采用Shapiro-Wilk法进行数据正态性检验,如符合正态分布且差异不显著则利用独立样本t检验法和Duncan氏新复极差法进行统计分析;否则使用非参数Mann-Whitney U和Kruskal-Wallis单因素ANOVA检验法进行统计分析。捕食功能反应模型的拟合程度分析采用F检验,干扰效应模型的分析采用卡方适合性检验。使用Origin 2019b软件作图。
2 结果与分析
2.1 隐翅甲和蜘蛛体长分析
经Shapiro-Wilk正态性检验,在22 ℃和12 ℃下,所有捕食者同一种类且同一虫态个体间的体长均呈正态性分布,且无显著性差异(0.776
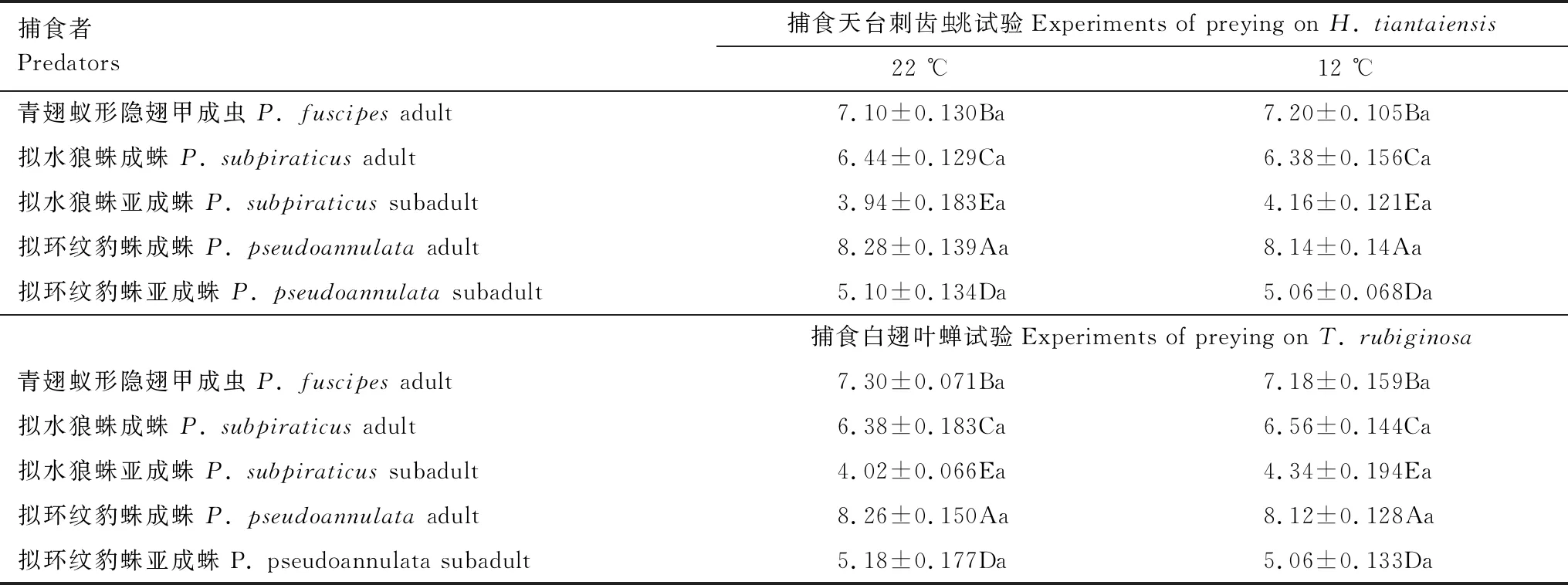
表1 两种温度条件下捕食天台刺齿虫兆和白翅叶蝉试验中隐翅甲和蜘蛛体长分析
2.2 不同温度下捕食性天敌对刺齿虫兆和叶蝉的捕食功能反应
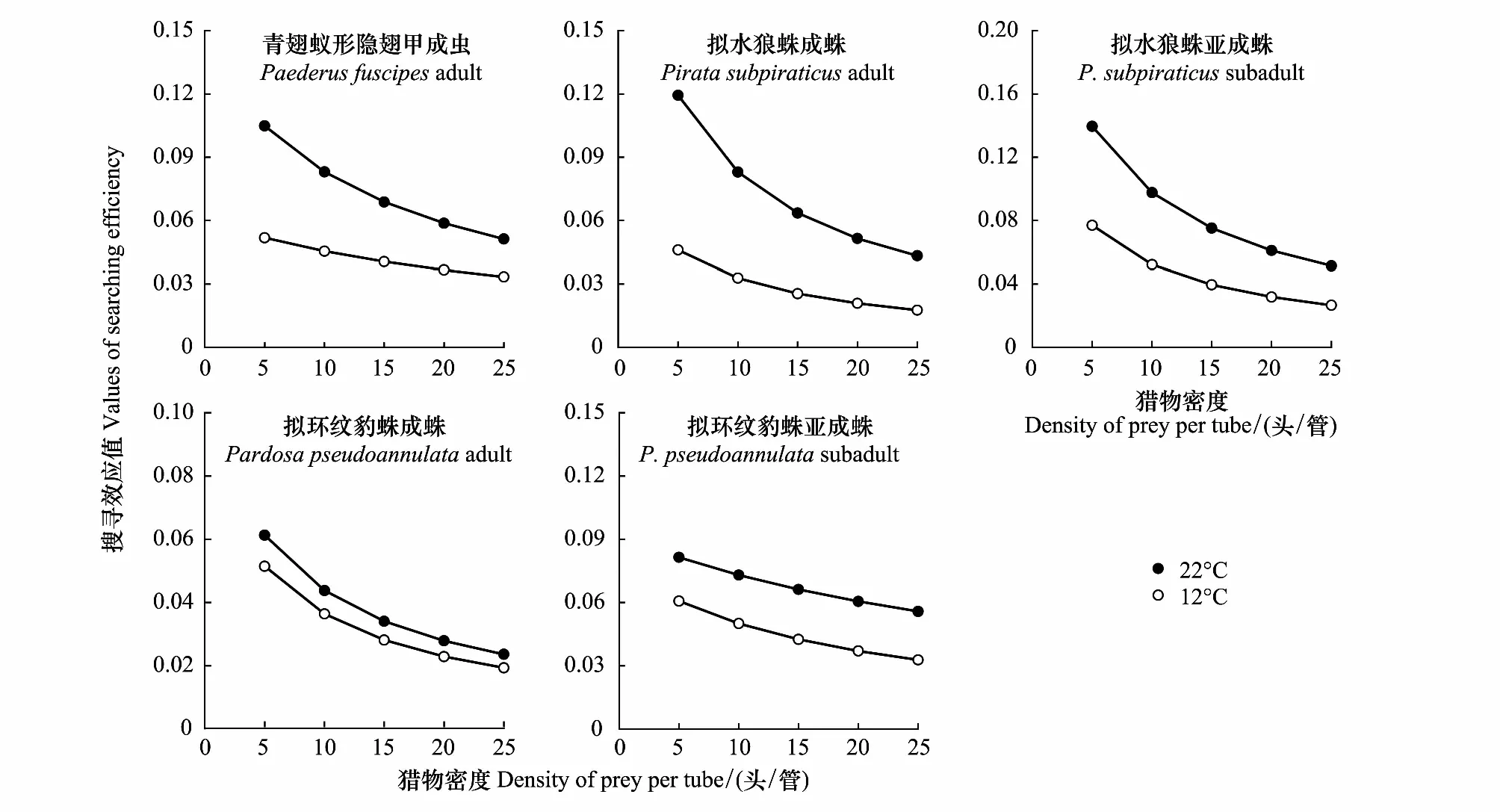
由图1和图2可知,在22 ℃和12 ℃下,随着这两种猎物密度的增加,所有捕食者对刺齿虫兆和叶蝉的捕食量均逐渐增加,捕食功能反应曲线呈上升趋势。
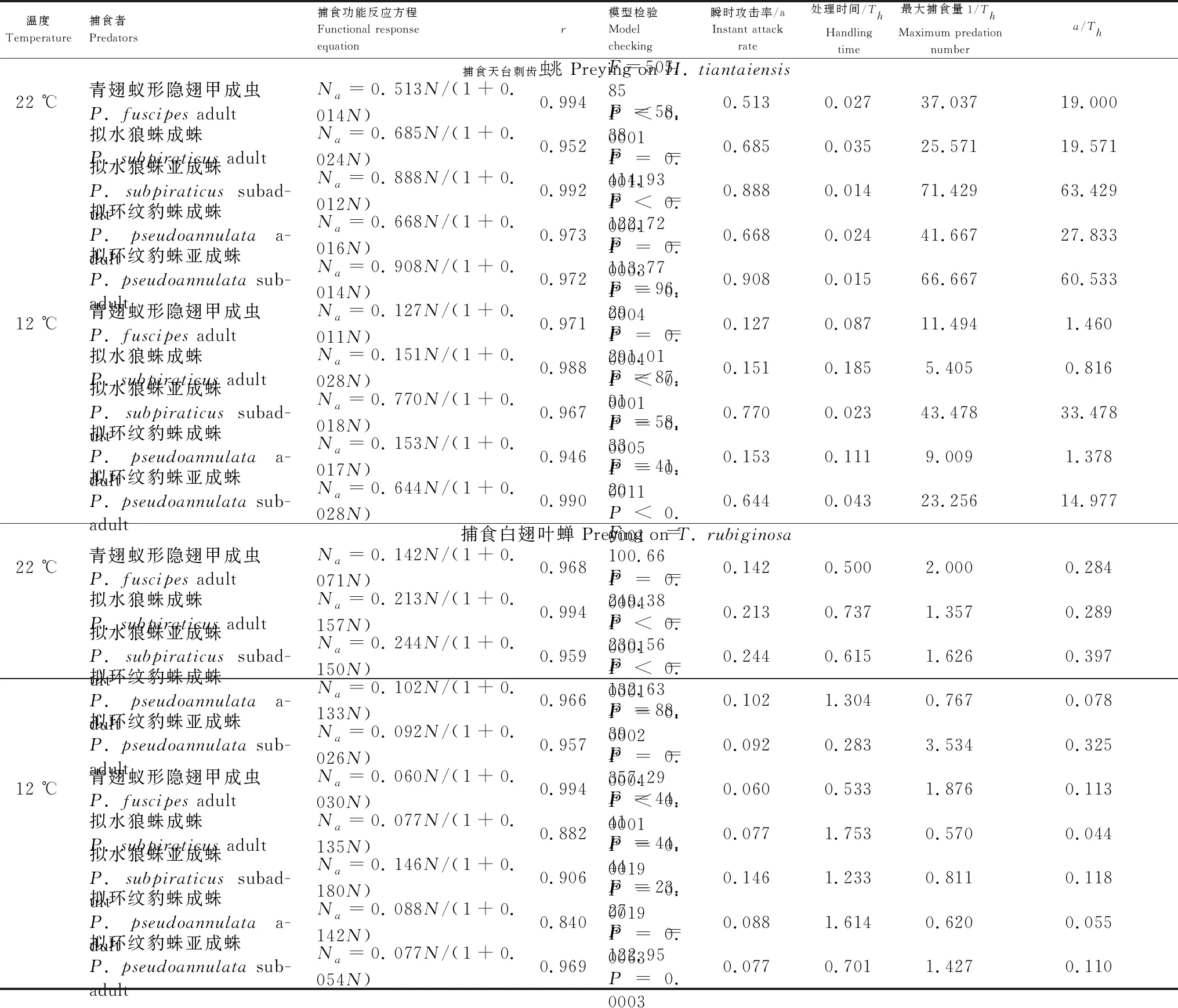
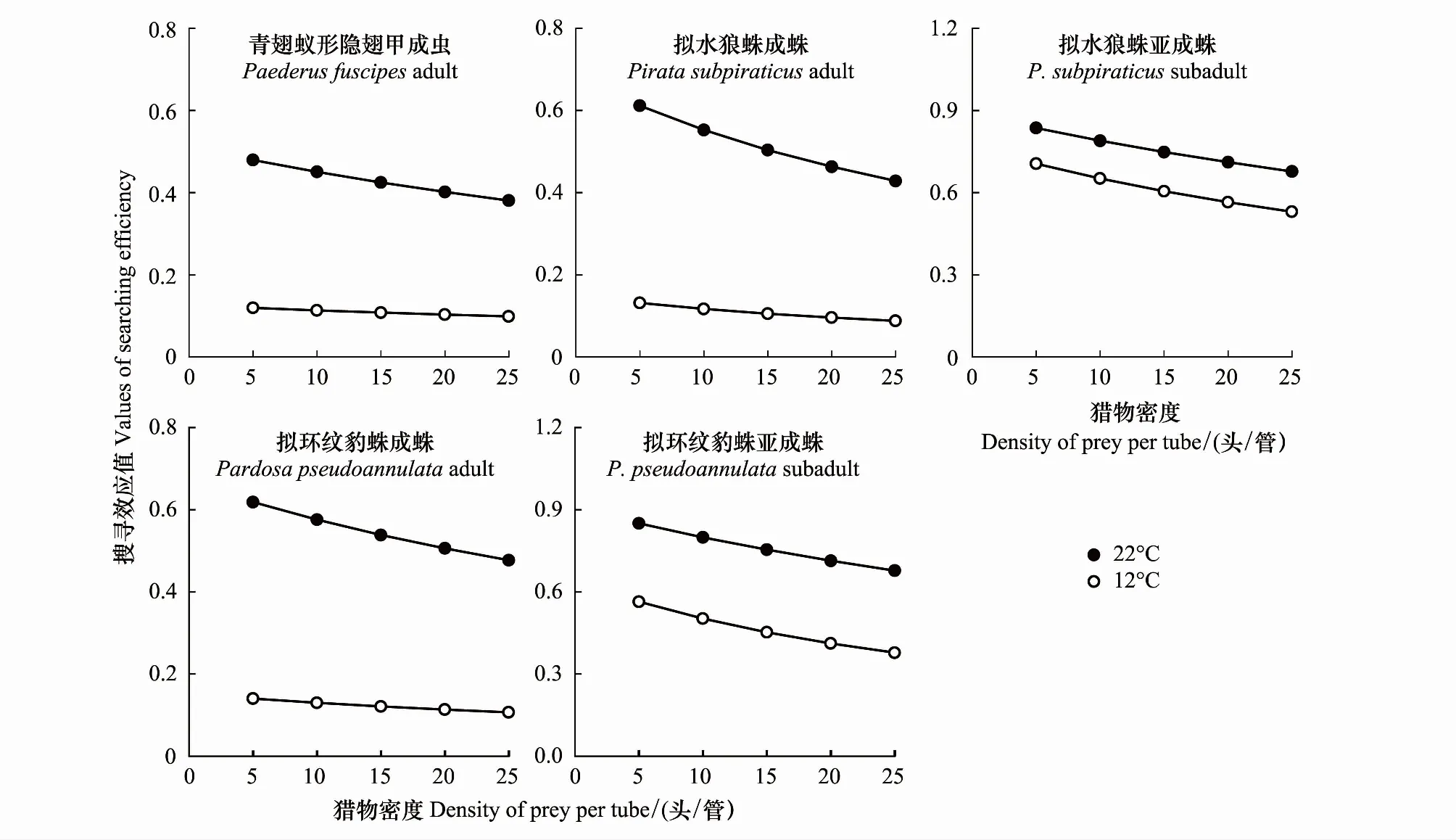
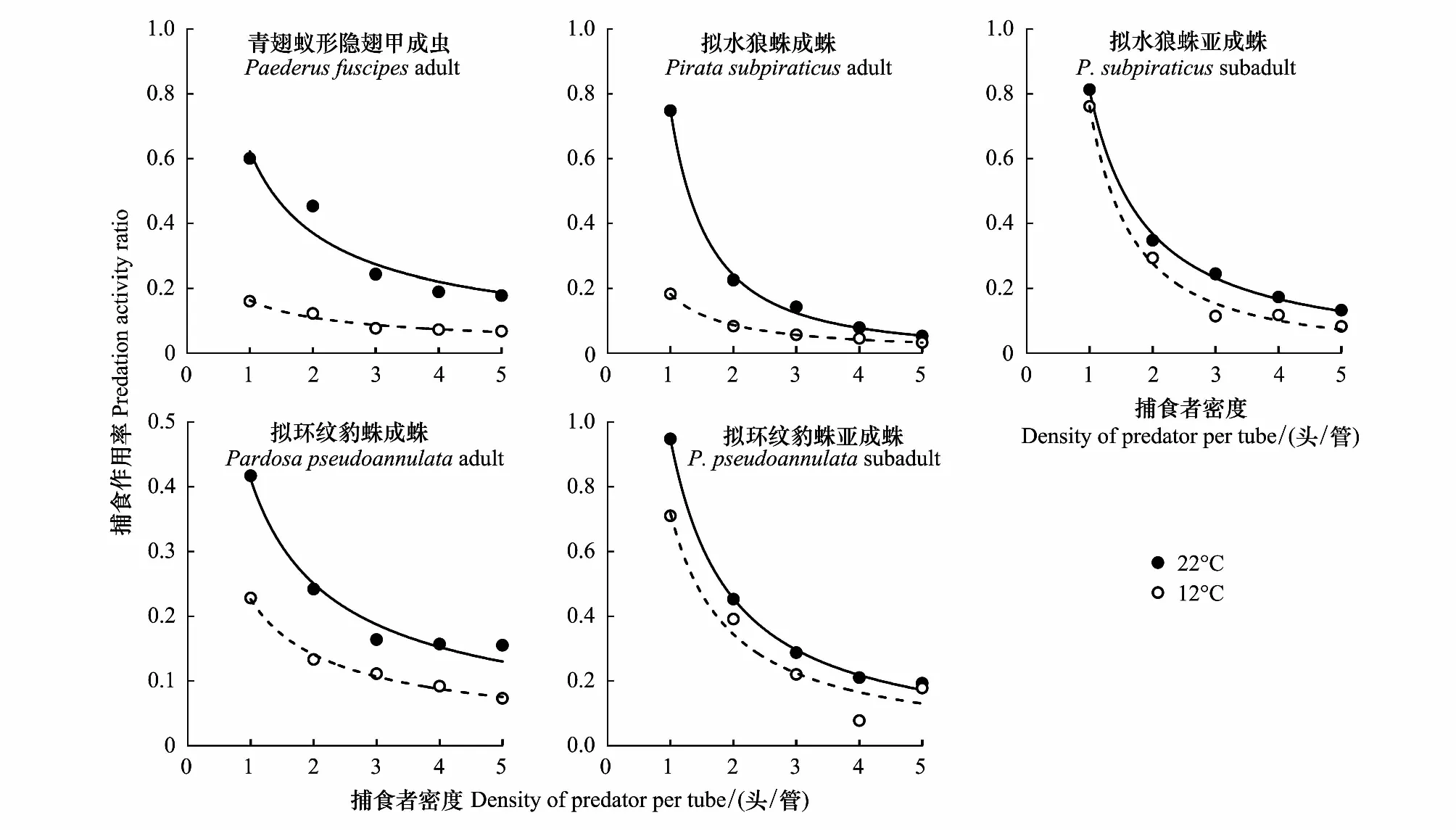
在捕食刺齿虫兆试验中,不同温度下捕食者对刺齿虫兆捕食量的变化规律明显不同。在22 ℃时,不同刺齿虫兆密度下同一捕食者的捕食量间均差异显著(3.79 图1 两种温度条件下隐翅甲和蜘蛛个体对天台刺齿虫兆的捕食功能反应曲线 在捕食叶蝉试验中,不同温度下捕食者对叶蝉捕食量的变化规律也明显不同。在22 ℃时,不同叶蝉密度下,除豹蛛成蛛外,同一捕食者的捕食量间均差异显著(9.60 图2 两种温度条件下隐翅甲和蜘蛛个体对白翅叶蝉成虫的捕食功能反应曲线 统计分析表明,这些捕食者的捕食功能反应模型拟合程度均达到了极显著水平(23.27 在捕食刺齿虫兆试验中,除了狼蛛亚成蛛外,22 ℃下其它捕食者的瞬时攻击率均明显高于12 ℃下的瞬时攻击率;而22 ℃下所有捕食者的处理时间均低于12 ℃下的处理时间;22 ℃下所有捕食者的最大捕食量和a/Th值均高于12 ℃下的最大捕食量和a/Th值;在22 ℃或12 ℃捕食刺齿虫兆试验中,豹蛛和狼蛛亚成蛛的瞬时攻击率、最大捕食量和a/Th值均明显高于其它捕食者,而它们的处理时间则明显低于其它捕食者的处理时间(表2)。 在捕食叶蝉试验中,除了豹蛛成蛛外,22 ℃下其它捕食者的瞬时攻击率均明显高于12 ℃下的瞬时攻击率;除了隐翅甲外,22 ℃下其它捕食者的处理时间均低于12 ℃下的处理时间;22 ℃下所有捕食者的最大捕食量和a/Th值均高于12 ℃下的最大捕食量和a/Th值。在22 ℃下,狼蛛亚成蛛的瞬时攻击率和豹蛛亚成蛛的最大捕食量,均明显高于其它捕食者的瞬时攻击率和最大捕食量,而豹蛛亚成蛛的处理时间和豹蛛成蛛的a/Th值均明显低于其它捕食者的处理时间和a/Th值。在12 ℃下,狼蛛亚成蛛的瞬时攻击率高于其它捕食者的值;隐翅甲的最大捕食量明显高于其它捕食者的最大捕食量;狼蛛和豹蛛成蛛的a/Th值相近,均明显低于其它捕食者的a/Th值(表2)。 另外,在22 ℃或12 ℃下,当猎物密度分别为20和25头/管时,所有捕食者对刺齿虫兆的捕食量均显著高于对叶蝉的捕食量(-3.19 表2 两种温度下隐翅甲和蜘蛛对天台刺齿虫兆和白翅叶蝉的捕食功能反应模型及参数值 从捕食者对刺齿虫兆和叶蝉的搜寻效应结果(图3、图4)可知,22 ℃下所有天敌的搜寻效应值(刺齿虫兆:0.381—0.850,叶蝉:0.024—0.139)均高于其12 ℃下的搜寻效应值(刺齿虫兆:0.089—0.706,叶蝉:0.018—0.046);两种温度下所有天敌对刺齿虫兆和叶蝉的搜寻效应均与猎物密度呈负相关,随猎物密度的增加搜寻效应曲线呈明显下降趋势。隐翅甲、狼蛛和豹蛛成蛛对刺齿虫兆的搜寻效应受温度影响明显,搜寻效应值从22 ℃下的0.5以上急剧下降到12 ℃下的0.1,下降了73.75%—79.21%;狼蛛和豹蛛亚成蛛对刺齿虫兆的搜寻效应受温度干扰小,22 ℃和12 ℃下的搜寻效应值均在0.35以上,温度变化导致的该效应值变化范围在0.3以内,下降了15.55%—44.02%。 图3 两种温度条件下隐翅甲和蜘蛛捕食天台刺齿虫兆的搜寻效应曲线 图4 两种温度条件下隐翅甲和蜘蛛捕食白翅叶蝉成虫的搜寻效应曲线 所有捕食者对叶蝉的搜寻效应随猎物密度的增加呈较为平缓下降趋势,特别是豹蛛成蛛的下降幅度最小。所有捕食者22 ℃时的搜寻效应值与12 ℃时的值相差较小,仅狼蛛成蛛和亚成蛛的值出现了明显变化,变化范围为0.024—0.073,下降了44.60%—61.34%;而隐翅甲、豹蛛成蛛和亚成蛛受温度变化影响最小,该值由22 ℃降为12 ℃时的变化范围为0.004—0.053,下降了16.80%—50.57%。 表3 两种温度下隐翅甲和蜘蛛自身密度对其捕食刺齿虫兆和叶蝉成虫的干扰效应模型及参数值 在22 ℃下,豹蛛和狼蛛亚成蛛对刺齿虫兆的搜寻常数值最高,其次是狼蛛成蛛、隐翅甲和豹蛛成蛛的值,搜寻常数最高值是最低值的2.3倍;干扰系数值以狼蛛成蛛的为最高,其次是狼蛛亚成蛛、豹蛛亚成蛛、隐翅甲和豹蛛成蛛的值,干扰系数最高值是最低值的2.3倍。在12 ℃下,狼蛛亚成蛛对刺齿虫兆的搜寻常数值最高,其次是豹蛛亚成蛛的值,而豹蛛成蛛、狼蛛成蛛和隐翅甲的值要明显低于前2者的值,搜寻常数最高值是最低值的4.7倍;干扰系数值以狼蛛亚成蛛的为最高,其次是豹蛛亚成蛛和狼蛛成蛛的值,而豹蛛成蛛和隐翅甲的值最小,干扰系数最高值是最低值的2.6倍(表3)。 在22 ℃下,狼蛛亚成蛛对叶蝉的搜寻常数值最高,其次是三者间接近的豹蛛亚成蛛、隐翅甲和狼蛛成蛛的值,最小的是豹蛛成蛛的值,搜寻常数最高值是最低值的3.5倍;干扰系数值以狼蛛亚成蛛的为最高,其次是豹蛛亚成蛛、狼蛛成蛛、隐翅甲和豹蛛成蛛的值,干扰系数最高值是最低值的2.5倍。在12 ℃下,豹蛛亚成蛛和隐翅甲对叶蝉的搜寻常数值最高,其次是狼蛛成蛛和亚成蛛、豹蛛成蛛的值,搜寻常数最高值是最低值的1.5倍;干扰系数值以隐翅甲的为最高,其次是豹蛛亚成蛛和成蛛的值,而狼蛛成蛛和亚成蛛的值最小,干扰系数最高值是最低值的1.9倍(表3)。 在两种温度下,所有捕食者对刺齿虫兆和叶蝉的捕食作用率均随捕食者自身密度的增加而下降(图5、图6)。其中,在22 ℃下,所有捕食者的捕食作用率下降幅度均较大;在12 ℃下,多数捕食者的捕食作用率下降幅度较大,但隐翅甲和狼蛛成蛛对刺齿虫兆、狼蛛亚成蛛对叶蝉的捕食作用率下降幅度较小。当捕食者密度为5头/管时,两种温度下所有捕食者自身密度对其捕食能力的干扰作用最大,对刺齿虫兆和叶蝉的捕食作用率均降至最低值,降幅为50.19%—92.65%,说明此时捕食者自身密度对其捕食能力存在较强的干扰作用(图5、图6) 图5 两种温度下隐翅甲和蜘蛛自身密度对其捕食天台刺齿虫兆的干扰效应曲线 图6 两种温度下隐翅甲和蜘蛛自身密度对其捕食白翅叶蝉成虫的干扰效应曲线 当猎物总数固定为20头时,以不同比例放入刺齿虫兆和叶蝉后所有捕食者在两种温度下的猎物选择性指数均大于1,表明这些捕食者在捕食刺齿虫兆和叶蝉时存在选择偏好性,总是更偏好捕食刺齿虫兆(表4)。同时,温度也显著影响了这些捕食者的捕食作用,不同温度下这些捕食者的猎物选择性指数变化规律明显不同。其中,在22 ℃下,10头叶蝉和10头刺齿虫兆处理中的豹蛛成蛛的选择性指数(28.00)要远高于其它所有处理中的捕食者的选择性指数(2.40—9.67);在12 ℃下,捕食者的选择性指数最高值(17.33)属于5头叶蝉和15头刺齿虫兆处理中的豹蛛亚成蛛,其次是该处理中的狼蛛亚成蛛选择性指数(15.00),而隐翅甲的选择性指数均较低(1.44—3.75)(表4)。 表4 两种温度条件下隐翅甲和蜘蛛对叶蝉和刺齿虫兆的捕食偏好性 在22 ℃和12 ℃快速冷驯化后,隐翅甲、狼蛛成蛛和亚成蛛、豹蛛成蛛和亚成蛛各自SCP和FP均呈规律性变化(图7)。其中,两种温度下的SCP均值依次为-9.2 ℃、-8.8 ℃、-6.4 ℃、-7.0 ℃、-7.9 ℃,而FP均值分别为-7.0 ℃、-5.9 ℃、-3.9 ℃、-4.7 ℃、-5.5 ℃。两种温度间除了狼蛛成蛛的SCP和豹蛛亚成蛛的FP显著差异外(t= -6.328,t=2.644;P<0.05),其它捕食者间的SCP和FP均无显著性差异(-1.884 在22 ℃和12 ℃下,这些捕食者间的SCP或FP均差异极显著(6.060 图7 隐翅甲和蜘蛛在两种温度快速冷驯化后的过冷却点SCP和结冰点FP 本文以冬水田休耕季节主要捕食性天敌青翅蚁形隐翅甲成虫、拟水狼蛛和拟环纹豹蛛成蛛、亚成蛛及它们的主要猎物天台刺齿虫兆和白翅叶蝉为代表,研究了温度变化对捕食者捕食猎物的影响,重点强调了近年重新鉴定和明确的天台刺齿虫兆在6—8个月非作物生育期低温条件下作为这些捕食者猎物的重要性。 本试验以冬水田休耕期优势捕食性天敌拟水狼蛛和拟环纹豹蛛两种常见的虫态以及青翅蚁形隐翅甲成虫为代表研究了它们的捕食作用。我们及其它相关研究指出,拟水狼蛛和拟环纹豹蛛均为冬水田或冬闲田休耕季节优势捕食者,均以成蛛、亚成蛛和幼蛛越冬[4,21—22,27];而冬水田休耕期或稻田生境越冬期青翅蚁形隐翅甲成虫的密度很大[4,20,28]。同时,结合李剑泉等[21]和王智[22]室内饲养拟水狼蛛和拟环纹豹蛛的研究结果可知,本研究供试的拟水狼蛛和拟环纹豹蛛亚成蛛分别约为4龄和5龄。由于捕食者体型或虫龄大小能显著影响它们对猎物的捕食量和捕食功能反应[12,14,29—30];而本研究结果表明,同一温度或不同温度条件下捕食者同一种且相同虫态个体的体长间均符合正态分布,且差异不显著,说明这些供试捕食者个体能够用于本试验中不同温度条件下的捕食作用比较试验。 本研究通过分析休耕季节冬水田2种蜘蛛和青翅蚁形隐翅甲对两种主要猎物的捕食作用,强调了天台刺齿虫兆等弹尾虫种类在该季节维持这些天敌种群发生的重要性。虽然白翅叶蝉是南方稻区水稻上的重要害虫之一[31],一些水稻生育期的研究分析了这3种天敌对白翅叶蝉的捕食作用[32],但尚无休耕季节冬水田白翅叶蝉的研究报道。迄今,尽管对稻田生态系统弹尾虫生态功能的研究较多,但因缺乏相关鉴定资料或对该类群分类的不重视,多数研究对它们的分类以目/纲级分类单元为主[33—35],而在种级分类水平上尚存疑问[18],甚至有错误现象[23,36]。近年来通过广泛调查发现,我国不同稻区地表弹尾虫的种类分布差异较大,但也存在一些发生量大且重要的广布种类,如我们重新鉴定的长角虫兆科中的2个种,即天台刺齿虫兆和纵纹刺齿虫兆H.social[4—5]。我们前期研究表明,这2种弹尾虫是不同种类蜘蛛的重要猎物,其中拟水狼蛛在冬水田休耕季节捕食纵纹刺齿虫兆的阳性检出率达到了69%[37];进一步利用定量实时聚合酶链反应(Quantitative real-time PCR)对冬水田不同种类蜘蛛捕食该刺齿虫兆的DNA量化比较结果显示,拟水狼蛛是更为活跃的该刺齿虫兆捕食者[5]。这里需要特别说明的是,这些弹尾虫种类在休耕季节稻田不应该仅仅被定义为捕食性天敌的替代猎物,而是捕食者的主要猎物;同时它们又能促进水稻等田间植物残体的分解,是稻田休耕期重建的节肢动物群落中的“关键或中心”节肢动物类群[4]。与上述水稻休耕季节冬水田弹尾虫的研究结果相比,在水稻生育期“水稻-害虫-天敌”三营养系统中它们的生态功能更多地被定义为“中性或无害”,仅仅作为优势捕食者蜘蛛等的随机选择性猎物,起到调控捕食性天敌拟水狼蛛等对主要害虫稻飞虱等捕食的作用[33—35,38]。另外,在水稻生态系统,鲜有研究青翅蚁形隐翅甲捕食弹尾虫的报道。 虽然捕食性天敌捕食作用研究的新方法和新技术在不断发展,如先后出现的酶联免疫方法[32,36,39]、稳定同位素技术[38]、分子技术[5,37]和视频录像分析技术[40]等,但依靠直接观察和计数的传统的捕食功能反应等技术和方法仍然在捕食性天敌捕食作用的研究中不可或缺[11,14]。另外,影响捕食性天敌捕食作用的非生物胁迫因素有很多,其中温度对捕食性天敌的捕食量影响较大,特别是低温条件[12—13,41—42]。由于狼蛛和豹蛛成蛛、亚成蛛及隐翅甲成虫在冬水田越冬期均有捕食活动,特别是冬水田休耕季节的前期(再生稻期)和后期(水稻移栽前期)[4—5];甚至稻田气温达5 ℃以上的晴天,蜘蛛也能出洞觅食[21]。因此,本研究利用捕食功能反应方法研究模拟条件下的冬水田季节温度变化对稻田主要捕食者捕食作用的影响有重要的现实意义。 本研究结果表明,在22 ℃和12 ℃下,所有捕食者对刺齿虫兆和叶蝉的捕食功能反应均符合Holling II反应模型,这些捕食者的捕食量与猎物密度极显著正相关。相关的研究表明,尽管捕食者对猎物不同龄期的功能反应模型存在差异[43],但通常单头捕食者的捕食功能反应符合Holling II反应模型[17—18,44—45]。在一定范围内,温度越高,捕食者的生理代谢就越快,捕食率也就越高[46]。尽管如此,我们发现温度降低没有影响狼蛛亚成蛛对猎物的瞬时攻击率和作用时间,相反却增强了它们的这种捕食反应特性,这可能反映了这些蜘蛛亚成蛛更强的抗低温适应能力和捕食作用。相关的报道也指出,与成蛛相比,拟水狼蛛若蛛捕食白背飞虱Sogatellafurcifera的能力要更强[47]。 另外,从本试验两种温度下所有捕食者对刺齿虫兆的瞬时攻击率、最大捕食量和a/Th值均相应地高于对叶蝉的这些参数值可知,刺齿虫兆是这些捕食者的主要嗜食猎物,而低温条件下隐翅甲相对于狼蛛和豹蛛而言也更嗜好捕食叶蝉。 搜寻效应是捕食者捕食猎物的行为反应结果。随猎物密度增加,搜寻效应则下降[14]。本研究结果表明,尽管在同一温度下,捕食性天敌对刺齿虫兆和叶蝉的搜寻效应值均随猎物密度增加而下降,但22 ℃下的搜寻效应值要高于12 ℃的值,且该搜寻效应值与刺齿虫兆密度的负相关程度要明显高于与叶蝉密度的负相关程度,这说明刺齿虫兆密度对该搜寻效应的影响更大;另外,隐翅甲成虫、狼蛛和豹蛛成蛛对刺齿虫兆的搜寻效应受温度影响明显,而对叶蝉的搜寻效应受温度影响不明显,这进一步说明了这些捕食者对两种猎物嗜食程度存在差异。相关研究指出,温度能显著影响捕食者对猎物的搜寻效应[43],且捕食者的这种搜寻效应与不同猎物种类有关[18]。 在捕食猎物时,隐翅甲、狼蛛和豹蛛自身密度会影响其捕食效率;其自身密度越大,干扰作用就越强,捕食作用率也会逐渐降低[44,48—49]。本研究结果表明,无论猎物是刺齿虫兆还是叶蝉,两种温度下这些捕食者随自身数量增多,其捕食作用率均下降,这种关系显然符合Hassell模型;且这些天敌的干扰效应方程均能很好地反映它们自身密度的干扰效应。但在两种温度和不同猎物条件下,该干扰效应方程的参数值出现了较大变化。在两种温度下,捕食者对刺齿虫兆的搜寻常数值和干扰系数值均以狼蛛和豹蛛亚成蛛的为最高;在22 ℃下它们对叶蝉的搜寻常数值和干扰系数值均以狼蛛亚成蛛的为最高,而在12 ℃下的这两个参数值却以豹蛛亚成蛛或隐翅甲成虫的为最高,这也反应了这些捕食者对温度和猎物种类变化的响应程度存在明显的差异。研究指出,温度变化能影响捕食者的干扰效应;同一捕食者密度下,随温度升高,捕食者的捕食作用率增大,其自身干扰效应就增强,干扰系数和搜寻常数会变大[50];而这显然也与它们对猎物的捕食选择性本身就存在差异密切相关[18]。 大量研究指出,青翅蚁形隐翅甲、拟水狼蛛和拟环纹豹蛛都是游猎型广食性捕食者,能捕食多种猎物[17,19,22,47,51—52]。尽管如此,本研究发现,这些捕食者对不同猎物的捕食显然存在选择偏好性,更嗜好捕食刺齿虫兆;这与庞保平等[18]的研究结果有一定相似性,即在长角虫兆Salinasp.和白背飞虱共存系统中,拟环纹豹蛛更嗜好捕食该长角虫兆。李剑泉和赵志模[53]研究发现,在不同猎物共存系统中,拟水狼蛛最喜欢捕食褐飞虱Nilaparvatalugens,其次是白背飞虱,对黑尾叶蝉Nephotettixnigropictus的捕食量很小。周集中和陈常铭[51]研究表明,相较于褐飞虱其他虫态,拟环纹豹蛛更嗜好捕食短翅成虫。Mishra等[54]研究指出,与稻缘蝽Leptocorisavaricornis和蚱蜢Oxyanitidula若虫相比,稻田优势种苏门答腊豹蛛Pardosasumatrana对黑尾叶蝉、稻纵卷叶螟Cnaphalocrocismedinalis和三化螟Scirpophagaincertulas成虫的捕食量更大。同时,本试验也发现,温度变化也对这些捕食者的猎物选择性指数产生了较大影响,但总的取食偏好性未发生改变。其它相关研究也指出,不同温度下环纹小肥螋Euborelliaannulipes总是对小菜蛾Plutellaxylostella幼虫具有捕食偏好性[55]。总之,研究冬水田休耕季节这些广食性捕食者对温度的适应性和对猎物的选择偏好性,有益于采取更加有效的天敌保护对策从而为翌年制定更加有效的稻田绿色防控措施打下坚实的理论参考依据。 昆虫是变温动物,对季节变化的自身温度调节能力直接影响着其种群的生存、繁殖和分布,特别是适应低温严寒的能力。因此,研究冬水田休耕季节隐翅甲、狼蛛和豹蛛的耐低温特性对保护它们的种群发生,尤其是捕食作用具有重要意义。由于捕食者的SCP和FP存在显著的季节性变化规律[15],为此,本研究模拟了冬水田休耕季节两种代表性温度条件,测定了它们诱导下的这些捕食者的SCP和FP,并结合其捕食作用,分析它们耐冷性参数与其捕食作用的关系。 本研究表明,通常两种温度间的各捕食者的SCP和FP差异不显著。但是,不同温度诱导下,隐翅甲、狼蛛与豹蛛之间的SCP和FP存在显著性差异;其中,隐翅甲的SCP和FP均低于狼蛛和豹蛛成、亚成蛛的值,这说明隐翅甲有更强的适应低温的能力。尽管如此,由于SCP和FP只是反映昆虫存活的低温下限,故本研究中没有发现它们与这些捕食者的捕食作用强弱有显著的正相关性;但在远高于它们SCP和FP的试验温度中,通常狼蛛和豹蛛亚成蛛的瞬时攻击率、最大捕食量和a/Th值都最大,表现出与它们的SCP和FP负相关的关系;而隐翅甲仅在12 ℃下与狼蛛和豹蛛亚成蛛一起表现出了较强的捕食叶蝉能力,这也可能与其本身较强的耐冷能力和嗜好捕捉叶蝉有关。我们相关的研究指出,在拟环纹豹蛛成蛛和青翅蚁形隐翅甲成虫共存系统中,隐翅甲具有更强的耐冷特性,在低温条件下其捕食行为表现的更为积极;尽管在22 ℃各处理条件下隐翅甲总是豹蛛的集团内猎物(单向性捕食),隐翅甲被捕食率为89%—100%,但在12 ℃相同各处理条件下豹蛛也常被隐翅甲作为集团内猎物捕食(双向性捕食),豹蛛被捕食率为10%—60%(作者待发表资料)。另外,我们也发现,相对于成蛛,温度降低后狼蛛和豹蛛亚成蛛的耐冷能力均增强,这也在一定程度上解释了为什么这些亚成蛛在低温条件下表现出了更强的捕食能力。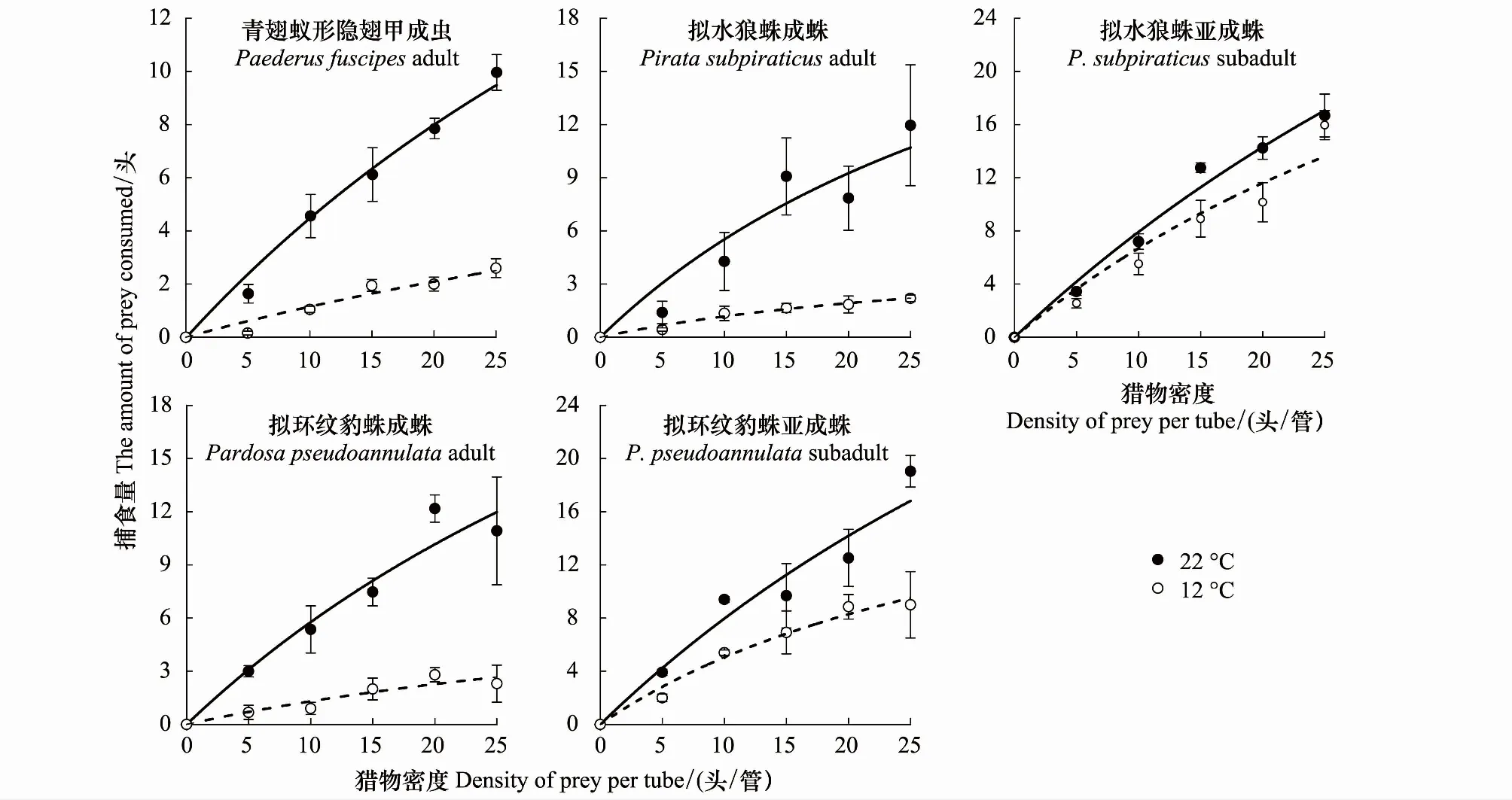

2.3 不同温度下捕食性天敌对刺齿虫兆和叶蝉的搜寻效应
2.4 不同温度下捕食性天敌自身密度对其捕食的干扰效应

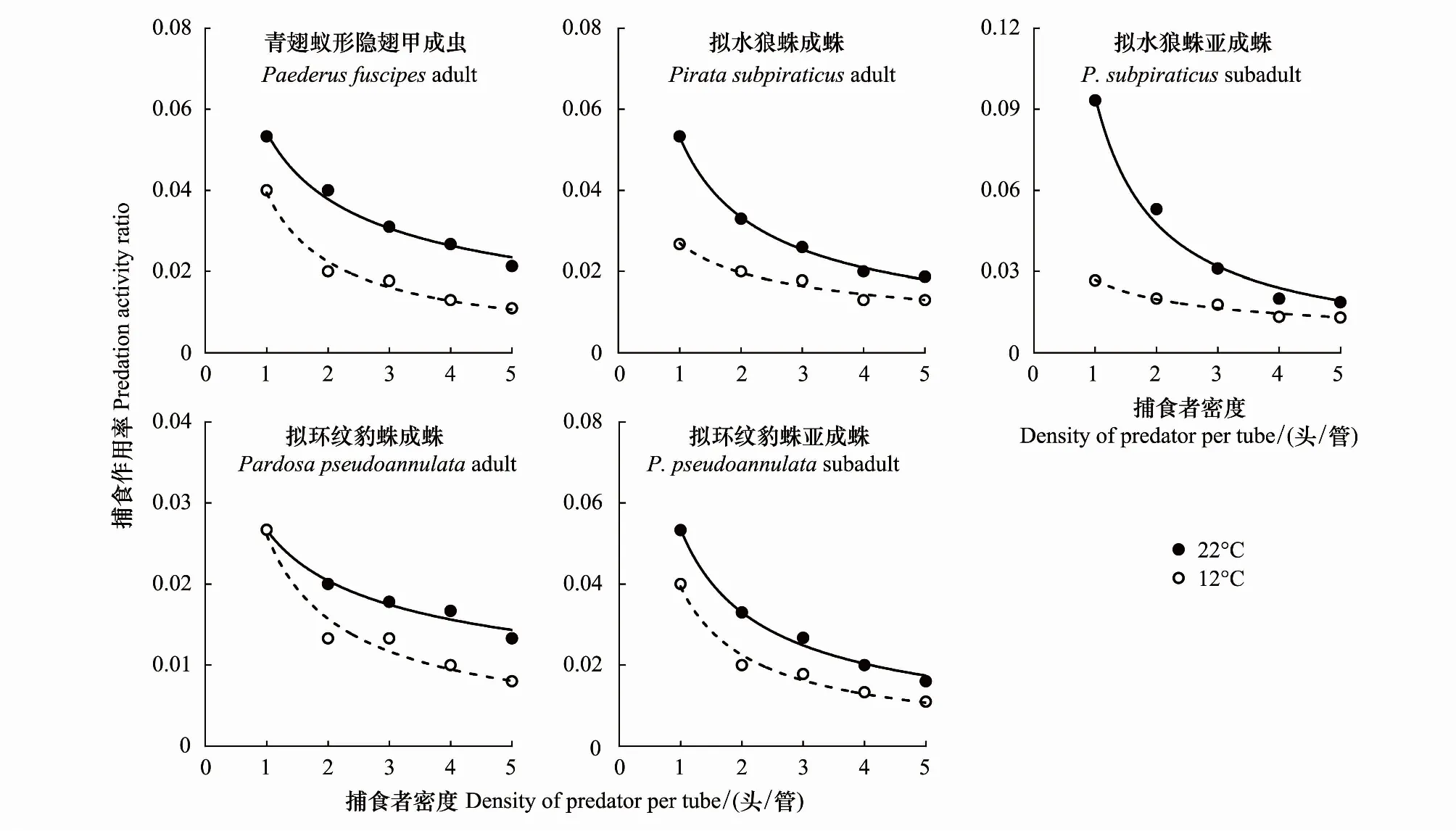
2.5 不同温度下捕食性天敌的捕食偏好性分析

2.6 不同温度下捕食性天敌的耐冷性分析
3 讨论
3.1 休耕期冬水田捕食性天敌的越冬虫态及猎物的生态功能分析
3.2 温度变化对捕食性天敌捕食功能反应的影响
3.3 温度变化对捕食性天敌搜寻效应及其自身密度干扰效应的影响
3.4 温度变化对捕食性天敌捕食偏好性的影响
3.5 捕食性天敌的耐冷性与其捕食作用关系
