湿热河谷芭蕉群落优势物种空间分布格局与竞争对称性研究
2022-04-25张昊楠周大庆钱者东刘邦友
张昊楠,周大庆,钱者东,陈 皓,曹 晓,梁 盛,刘邦友,高 军 ,*
1 生态环境部南京环境科学研究所, 南京 210042
2 贵州习水国家级自然保护区管理局, 遵义 564600
3 贵州赤水桫椤国家级自然保护区管理局, 赤水 564704
植物是固着的自养型生物体,其空间分布格局的形成与自身群落的构建方式(繁殖、扩散等)和相邻个体间对有限的资源(光照、水份等)的竞争关系联系紧密[1—3]。在森林生态系统中,植物个体对群落内有限的资源的竞争关系如何在演替中塑造其空间分布格局一直是植物群落学、生态学领域的重要科学问题[4—8]。光照是郁闭森林中最为有限的资源之一[9—10],而同一森林群落内对同一资源处于竞争状态的植物个体间并不会平均的分配资源,个体较大(胸径、树高等)的植株获得光资源的比例可能会更高,而个体较小植株则多处于竞争劣势,进而形成不对称竞争过程[9,11]。反之,个体差异小的植物个体间则可能处于对称竞争过程。总的来说,植物竞争的对称性理论从不同个体资源获取策略最大化的角度来阐述个体之间竞争关系,能够更好的理解在自然资源有限的情况下植物个体间分配资源的方式,这一重要竞争机制与生物多样性维持、生物入侵等热点问题联系紧密,具有较高的研究价值[9—11]。
植物个体之间对光照的不对称竞争被认为是塑造森林植物群落空间结构和物种共存的主要驱动力之一[9—14]。随着森林的演替,植物个体的空间分布格局通常会从聚集分布转变为均匀分布或者随机分布,而不对称竞争过程导致的负密度制约调控机制往往在这一过程中发挥重要作用[1,6,9]。例如,当光照透过林冠层照射进森林时,高大的植物会遮荫矮小的植物。对光照的不对称竞争过程将对矮小个体的生长产生负面影响,抑制其直径和高度的增长,甚至导致其密度依赖性死亡率的上升和空间格局均匀分布[11]。然而,同样的过程会对更高(和更大)个体的生长产生积极影响,提高这些个体及其种群的生存力[9]。在这一过程中,不对称竞争会导致相邻植物个体呈现出成对的优势树和受抑制树,受抑植物个体大小通常小于优势树且死亡率更高[9,15],而两者之间的种间关联往往会表现为显著的负相关关系[16]。
检测相邻植物个体的大小差异在空间上的分布模式是研究森林群落中植物个体对的光竞争对称性的重要手段之一[9,12]。例如,采用标记变异系数分析相邻树木个体之间的大小差异[2,5,15],可以精确测定森林中相邻的同种或异种个体之间由于密度效应而引起的胸径、树高等植物个体属性在生长量上的降低[7,17—18],探明不同植物个体间或种群内的光竞争对称性[12]。当相邻树木为异种个体时,双变量标记变异系数分析也可以用来探索不同物种之间潜在的种间竞争关系[19]。
不对称竞争过程是导致植物物种生物多样性丧失的重要机制之一[20—23],对其开展研究不仅对于揭示物种对自身空间分布格局的形成和群落构建模式具有重要意义,也有助于解释群落物种多样性变化的内在驱动力及森林生态系统稳定性。本研究在贵州赤水地区湿热河谷植物群落中设置大型固定样地进行调查,使用基于距离和胸径标记的空间点格局与变异系数分析,对芭蕉群落优势种的空间分布格局和竞争对称性开展研究,旨在探明不对称竞争和对称竞争过程如何塑造湿热河谷植物群落优势种的空间分布格局,阐述驱动其占据空间、成为群落优势种的空间策略和种间竞争机制等重要生态学过程。主要包括以下内容:(1)赤水湿热河谷芭蕉群落中优势植物的空间分布格局特点。(2)优势种的种内竞争对称性。(3)优势种的种间竞争对称性。(4)优势种的种间关联关系。
1 材料与方法
1.1 研究区概况
本研究区域位于贵州大娄山脉北支的赤水河畔,样地设置在贵州赤水桫椤国家级自然保护区内。该区域地处贵州高原北部向四川盆地的过渡地带,地貌上具有典型“V”形河谷特点,尽管其在气候带上属于中亚热带湿润季风气候区,但海拔600 m以下的河谷小气候较为特殊。研究区域大于10℃活动积温为4200—5100℃,年降水量1200—1300 mm,相对湿度常年较高,为86%—90%。本区域土壤发育受到母质和河谷小气候影响,主要为紫色土,土层较深且呈现出中性或微酸性。长期稳定的水热条件以及较大的海拔垂直差异产生了研究区域内独特的植被类型垂直地带性变化[24]。研究区域植被演替的顶极群落类型是亚热带常绿阔叶林,但在研究区域内海拔700 m以下的沟谷地带分布有大量的南亚热带、热带雨林层片,其多雾阴湿环境形成了湿热河谷雨林植被群落。历史上这一区域受到人为活动的破坏较大,正处于次生森林向顶级植被类型的演替恢复过程中[24—25]。芭蕉在本植被区分布范围较广,是本区特色之一[24—25]。
1.2 研究方法
1.2.1样地设置和调查
本研究样地建设在赤水河一级支流金沙河河谷南侧,受赤水河谷狭长分布、两侧低山坡度过陡等客观因素限制,样地建设尺寸为240 m × 40 m的长方形,样地内主要植物群落为赤水地区典型的湿热河谷芭蕉群落,平均海拔567 m,共记录到1989株胸径大于1 cm的植物个体(表1,图1)。样地内共记录到79种木本植物和大型草本,其中优势植物物种主要包括芭蕉(Musasp.)、粗糠柴(M.philippensis)、粗叶木(L.chinensis),川钓樟(L.pulcherrima)和红果黄肉楠(A.cupularis)等5种。通过克隆繁殖的方式扩散的芭蕉种群数量最大(表1,图1),样地内的群落外貌也主要由芭蕉来表现,在颜色表现为浅绿色并掺杂少许深绿色小块斑,季相变化不明显,林冠形状也较为单一。由于该区域低海拔河谷地带早年间受到人类活动影响较大,样地内的植物群落次生性较强,各种植物物种在保护区建设后正处于逐步恢复的状态,因此样地内植物的胸径和植株高度整体均较小,平均值分别为8.77 cm和4.98 m(表1,图1)。
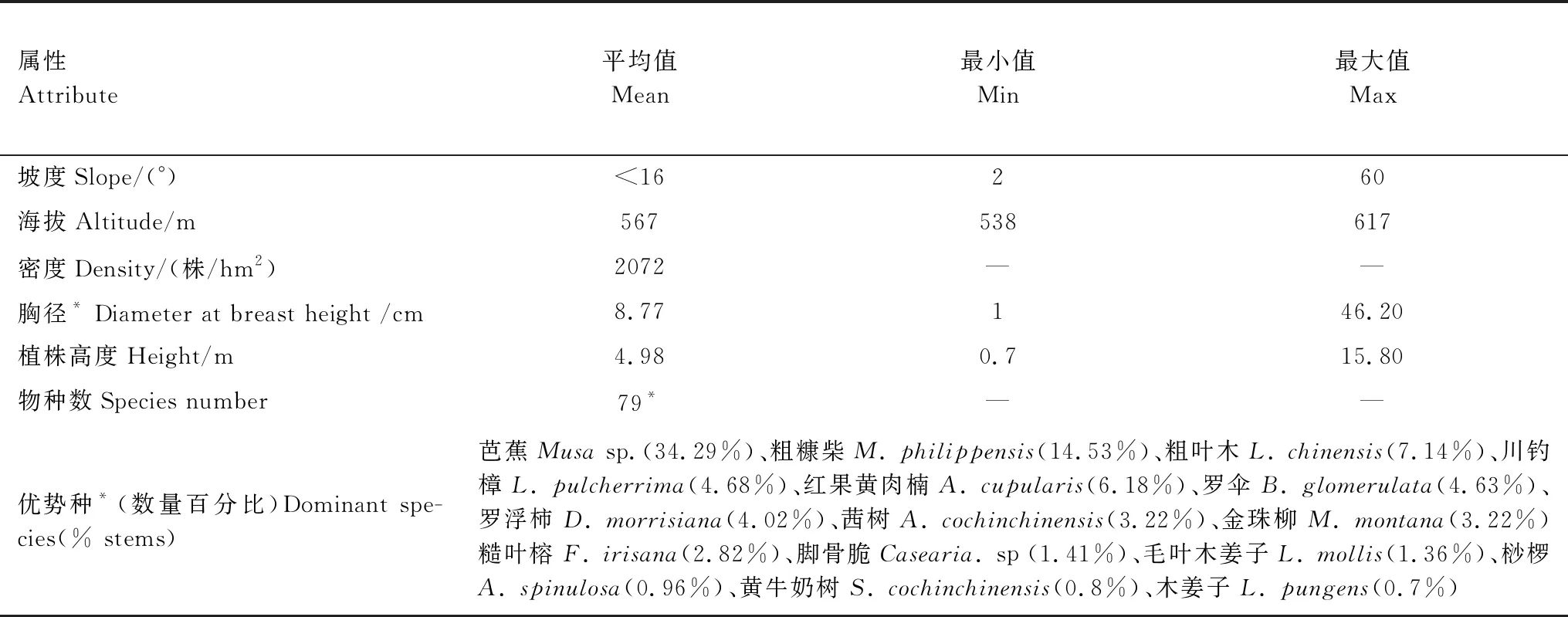
表1 湿热河谷雨林样地群落基本属性
样地建设和调查按照《生物多样性观测技术导则—陆生维管植物》(HJ 710.1—2014)规范,并参照巴拿马Barro Colorado Island(BCI)50 hm2热带雨林样地建设技术规范和经验,用森林罗盘确定样地的方向和基线,然后用经纬仪(全站仪)将样地划分成20 m × 20 m样方;记录过程中测量点之间的水平距、斜距和高差;对每个样方的顶点编号并永久标记;最后,用卷尺、测绳或便携式激光测距仪将每个20 m × 20 m样方划分为5 m × 5 m小样方,样方顶点用临时PVC管标记,边界用塑料绳或其他材料临时标记,之后在每个样方顶点处安置固定样桩,采用直径15 cm的不锈钢管,打入地面深度15至20 cm。将5 m × 5 m样方作为胸径(Diameter at Breast Height: DBH)≥ 1 cm乔木和灌木的基本观测单元。在观测单元内逐一记录所有个体的植物物种,测量树高(植株高度)、胸径等植物个体的生长属性并挂牌标识,样地内群落空间分布格局见图1。群落内物种的重要值按下式计算:重要值=(相对密度+相对优势度+相对频度)/ 300。其中,相对密度=(某一种的个体数/全部种的个体总数)× 100;相对优势度=(某一种的基面积之和/全部种的基面积之和)× 100;相对频度=(某一种的频度/全部种的频度之和)× 100。植物种群的植株密度按下式计算:密度=种群密度/样方内某种植物的个体数。
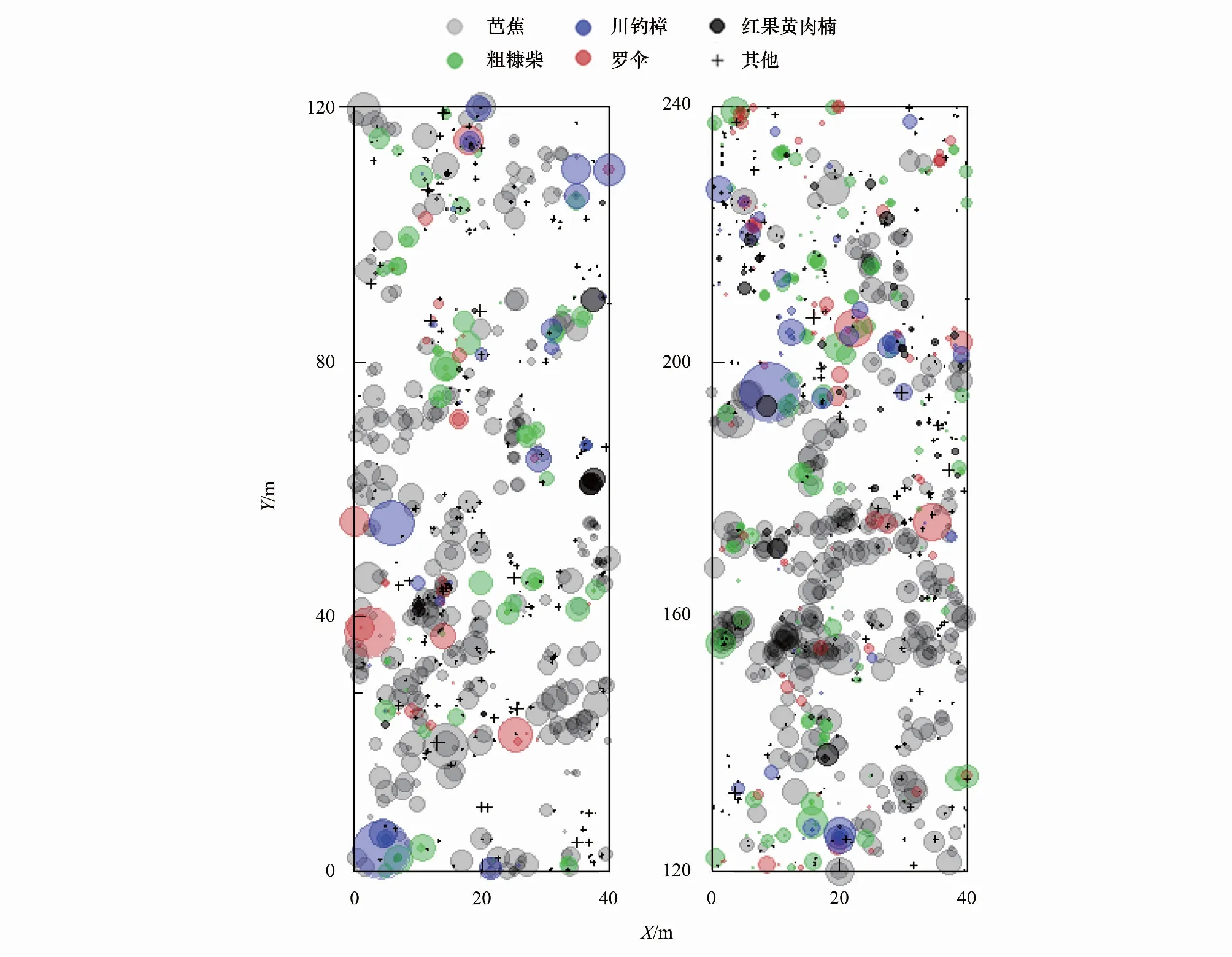
图1 湿热河谷雨林大样地内胸径大于1 cm的植物个体空间分布示意图
1.2.2 统计方法
(1)空间点格局分析和种间关联分析(pair-correlation function, PCF)
使用对数相关函数对植物个体的空间点格局和种间关联进行分析,该函数是一个概率密度函数,其在运算取样过程中使用圆环代替了早期的Ripley′s K函数中半径为r 的圆形,避免了积累效应[5]。该函数计算公示如下[17]:
对于同一个种群内的同种个体即种内竞争,采用空间点格局分析(即单变量对数函数分析,Univariate pair-correlation function: Uni-PCF)方法g11(r)来进行;对于不同种群中的不同种个体的空间关联性即种间竞争,采用种间关联分析(即双变量对数函数分析,Bivariate pair-correlation function: Bi-PCF)方法g12(r)来计算,通过计算以样地内不同种群内不同树种个体1在以自身为圆心、半径为r的圆环区域内另一个树种个体2的分布数量来分析植物个体1和2的相关关系[7],其中个体1和2是群落中不同树种的两个任意个体。在本研究的单变量分析和双变量分析中,若g(r)=1,则同一种群中植物群落的分布格局为随机分布或者距离为r的不同种群中的物种1和物种2的个体在空间上无相关关系;若g(r)>1,则同一种群中同种植物个体空间分布格局聚集性分布或不同种群中的不同个体呈现种间正关联;若g(r)<1,为均匀分布或种间负关联[7]。
(2)竞争对称性分析(mark variogram function, MVF)
本研究以群落中树木个体的胸径(DBH)作为标记属性,采用单变量标记点格局分析(Univariate mark variogram function: Uni-MVF)和双变量标记点格局分析(Univariate mark variogram function: Bi-MVF)方法研究赤水河谷群落内植物优势种个体标记(胸径)大小在空间分布上的自相关关系,使用标记变异分析γmm(r)函数进行计算,其结果可以用来检测相邻树木个体在生长量(胸径、冠幅或树高)上的不对称竞争(asymmetric competition)[7,26—27]。对于同一个种群内的同种个体即种内竞争对称性,采用单变量标记点格局分析(Uni-MVF)方法即γmm来计算;对于不同种群中的不同种个体的种间竞争对称性,采用双变量标记点格局分析(Bi-MVF)方法γ(m1,m2)来计算,计算公式如下[9]:
对于胸径单变量和双变量标记点格局分析,当γ(m1,m2)(r)=1时,距离为r的物种1和2的胸径之间无相关关系;当γ(m1,m2)(r)>1时为正相关关系,表明距离为r的两植物个体1和2的标记属性的大小的差异大于整个群落的平均水平,即为不对称竞争关系;当γ(m1,m2)(r)<1时为负相关关系,即距离为r的两植物个体1和2的标记属性的大小的差异低于整个群落的平均水平,其竞争过程表现为对称性[9]。
(3)零模型选取与显著性检验
根据不同科学问题,本研究共使用了3种零模型(null model)对概率密度分析结果的显著性进行检验。针对不同空间分析选取恰当的零模型十分重要,若选取不恰当零模型来进行Monte Carlo拟合往往会导致对空间格局错误的解释[28]。因此,本研究中在空间统计分析中,针对空间点格局分析、种间关联分析和竞争对称性分析,分别使用了3种不同的零模型,具体如下:一是为了消除空间异质性对实验结果的影响,在基于距离的空间点格局分析(即单变量对数函数分析,Uni-PCF)中使用异质泊松过程作为零模型(heterogeneous poisson, PH)来进行Monte Carlo拟合以形成置信区间[6,29];二是在种间关联分析中(即双变量对数函数分析,Bi-PCF),研究样地内不同树种进入群落演替的时间并不一致,繁殖方式(种子繁殖、克隆繁殖)也不一样,是相互独立的过程,因此采用种群独立零模型(population independence,PI)进行拟合[5,30];三是在竞争对称性分析中(即单变量和双变量标记变异系数分析,Uni-MVF和Bi-MVF),将群落中每一颗树木的胸径随机标记在树木个体的空间位置上,通过Monte Carlo方法不断拟合以形成树群落中胸径标记潜在的空间结构的置信区间,直到显示出标记中潜在的空间结构,故采用完全随机独立标记零模型(independent marking,IM)[26]。
本研究中Monte Carlo 拟合检验采用Getzin S和Martin Svátek等的方法[1,26],用199次Monte Carlo 拟合中5个最高和最低的拟合结果来构建95%置信区间。使用空间统计学软件programita、R studio软件进行统计分析和作图,其中R studio软件所使用的程序包主要包括spatstat、ads等[31—32]。
2 结果与分析
2.1 优势群落结构特征和空间分布格局
研究发现,湿热河谷大样地群落中优势种优势度明显,芭蕉的重要值占整个群落比例为33.6%,其在密度、胸径和重要值上均明显大于其他物种(表2)。通过对胸径和树高(植株高度)的统计分析发现,芭蕉的平均胸径在5种优势种群最大(11.86 cm),其次是川钓樟(9.83 cm),而粗糠柴、粗叶木和红果黄肉楠则相对较低(表2);此外,研究样地内群落物种垂直分层不明显,高度差异不大,木本植物尤其是乔木树种平均树高中最大值仅为5.98 m(川钓樟),与大型草本植物芭蕉的植株高度(5.15 m)差异不大(表2)。这表明尽管研究区域的地带性植被虽然是亚热带常绿阔叶林,但是在本研究中设立的湿热河谷样地中,形成了以大型草本芭蕉为主的湿热河谷灌草群落,群落内植物个体高度较低。
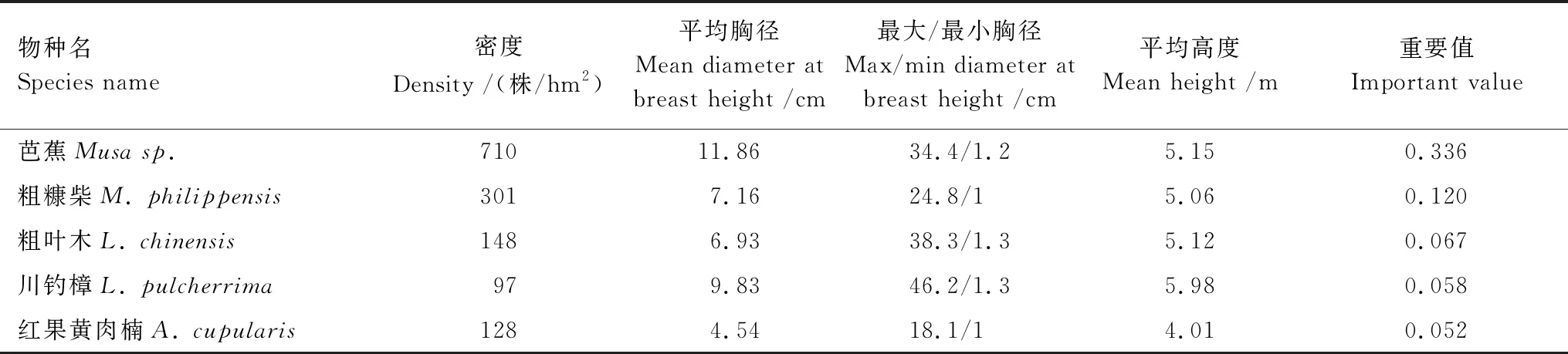
表2 湿热河谷雨林样地内重要值前5位优势种的数量特征
在基于距离的单变量对数函数(Uni-PCF)分析中,5种优势种的空间分布格局全部呈现出显著的小尺度下的聚集分布(aggregated),并且表现为随着距离的增加,聚集度逐渐下降的特点,最终呈现出完全随机分布(random)。其中,芭蕉在0—5 m的尺度呈现出聚集分布的格局,随着距离的增加,聚集度逐渐下降,在大于13 m的尺度上呈现出完全随机的分布。粗糠柴和粗叶木在0—2 m的尺度下呈现聚集分布。川钓樟、红果黄肉楠等径级个体较大物种在0—1 m的尺度下呈现出聚集分布的格局(图2)。
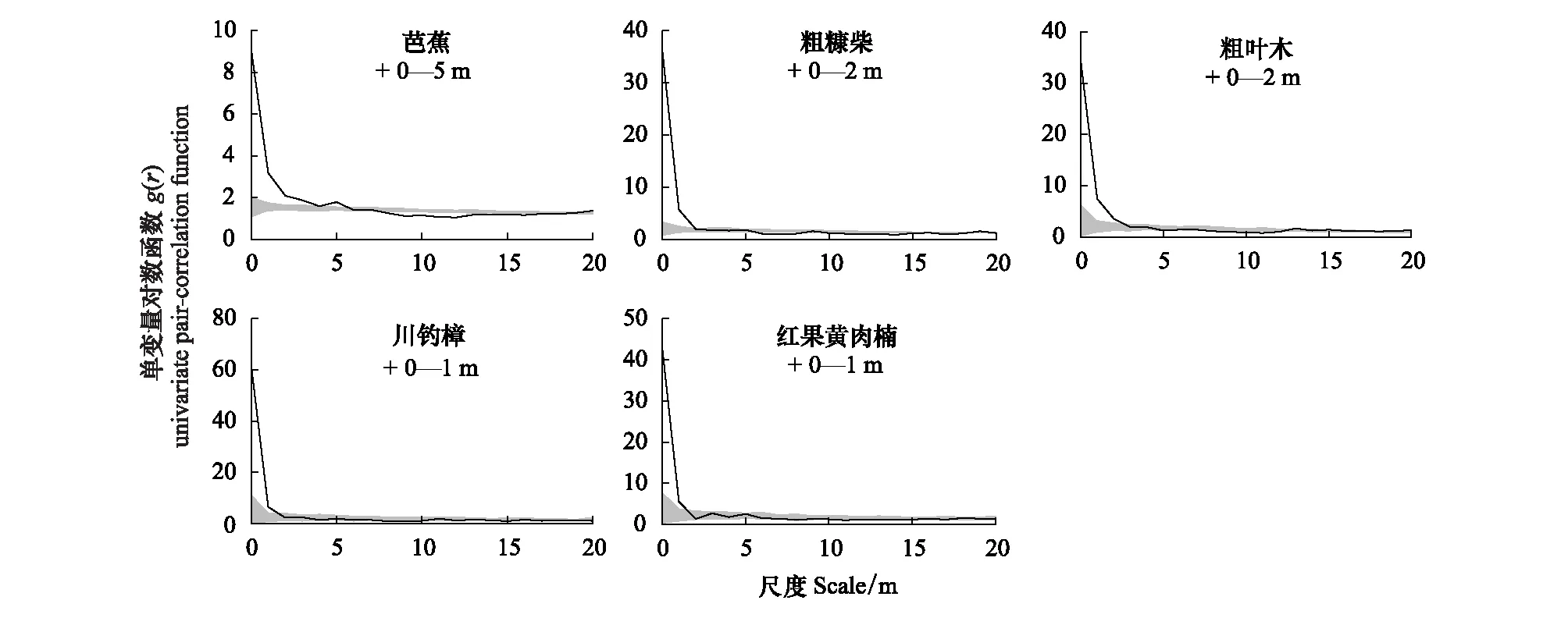
图2 样地内5位优势物种单变量对数函数分析(空间分布格局)
2.2 优势种种内竞争对称性分析(Uni-MVF)
在基于植物个体胸径大小的单变量变异系数分析(Uni-MVF)中,芭蕉种群在0—5 m的尺度下呈现出显著小于置信区间的下包迹线(对称性竞争),这说明距离较近的芭蕉个体大小(胸径)的差异要显著小于群落内的平均水平,表现为显著的种内对称竞争关系。与之相反,粗糠柴等其余4种优势种均在小尺度下表现为种内不对称竞争的特点(γmm>1),但均未高于置信区间的上包迹线,结果并不显著(图3)。
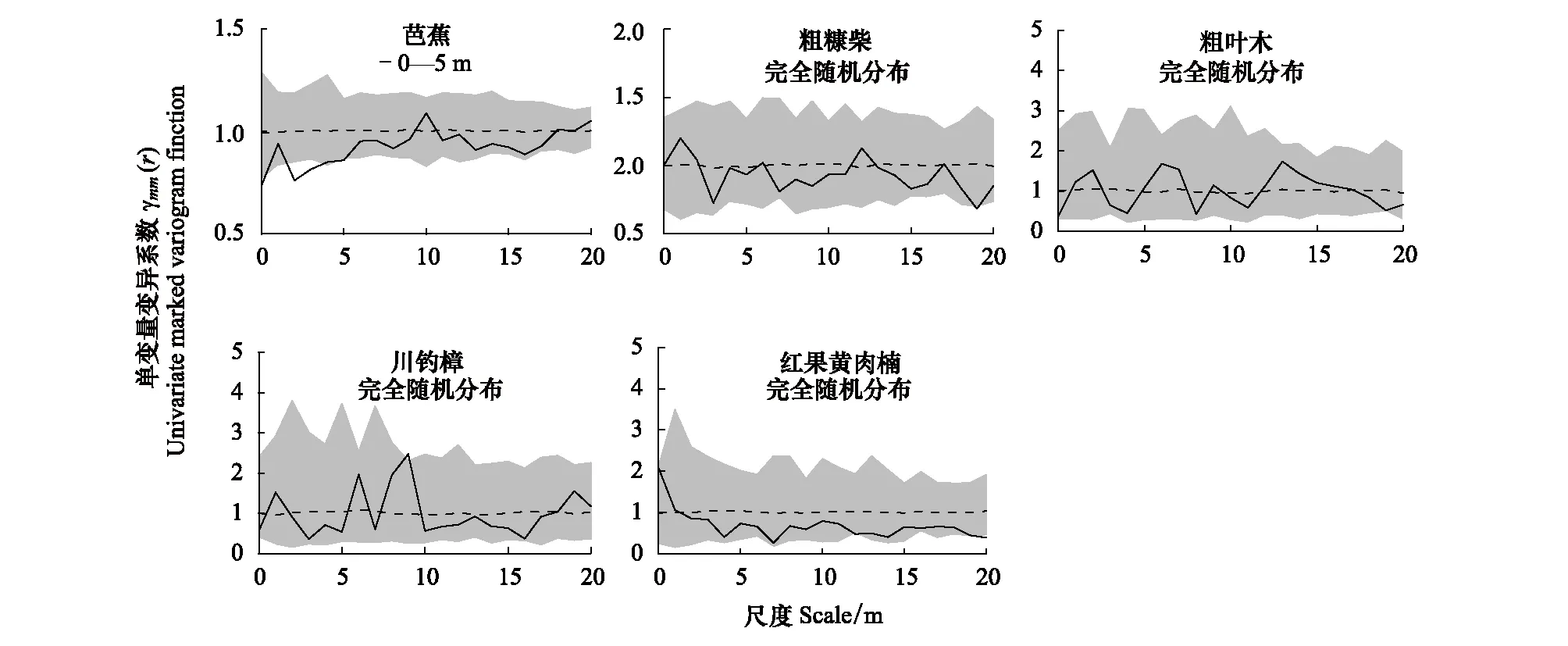
图3 样地内5种优势物种的单变量胸径标记变异系数分析(种内竞争对称性)
结合2.1中PCF分析的结果,尽管5种优势树种均呈现出在小尺度下种内聚集分布的特点,但芭蕉种群种内聚集分布的个体大小差异很小,表现为显著的种内对称性竞争。其他4种优势种的种内聚集表现为不显著的种内不对称竞争过程。
2.3 优势种种间竞争对称性分析(Bi-MVF)
采用基于植物个体胸径大小的双变量标记变异系数(Bi-MVF)分析了重要值排名前5位优势种之间的竞争对称性(共计10对物种)。研究结果发现,湿热河谷样地内的5种优势种种群间的竞争以不显著的对称性竞争关系为主,仅有芭蕉和粗糠柴两种在5 m的小尺度上呈现出显著的不对称竞争关系(图4),这说明距离较近的芭蕉和粗糠柴的胸径大小的差异要明显高于群落内的平均水平,这可能是由于距离较近的树种个体间对(水、阳光)等资源的竞争会导致生长量的下降。此外,芭蕉和粗叶木、川钓樟和红果黄肉楠等另外3种优势树种也呈现出小尺度的不对称竞争关系,但结果并不显著。其余4种树种组成的6对竞争对称性分析则主要呈现出小尺度下的对称竞争关系(γm1m2<1,图4),尽管结果并不显著。
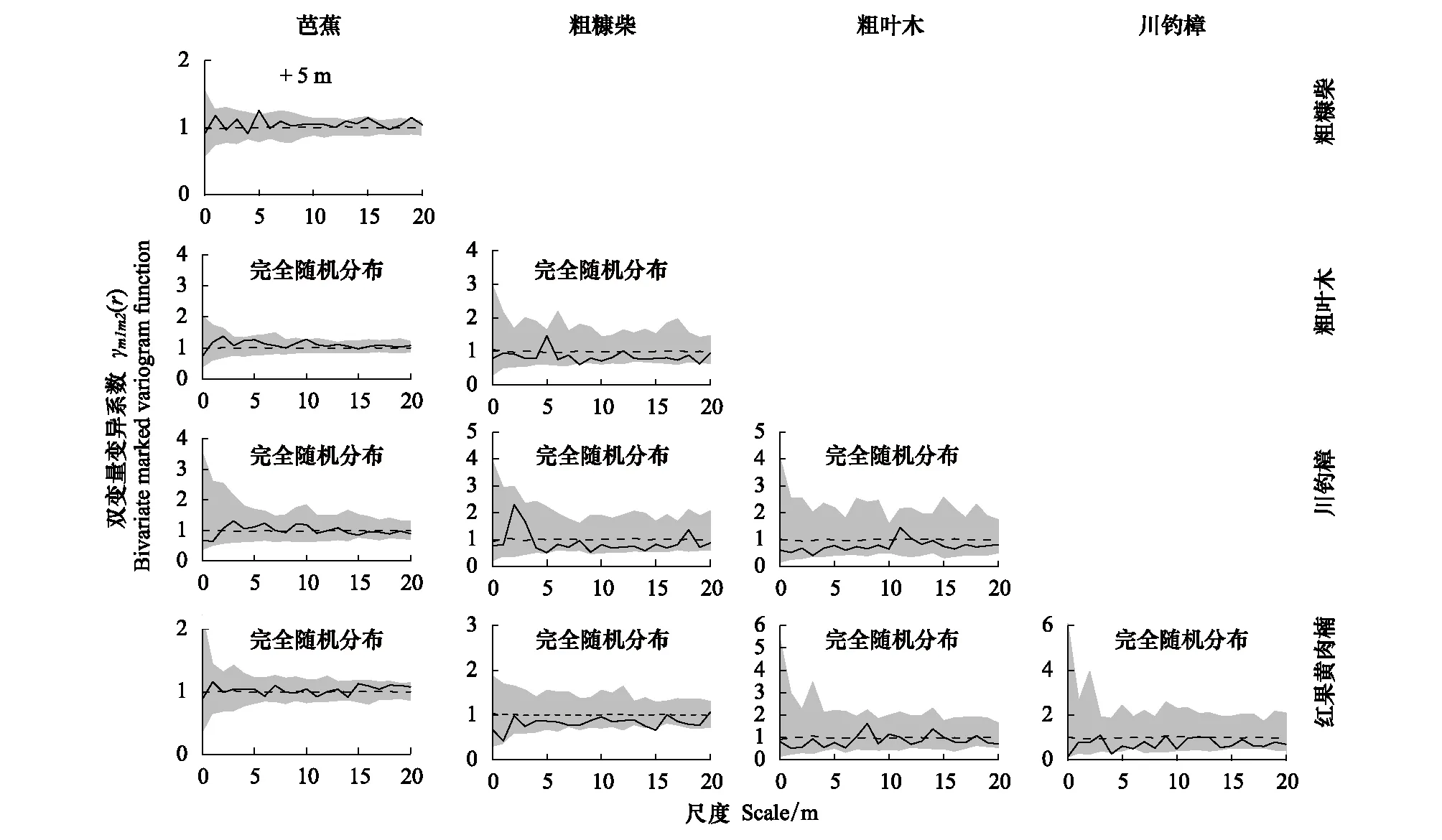
图4 样地内5种优势物种的双变量胸径标记变异系数分析(种间竞争对称性)
2.4 优势树种种间关联分析
双变量点格局分析(Bi-PCF)主要检测了重要值排名前5的群落优势种两两对应的10对树种的空间关联与竞争关系。结果表明,群落优势种芭蕉和粗糠柴在6 m的尺度下低于置信区间下边界,这说明芭蕉与粗糠柴种群中树木个体在这一尺度呈现出均匀分布的格局,即为显著的种间负关联。群落优势种粗叶木和粗糠柴、川钓樟在1 m的尺度下出现了显著正相关关系,粗叶木和红果黄肉楠在0—2 m的尺度下也出现了微弱的显著正相关关系,而粗糠柴和川钓樟在9—10 m的尺度下出现了显著正相关关系(图5)。这表明本研究中5种优势种之间的种间负关联关系主要出现在芭蕉和其他4种优势种之间(仅1对显著),其余优势种之间的种间关联主要表现为显著正相关和无相关关系。结合2.3中优势种种间竞争对称性分析结果,芭蕉与粗糠柴之间的不对称竞争关系可能是导致两者在种间关联中呈现显著负相关的主要原因。
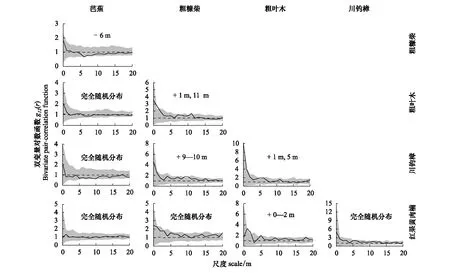
图5 样地内5种优势物种的双变量对数函数分析(种间关联)
3 讨论
3.1 优势树种空间分布格局
植物个体空间分布的聚集程度和斑块性,对植物种如何利用自然资源、作为资源而被使用以及其繁殖、扩散等生物学研究起到了至关重要的作用[3]。由于植物个体本身不具可移动性,其自身群落的构建过程对其形成特定的空间分布格局贡献很大,根据植物个体在自然更新中扩散方式的不同,也可能产生不同空间分布格局[2—3]。目前,对种子植物通过有性繁殖方式更新扩散的空间点格局研究已经非常之多[33—35],其空间分布主要与果实类型、大小和传播方式有关,如壳斗科坚果体积大且籽粒密实,受到重力作用,种子在母树周围聚集,由种子萌发产生的幼苗或幼树在小尺度上表现为聚集分布[33]。然而,在热带、亚热带和温带地区,以克隆繁殖的作为主要繁殖方式的物种有很多[1,36—37]。在本研究中,单变量点格局分析发现主要优势种芭蕉种群在0—5 m的尺度下呈现出显著的空间聚集分布,其的空间聚集度也远大其它优势种。这种空间分布的特点可能与本研究中芭蕉属植物主要采用无性克隆繁殖的方式进行扩散有关,其营养繁殖能力很强,地下走茎较为发达,在条件适宜的情况下扩散主要通过地下茎出芽形成分株实现[1,36],而更新产生的分株往往聚集母株距离较近,因而相比于群落内其他采用种子繁殖的物种,芭蕉在空间格局分布上表现为较强的聚集分布特性。
此外,尽管本研究中芭蕉种群的数量在湿热河谷群落中处于优势地位,且个体成长十分迅速,但并未形成长期、稳定的单优群落,这可能是由于随着原生群落的演替和发展,川钓樟等乔木植物逐渐占据群落上层空间,使喜阳性的野芭蕉获取的阳光等资源逐渐减少,这可能会导致部分芭蕉因得不到充足的阳光和其他资源,而在竞争中处于劣势;另一方面,芭蕉作为一种大型草本植物并无发达的木质化构造,地面上主干在成长到一定高度下会因难以承受本身的重量而倒伏[38—39]。
3.2 优势种种内竞争对称性
在森林的演替中,种内聚集可能会导致相邻植物个体之间表现为不对称竞争过程,密度效应会引起的胸径、树高等植物个体属性在生长量上的降低,当竞争强度足够时,也可能会引发植物个体的空间均匀分布和自疏现象[7,9,12]。而本研究中的结果则有所不同,虽然种内聚集度较小的粗糠柴等4种优势树种呈现出种内不对称竞争的趋势,但种内聚集最大的芭蕉种群则表现为显著的种内对称性竞争。这种不同优势种在竞争对称性上的差异说明了植物种群构建自身的方式(繁殖、扩散等)也是其形成特定空间分布格局和竞争关系重要影响因素之一[3,5]。相比于有性繁殖过程,采用无性克隆繁殖的植物个体在扩散过程中的优势往往更大。在条件适宜的情况下,一些生长快、繁殖能力强的物种甚至可以在较短时间内形成单优群落[23,40—41]。本研究中,相比于粗糠柴、川钓樟等优势种的有性繁殖过程,芭蕉种群主要采用无性克隆分株繁殖且生长较快[38—39]。在森林演替中,出芽生殖的繁殖方式和较快的生长速度,将会减少同种负密度制约对种内聚集个生长的影响,从而引导其种内竞争关系向对称性竞争转变,进而降低种内聚集导致的密度依赖性死亡率[23]。另一方面,粗糠柴等4种优势种的种内不对称竞争尚未引起明显的自疏从而导致均匀分布。
总的来说,芭蕉种群克隆繁殖和种内竞争在个体大小上的对称性推动了其种内聚集分布格局的形成(种内聚集度大于其余4种优势种),减少了群落内部的自疏,增加了该种群在本地区湿热河谷群落内的生存力,是其能够在湿热河谷群落内占据优势,成为群落中优势度最大物种的重要原因之一。
3.3 优势种种间竞争对称性与种间关联
尽管植物种类繁多,不同个体之间在形态、生理和分布上差异很大,但是其生存所需要的资源种类几乎相同(光照、水份、土壤养分等),植物个体在生长发育、繁殖扩散过程中对这些有限资源的争夺,会导致不同种个体间呈现相互竞争的过程,当这种竞争关系为不对称的过程时,生长速度快、生存能力强的物种可能会处于不对称竞争过程中优势地位[9,42]。当竞争强度足够时,异种密度效应不仅导致个体生长量(胸径等)的下降,也会引发种间关系呈现出负关联的趋势[6,43]。
本研究的结果显然支持了这种观点,即芭蕉和粗糠柴等4种优势物种之间的竞争关系呈现出显著的不对称性和负关联的特点。在这一竞争机制下,相邻的芭蕉和粗糠柴个体间呈现出在个体大小上(胸径)的显著差异,而个体较大且生长速到较快的芭蕉种群将在不对称竞争过程占据优势,并通过对周边相对矮小的粗糠柴等物种的遮阴作用抑制其胸径和树高的增长[15,26],进而导致了相邻的异种个体间密度依赖性死亡率的上升[11],最终引起了芭蕉和粗糠柴个体之间分布趋向于均匀化,在种间关联分析中呈现出显著的负相关关系。此外,芭蕉与其余3种优势种之间的不对称竞争关系和种间关联也呈现出类似的机制,尽管并不显著。
因此,芭蕉和其他优势种之间的不对称竞争过程是本研究中森林群落次生演替主要驱动力之一,也是芭蕉种群在湿热河谷群落中占据主要空间、形成优势的重要机制之一。与本研究结果类似,张光明和唐建维等的研究结果也报道了芭蕉种群在退化生态系统恢复与重建季节性雨林过程中的关键作用,并同样认为野芭蕉的个体数量和优势度将随着演替的进行更趋明显,最终形成物种丰富、结构复杂且相对稳定的次生林生态系统[36—39]。
4 结论和展望
本研究在赤水地区湿热河谷植物群落中设置大型固定样地进行调查,从空间分布格局和竞争对称性的角度来探索了以5种优势植物物种如何在亚热带森林次生演替中塑造其空间分布格局的生态学过程。结果表明,相比较采用种子繁殖方式的4种优势树种,以无性克隆繁殖为主要扩散方式的芭蕉种群的空间分布在种内聚集程度上更大,其种内竞争呈现出较为明显的对称性;在5种优势种之间的种间关联和种间对称性上,芭蕉与其他优势种主要表现为不对称竞争和种间负关联关系。根据研究结果,我们认为:(1)芭蕉种群种内竞争的对称性推动了其种内聚集分布格局的形成,减少了种群内部的负密度依赖死亡与自疏作用,增加了该种群在本地区湿热河谷群落内的生存力,而芭蕉和其余4种优势种之间的不对称竞争过程和种间负关联进一步增加了其在群落中的竞争力,是其能够在湿热河谷群落内占据优势,成为群落中优势度最大物种的重要机制之一。(2)不同优势种在种间竞争对称性上的差异说明了植物种群构建自身的方式是其形成特定空间分布格局和竞争关系重要影响因素之一,相比于有性繁殖过程,采用无性克隆繁殖的植物个体在扩散过程中的优势往往更大,条件适宜的情况下,一些生长快、繁殖能力强的物种可能会通过与相邻的其他物种间的不对称竞争过程来提升种群获取有限资源的能力,成为种间不对称竞争过程中的优势个体。
我国亚热带地区长期受人为活动的影响,致使部分地区丧失了植物赖以生存的原有生境[44—46],尤以低海拔地区影响最为严重[47]。大量受破坏的森林处在次生演替恢复的早期过程中,一些先锋物种进入演替后对植物多样性和物种共存关系产生了巨大的影响,例如近年来多位学者报道了关于克隆植物(如毛竹等)对原生森林生物多样性、群落次生演替和次生森林恢复产生了负面影响[20,23,40—41],而竞争关系的对称性可能是决定着次生森林能否恢复至原生状态的关键生态学过程之一[20—23]。因此,建议今后要在加强克隆植物空间分布格局和竞争关系的相关基础研究,深入理解不对称竞争过程与次生演替和生物多样性丧失之间的关系,相关理论基础也可对决策者今后制定植物物种多样性保护政策提供科学参考。
