杉木林改造成乡土阔叶林对林下植物物种组成和多样性的影响
2022-04-25谭许脉莫雪青高冠女尤业明明安刚黄雪蔓
谭许脉,张 文,肖 纳,莫雪青,高冠女,尤业明,3,明安刚,黄雪蔓,3,*
1 广西大学林学院,广西森林生态与保育重点实验室,南宁 530004
2 中国林业科学研究院热带林业实验中心,凭祥 532600
3 广西友谊关森林生态系统国家定位观测研究站,凭祥 532600
杉木(Cunninghamialanceolata(Lamb.) Hook)由于生长快、抗虫性强和材质优良等特点,在我国亚热带地区有悠久的广泛种植历史,是我国特有的优良针叶树种,也是我国重要的森林资源之一[1]。根据数据统计结果显示,杉木在中国的种植面积已超过1000万hm2,约占我国人工林总面积的26.5%和世界人工林总面积的5%[2—6],为我国经济的发展和满足人们对木材的迫切需求作出了巨大贡献。然而,有研究发现长期的杉木纯林种植,尤其是连栽经营极易产生土壤酸化、持水能力下降、肥力急剧退化和病虫害增加等一系列生态环境问题,最终导致杉木人工林的生产力严重下降[7—10]。
随着改革开放的深入,我国经济得到了飞速发展,人们生活水平得到了极大提升,加大了社会对中、高档珍贵木材的需求。但由于长期的大量肆意砍伐,导致天然的珍贵阔叶林资源锐减,我国每年需要花费大量的外汇从东南亚、非洲和美洲等地进口大径级珍贵的阔叶材来满足社会的需求[11—13]。因此,在我国非常适合珍贵阔叶树种生长的亚热带地区,越来越多的杉木林被改造成乡土阔叶树种,这不仅能够提供高价值木材满足社会的需求和增强我国木材产品在国际上的竞争力,而且能够提升生态系统服务功能,包括改善土壤质量和增加森林生物多样性等[14—15]。红锥(CastanopsishystrixMiq.)和米老排(MytilarialaosensisLec.)这两种乡土阔叶树种不仅材质优良,也有着发达的根系和丰富的林下凋落物,对于涵养水源、改善土壤肥力和防止水土流失等方面均具有重要作用,是目前亚热带地区杉木林改造中最为常用的经济用材树种[16]。
林下植被是保证人工林生物多样性和生态功能可持续性的重要指标之一[17—20],在人工林可持续经营发展中有着至关重要的作用,譬如在维持生态系统结构和功能的完整、促进系统物质循环和能量流动、涵养水源、保持水土、促进种子萌发和幼苗生长等方面均起着重要作用[20],保护林下植被是维持其生态系统多功能性的重要手段之一。了解并掌握人工林的经营和管理措施对林下植物群落的物种组成和多样性的影响,将在一定程度上有助于填补人工林树种单一化种植的缺陷,有利于维持人工林生态系统的稳定和减缓土壤肥力的衰退。杉木林改造成红锥林和米老排林,不仅改变了群落的冠层结构,而且其凋落物数量和质量的改变也将对土壤的理化性质产生不同程度的影响[21],但这些改变对人工林林下灌木层和草本层植物物种组成及其多样性的影响及其机制我们还知之甚少。本研究对在南亚热带杉木林采伐迹地上重新种植的杉木林、红锥林和米老排林的林下植被进行调查研究,以探明杉木林改造成乡土阔叶树种后其林下灌木层和草本层植物物种组成和多样性的变化及其主要影响因素,为科学经营管理人工林和提高人工林生态系统多功能性提供科学依据。
1 材料与方法
1.1 研究区域及样地概况
调查研究选在广西壮族自治区凭祥市广西友谊关森林生态系统定位观测研究站(106°51′ E—106°53′ E, 22°02′ N—22°04′ N)内开展。由于受海洋季风和大陆季风双重作用的影响,该区域属于典型的亚热带季风气候,其特点是夏季高温多雨,冬季温暖少雨;年均温约21℃,年均降雨量约为1400 mm,年均蒸发量约为1300 mm,相对湿度约为82%。其地形以山地丘陵为主,土壤类型以花岗岩风化后形成的红壤为主[22]。
本研究选取在1991年皆伐的杉木林迹地上重新种植的杉木林(对照,CK)、红锥林和米老排林作为研究对象,每个树种分别构建5块20 m×20 m的独立标准样地,同种林分的每块样地之间距离不少于200 m。在造林初期进行间伐抚育,之后没有再进行人工干扰,林下植被发育充分。
1.2 样地调查和样品采集分析
于2014年7月对研究样地内的植物群落进行全面调查。将每块20 m×20 m 的样地划分为4个10 m×10 m样方,调查并记录每个样方内乔木的胸径、树高和冠幅等;在10 m×10 m 样方的左下角设置一个5 m×5 m的小样方,并按照植物多样性调查方法,记录每个小样方灌木层和草本层植物的种名、高度、株数和盖度等。采用森林罗盘仪(DQL- 1B)测定样地的坡向和坡度,采用GPS测定样地的经、纬度和海拔,样地乔木层的冠层透射系数采用冠层图像分析仪(CI- 110)进行测定。在4个10 m×10 m 样方中随机选择其中3 个,在其右下角分别设置2 m×2 m 的小样方收集凋落物,置于烘箱中烘干(65℃)至恒重并采用Vario EL III 元素分析仪测定其C、N含量。
在每个20 m×20 m样地内,以距离样地中心5 m,在0、45、90、135、180、225、270、315°方向上分别布设8个采样点,用土钻(内径=5 cm)采集深度为0—10 cm的土壤样品,每个样地的土壤样品充分混匀后挑去石头和植物残体等杂质,自然风干后分别过2 mm和0.149 mm筛用于土壤理化性质的测定。采用Vario ELIII元素分析仪测定土壤的碳、氮含量,土壤容重用环刀法进行测定,土壤pH值、全钾、全磷和有效磷等的分析测定方法参照《土壤农化分析》进行[23]。样地的基本信息统计如表1所示。
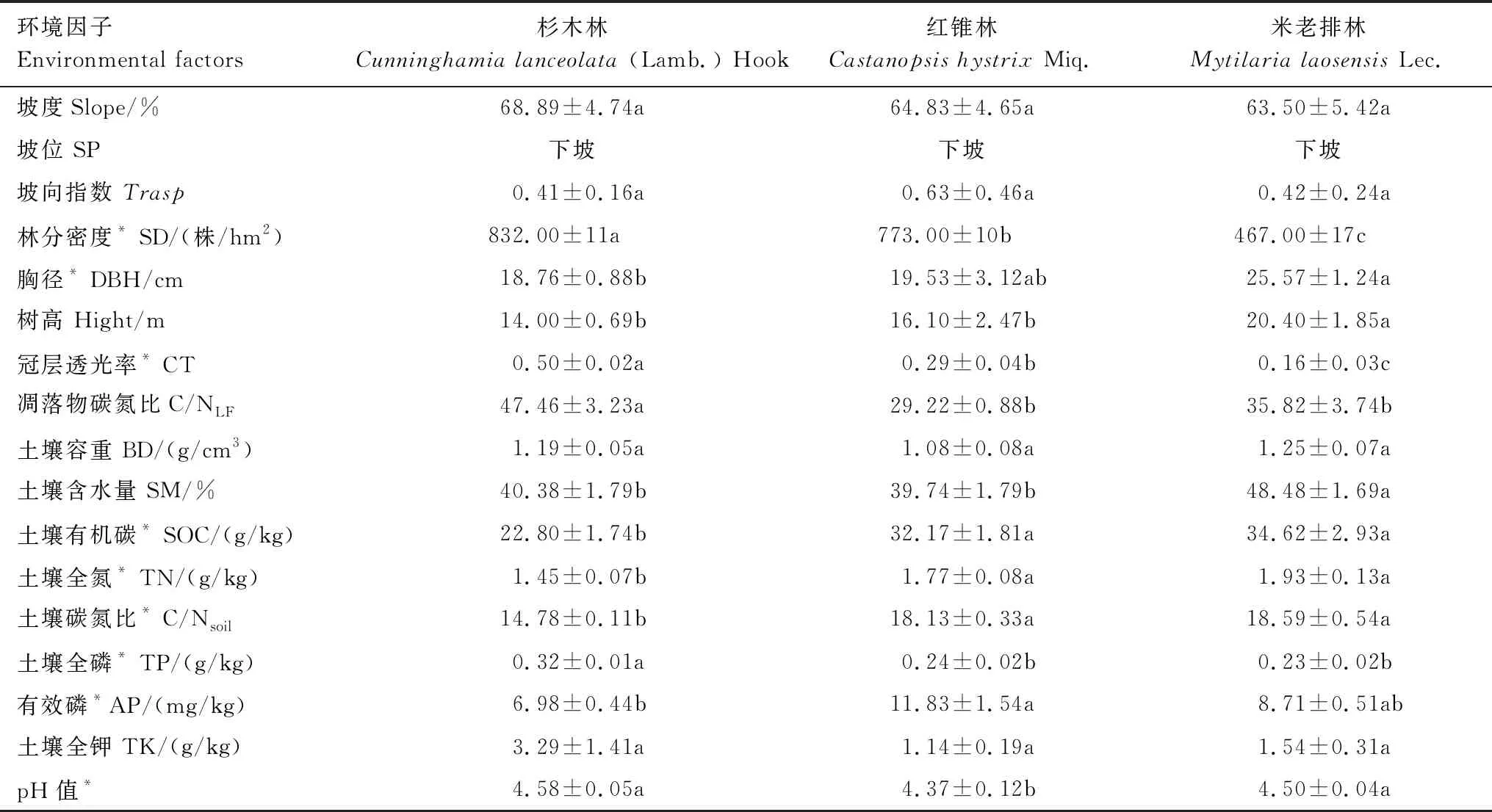
表1 试验样地的基本概况(平均值±标准误)
1.3 数据分析
采用相对重要值(IV)反映林下灌木层和草本层植物物种在其相应群落中的地位和优势,其计算公式如下:
IV=(相对多度+相对盖度+相对频度)/3
(1)
采用物种丰富度(S)、Shannon-Wiener指数(H)、Simpson优势度指数(D)和Pielou均匀度指数(Jsw)研究不同样地灌木层和草本层植物物丰富度和α多样性的变化情况,其中:
S=Ns
(2)
式中,Ns表示样方内物种总数
(3)
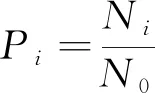
(4)
JSW=H/InS
(5)
采用 β 多样性指数描述不同环境梯度下灌木层和草本层物种多样性的变化,Whittaker β 多样性指数:
(6)
式中,S是研究系统中所记录到的总物种数,A为环境梯度上(样方)所发现物种的平均数。
运用SPSS 17.0软件的单因素方差分析(one-way ANOVA)分别检验不同林分之间灌木层和草本层植物的物种丰富度、Shannon-Wiener指数、Simpson优势度指数、Pielou均匀度指数和Whittaker β 多样性指数的差异性(α=0.05)。
在本研究中,把实验获得的17个环境因子归为3类,包括微地形:坡向、坡位、坡度;乔木特性:林分密度(SD)、胸高断面积(BA)、冠层透光率(TC)、凋落物量(LF)和凋落物碳氮比(C/NLF);土壤理化性质:pH值、土壤容重(BD)、土壤含水率(SM%)、土壤有机碳(SOC)、总氮(TN)、土壤碳氮比(C/Nsoil)、全钾(TK)、全磷(TP)、有效磷(AP)。不同的植物在生长过程中除了受其生物学特性的影响外,外部环境(如:土壤理化性质)也是重要的影响因素,因此,植物群落的物种组成可能会随着土壤养分种类和含量的变化而发生改变,土壤理化性质与植物群落多样性之间存在密切的关系[21]。
用主成分分析(Principal component analysis, PCA)分别比较杉木林改造成红锥林和米老排林后其灌木层和草本层植物物种组成的分异程度;采用冗余分析(Redundancy analysis)确定影响研究样地灌木层和草本层植物物种组成的最主要环境因子。分别采用灌木层和草本层的重要值作为物种数据,其中灌木层物种数据矩阵为15(样方数) ×75(物种数),草本层物种数据矩阵为15(样方数) × 31(物种数)。在环境变量进入RDA分析之前需经过转换和量化,其中坡向的转换采用将罗盘测定的0—360°之间的方位角转换为0—1之间的数值(transformation of aspect, TRASP)。转换公式如下:
TRASP ={1-cos[(π/180)(aspect-30)]}/2
(7)
式中,TRASP为坡向指数,aspect为罗盘测得的坡向方位角度[25]。而对于样地的坡位,上、中、下坡分别采用1、2、3进行赋值[26]。
以上PCA和RDA分析均在CANOCO for Windows 4.5 软件上进行。
运用“Vegan”数据包里的“Varpart”功能对影响灌木层和草本层物种组成的微地形、乔木层特性和土壤理化性质3类生境因子进行方差分解(Variation partitioning),该程序在R 3.01(R-Development Core Team 2009)上运行,所有分析的数据均经过海灵格(Hellinger)转换。运用韦恩图显示各类生境因子的独立效应和它们之间的交互效应,图中的数字代表各类生境因子的独立效应或交互效应对灌木层或草本层物种组成的解释率[27]。
2 结果分析
2.1 杉木林改造成红锥和米老排林后对其灌木层和草本层植被特征和物种多样性的影响
由表2可知,杉木林改造成红锥林和米老排林23年后,林下植物的总物种数分别降低了47.43%和52.56%,灌木层和草本层的的主要植被物种也发生了明显变化。在杉木林中,其灌木层以水东哥、紫麻、杉木幼树(苗)、山蒟和玉叶金花为主;而改造后,红锥林的灌木层以红锥幼树(苗)、杉木幼树(苗)、九节、酸藤子和琴叶榕为主,米老排林的灌木层以米老排幼树(苗)、杉木幼树(苗)、红锥幼树(苗)、粗叶榕和单叶省藤为主。在草本层,杉木林以金毛狗、蔓生莠竹、东方乌毛蕨、扇叶铁线蕨和红马蹄草为主;而红锥林以扇叶铁线蕨、金毛狗、淡竹叶、深绿卷柏和高秆珍珠茅为主,米老排林以扇叶铁线蕨、铁芒萁、金毛狗、东方乌毛蕨和淡竹叶为主。
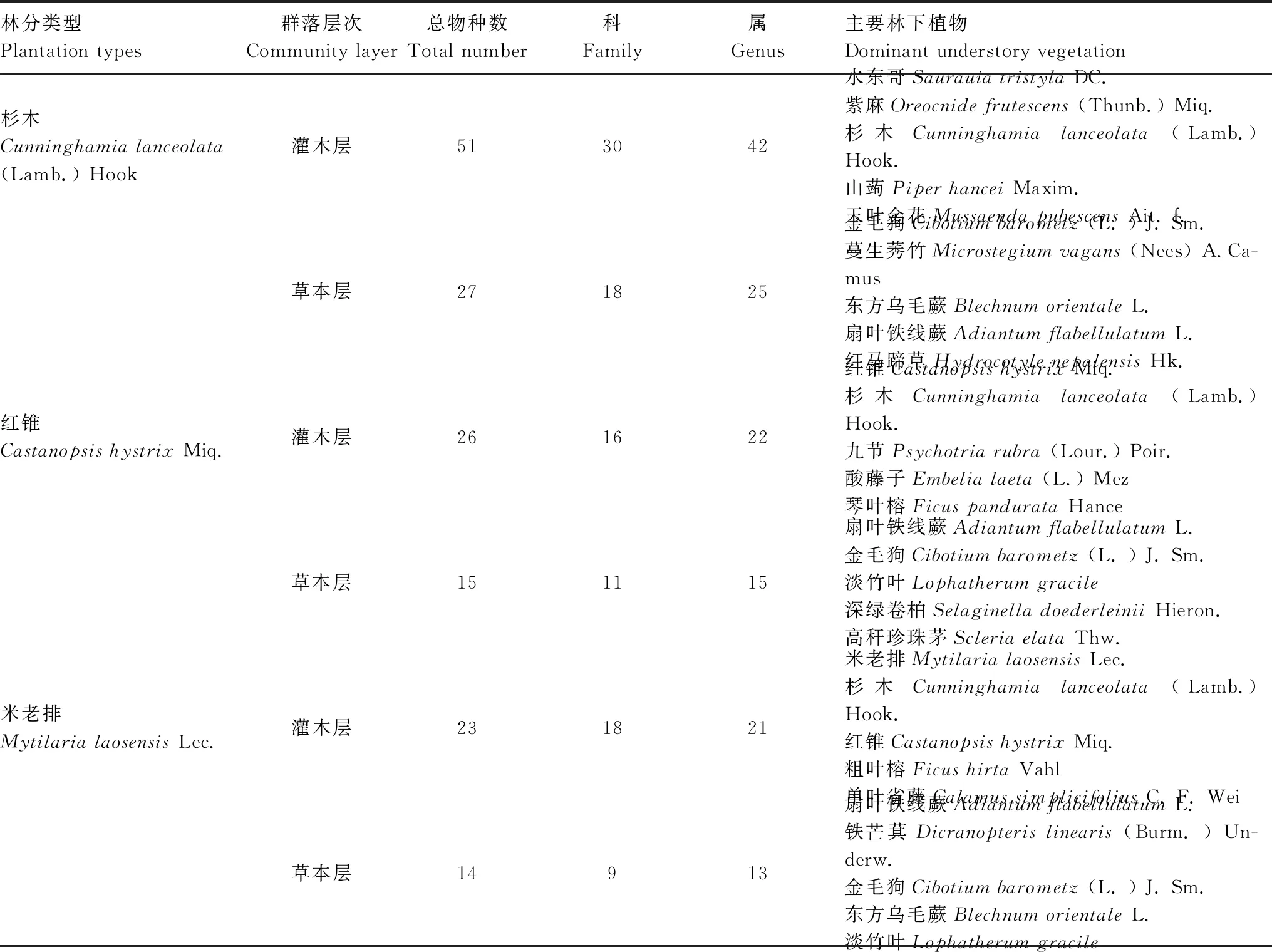
表2 不同人工林类型林下的主要植被特征
林下植物物种多样性指数研究结果如图1所示,与杉木林相比,改造后的红锥林和米老排林的灌木层植物物种丰富度分别极显著降低了64.55%和61.82%(P<0.01),Shannon-Wiener指数分别极显著降低了73.51%和63.53%(P<0.001),Simpson优势度指数分别极显著降低了70.07%和54.10%(P<0.001),Pielou均匀度指数分别极显著降低了63.43%和48.31%(图1),Whittaker β 多样性指数分别极显著增加了3.20倍和2.68倍;改造后的红锥林和米老排林的草本层植物物种丰富度分别极显著降低了53.62%和55.07%(P<0.001),但Shannon-Wiener指数、Simpson优势度指数和Pielou均匀度指数均无显著变化(P>0.05),而Whittaker β 多样性指数分别极显著增加了2.51倍和2.47倍(图1)。
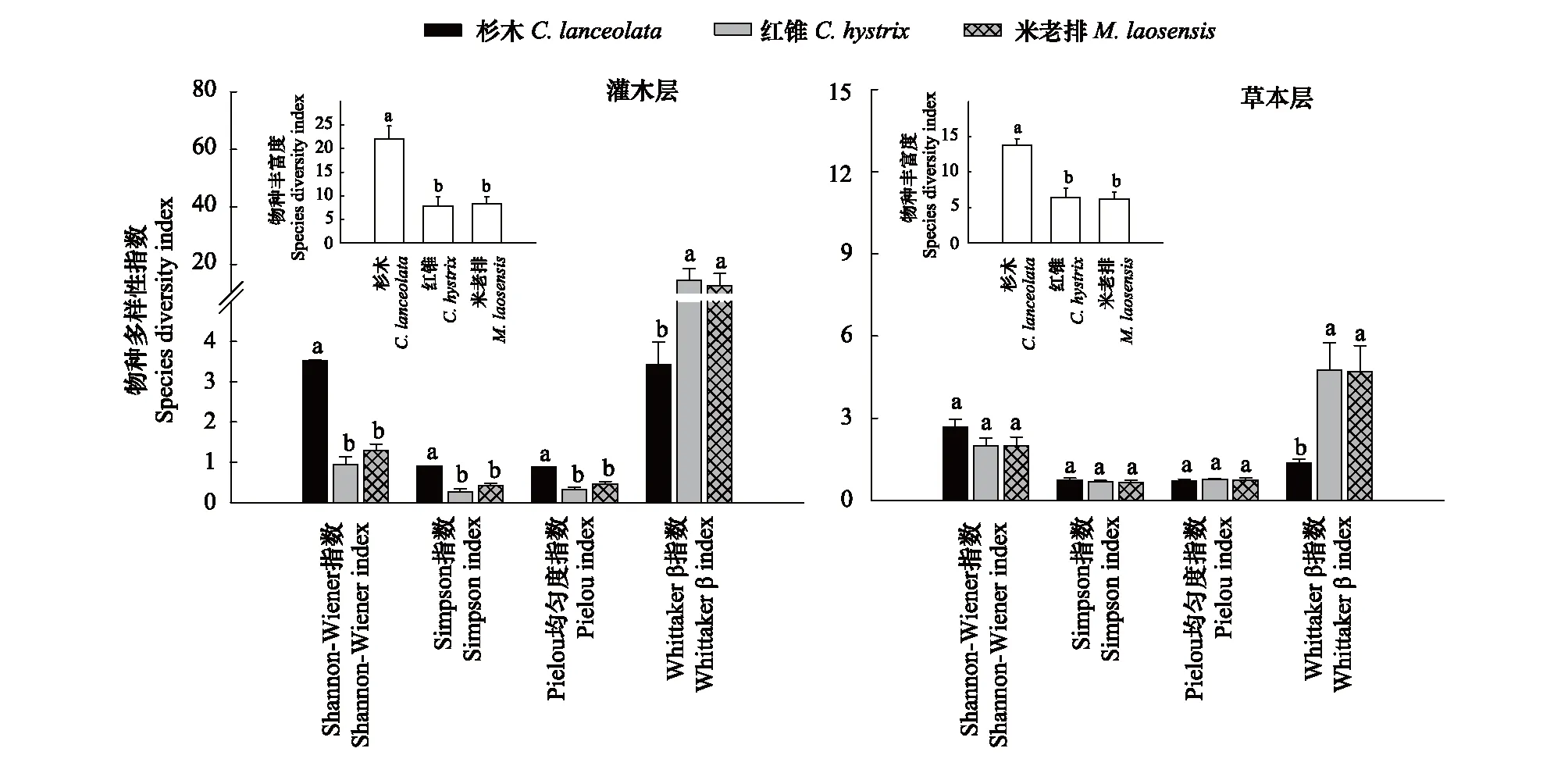
图1 杉木林改造成红锥和米老排林后灌木层和草本层植物物种丰富度与多样性的变化(平均值±标准误,n=5)
2.2 杉木林改造成红锥和米老排林后对灌木层和草本层植物物种组成的影响
对杉木人工林改造成红锥和米老排林23年后的灌木层和草本层植物物种的重要值进行主成分分析(PCA),结果表明:在灌木层(图2),第一主轴(PCA1)和第二主轴(PCA2)分别解释其植物物种组成变异的54.2%和30.0%,其中PCA1将红锥林与杉木林和米老排林完全区分开来,PCA2又显见的将杉木林与红锥林和米老排林完全区分开来;在草本层(图2),PCA1和PCA2分别解释了林下草本层植被物种组成变异的29.4%与24.6%,其中PCA2完全把杉木林与红锥林和米老排林区分开。这些结果表明杉木人工林进行阔叶化改造后,其林下灌木层和草本层的植物物种组成均发生了明显改变,而灌木层的变异程度要大于草本层。
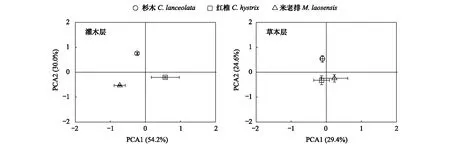
图2 杉木人工林改造成红锥和米老排林后灌木层和草本层物种组成的主成分分析(PCA)(平均值±标准误,n=5)
2.3 杉木林改造成红锥和米老排林后灌木层和草本层植物物种组成和环境因子之间的关系
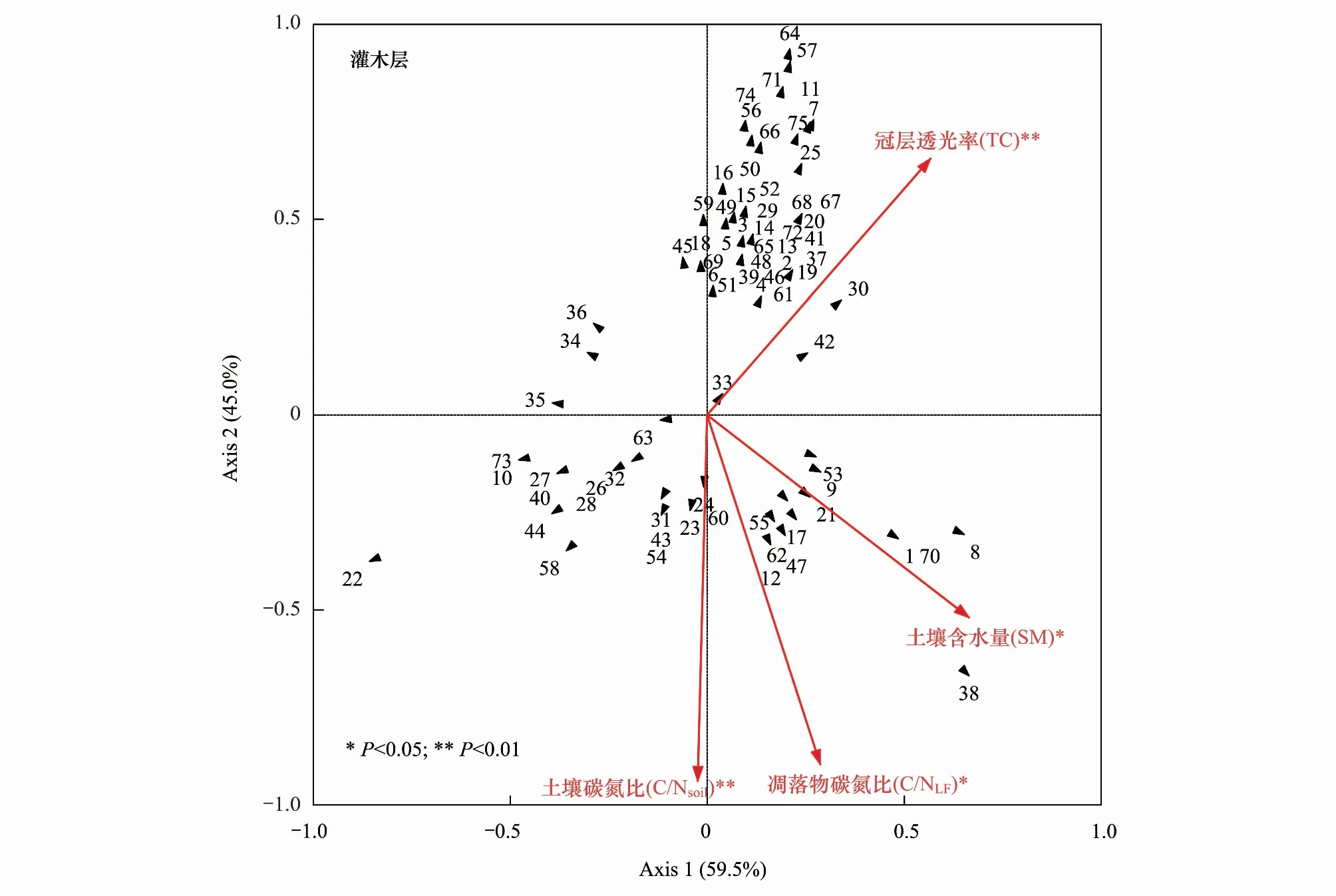
杉木人工林改造成乡土阔叶树种红锥和米老排后,林下植物物种组成发生了明显的变化。进一步通过对其林下植物物种组成和环境因子之间的关系进行冗余分析(RDA), 结果表明:在灌木层,对实验获得的17个环境因子进行蒙特卡洛检验预选,确定冠层透光率、土壤含水量、凋落物碳氮比、土壤碳氮比和4个环境因子与灌木层植物物种组成显著相关(P<0.05),并被选入最后的模型分析,最终得到冠层透光率和土壤碳氮比是影响林下灌木层植物物种组成的极显著环境因子(P<0.01),土壤含水量和凋落物碳氮比是影响林下灌木层植物物种组成的显著环境因子(P< 0.05)(图3);在草本层中,对实验获得的17个生物和非生物因子进行蒙特卡洛检验预选,确定凋落物碳氮比、土壤有机碳、土壤碳氮比、土壤含水量、冠层透光率5个环境因子与草本层植物物种组成显著相关(P<0.05),并被选入最后的模型分析,最终得到冠层透光率和土壤碳氮比影响林下草本层植物物种组成的显著环境因子(P<0.05)(图3)。
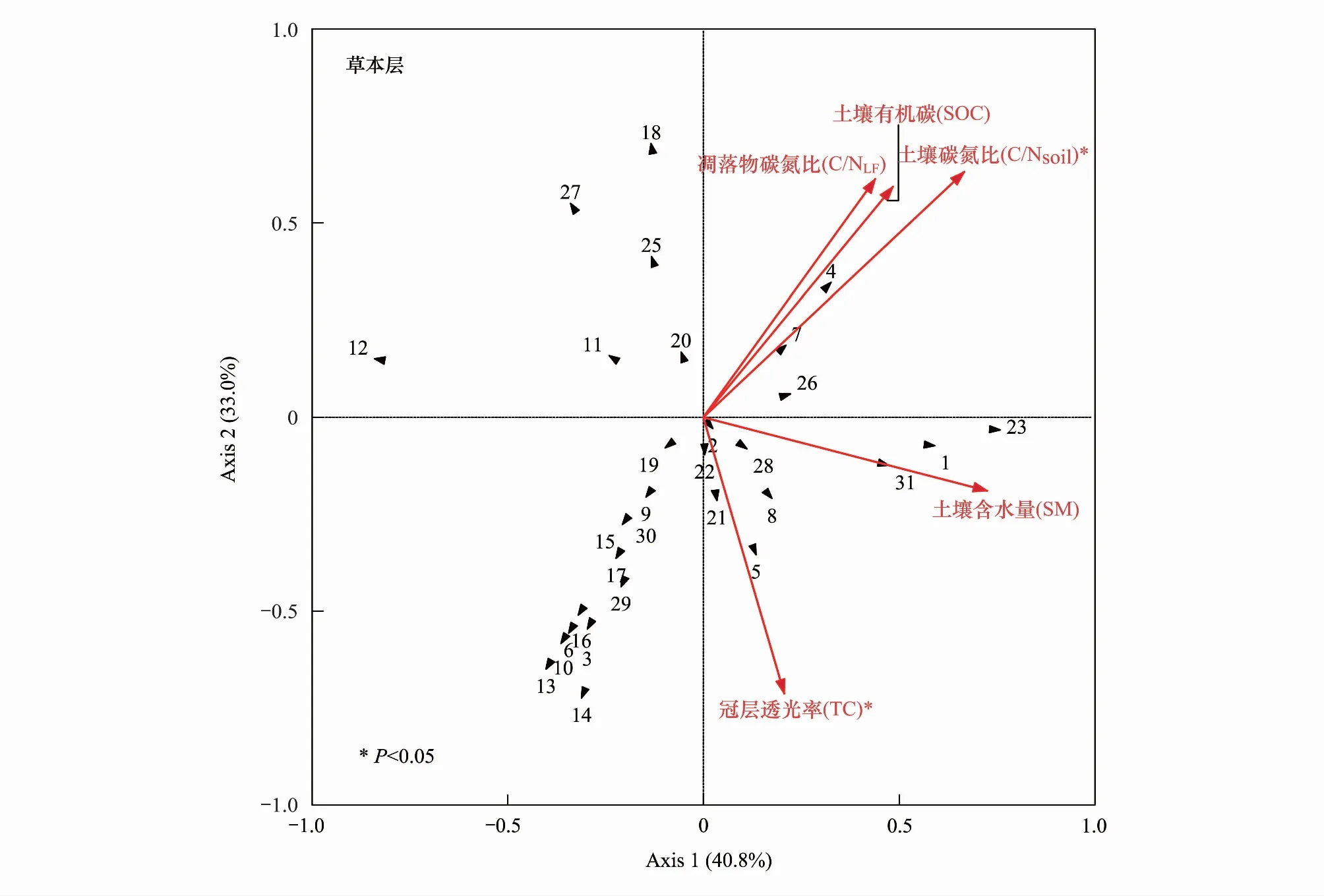
图3 杉木林、红锥林和米老排林的灌木层和草本层物种组成与主要环境因子的冗余分析(RDA)
2.4 影响杉木林、红锥林和米老排林灌木层和草本层植物物种组成的生境因子变异分离
分析结果表明,本研究所调查的三类生境因子,包括微地形、乔木层特性和土壤理化性质的总效应(纯效应和共同效应)分别解释了灌木层和草本层植物物种变异的80.9%和84.6%(图4)。在灌木层,影响其植物物种组成的因素主要来自乔木层特性的独立效应,以及乔木特性与微地形的共同效应,其次是土壤理化性质的独立效应,而微地形的独立效应以及微地形、乔木层特性和土壤理化性质三类因子之间的交互作用对灌木层植物物种组成的影响较小;在草本层,影响其植物物种组成的因子主要来自乔木层特性和土壤理化性质的独立效应,其次是微地形、乔木层特性和土壤理化性质三类因子之间的交互作用微地形的独立效应以及微地形的独立效应,而微地形与土壤理化性质的交互作用,以及乔木层特性与土壤理化性质的交互作用对草本层植物物种组成的影响较小。
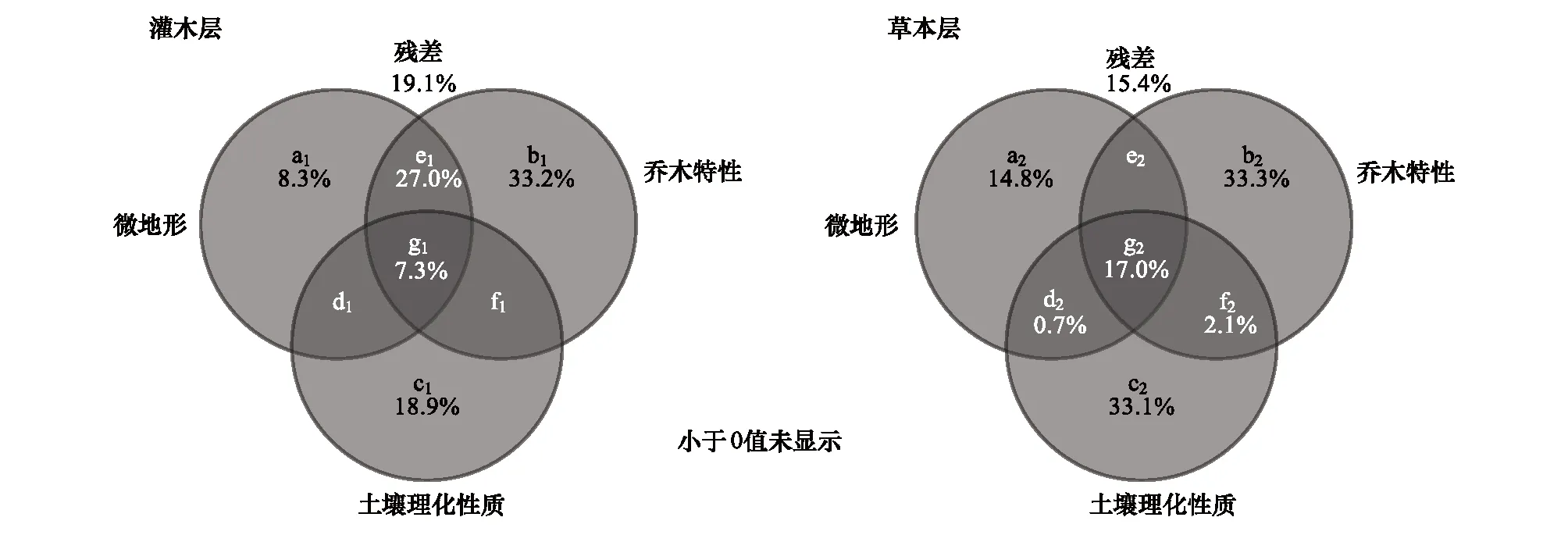
图4 影响灌木层和草本层植物物种分布的微地形、乔木层特性和土壤理化性质的方差分解分析
3 结论与讨论
林下植被作为人工林生态系统的重要组成部分,在一定程度上弥补人工林树种单一化种植的天然缺陷,其在调控人工林生态系统结构及功能的稳定性和多样性等方面发挥不可替代的作用[28]。本研究发现,杉木林改造成乡土阔叶树种红锥林和米老排林23年后,其乔木树种的变化导致林冠结构和土壤理化性质发生了明显改变,进而导致其林下灌木层和草本层植被群落的物种丰富度显著降低,这与秦新生等[29]的研究结果一致,认为相对于阔叶林,针叶林林下植物物种丰度较高,这主要归咎于针叶林的郁闭度更低而能为更多物种的繁殖和生长提供充足的光照。人工林林下植物群落多样性对维持其整个生态系统结构的稳定性具有极其重要的作用,而光照条件是影响林下植物群落物种多样性的最重要因素之一[30—31],改善林下群落的光照条件,为更多物种的侵入、定居和繁殖更新提供更加丰富和多样化的生存环境[32]。姜俊等[21]的研究发现,人工林林下植物的物种多样性指数与冠层郁闭度呈负相关关系,认为改善冠层的光照,有利于增加林下植被的多样性,这与本研究的结果相似。杉木林改造成红锥或米老排林后,其林分郁闭度增加而减少了冠层的透光率,显著降低了灌木层的植物物种多样性,但其草本层的植物物种多样性没有显著变化,表明其草本层植物物种多样性对林分改变的响应程度要低于灌木层植物物种,这与刘玉宝等[33]的研究结果相似。
人工林林下灌木层和草本层植物群落的物种组成是生物和非生物因子综合作用结果,其影响过程极其复杂[34—35]。Yu 和 Sun[36]的研究结果表明,不同的林分类型导致其冠层结构、土壤理化性质和微环境差异明显,最终影响其林下植被的物种组成。在人工林生态系统中,林下植被群落的物种组成不仅受到经营树种和经营管理措施的影响,还受到地形、地貌和其他微环境的影响[37—38]。人工林不同种植树种的冠层结构,被认为是影响其林下植物物种组成的最重要因子之一,其不仅可以通过改变林下植物的光照条件而影响不同植物物种的生长繁殖,也可以影响林下土壤的水分而调控不同植物物种的分布[39—40],这与本研究的结论相似。本研究的RDA分析结果较好地反映了生物和非生物因子与人工林林下植物物种组成的关系,揭示了杉木林改造成红锥林或米老排林后所导致的冠层透光率的显著变化是驱动其灌木层和草本层植物物种组成发生明显变化的最主要因子之一。此外,在本研究中也发现土壤碳氮比在一定程度上也是影响灌木层和草本层植物物种组成的重要因子之一,这可能与本研究的杉木林与红锥林或米老排林之间明显不同的凋落物碳氮比导致的土壤碳氮比存在显著差异有关,这也与一些研究认为的土壤养分质量是影响小区域尺度植被物种分布格局的重要因子的结论相似[41—42]。
土壤水分,在大的区域乃至全球尺度上都被认为是影响植物物种组成和分布的关键因子之一。但本研究发现,土壤水分含量只是影响杉木林、红锥林和米老排林灌木层的植物物种组成的重要因子,而对草本层植物物种组成的影响较少,这表明本研究林分灌木层的植物物种组成对土壤水分的敏感度比草本层植物要高。许多前期的研究表明不同的海拔梯度上植物的分布存在显著性的差异,海拔梯度被认为是调控物种分布格局的主要因子之一[43],但本研究发现海拔没有显著影响研究林分林下植被的物种组成,这可能与本研究的海拔梯度变化范围小有关。在地形因子中,坡向、坡位和坡度等在一定程度上能够影响小尺度上的光照、水分,因此在一定程度上也能够影响物种的组成[44—45],但本研究却发现这些因子都不是显著影响其灌木层和草本层植物物种组成的重要因子,这可能与本研究的3种林分类型的地形因素相似有关。
同时,本研究所调查的微地形、乔木层特性和土壤理化性质3类生境因子的独立效应和交互效应对灌木层或草本层植物物种组成的解释率均超过80%,表明所调查的生境因素能解释灌木层或草本层植物物种组成的大部分变异。此外,微地形、乔木层特性和土壤理化性质3类生境因子的独立效应对灌木层和草本层植物物种组成的解释要大于它们的交互效应,而且对灌木层和草本层植物物种组成的解释率有所不同。在灌木层,乔木层特性的独立效应及其与微地形的交互效应对物种组成的解释率要大于微地形和土壤理化性质的独立效应或交互效应,表明杉木林改造成红锥或米老排林后导致的乔木层特性变化是导致其灌木层植物物种变异的主要原因;而在草本层中,乔木层特性和土壤理化性质的独立效应能解释超过60%的植物物种组成的变异,表明杉木林改造成红锥或米老排林后导致的乔木层特性和土壤理化性质变化是导致其草本层植物物种变异的主要原因。由此可见,构建合理的林分的冠层结构和改善土壤理化性质对于增加该区域人工林林下植被物种多样性并促进其发育和演替至关重要。许多研究表明,相对于单一树种的针叶纯林,通过混交改造后形成的针/阔混交林具有更加丰富的冠层结构、更肥沃的土壤养分、更高的生物多样性和生产力等[21,46—49]。此外,林分密度也是影响林下植被物种组成和多样性的重要因素之一[21, 50]。
综上所述,杉木林改造成乡土阔叶树种红锥林和米老排林23年后,相对于杉木林,红锥林和米老排林的灌木层植物物种丰富度和α多样性指数均显著降低,但草本层植物只有物种丰富度显著降低,其α多样性指数无显著变化。杉木林改造成红锥林和米老排林后冠层透光率、土壤碳氮比、土壤含水量和凋落物碳氮比的显著变化是导致其林下灌木层植物物种组成发生明显变异的主要因子,而冠层透光率和土壤碳氮比的显著变化是导致其草本层植物物种组成发生明显变异的主要因子。因此,在把大面积分布的杉木林改造成乡土阔叶林过程中,有必要注意适度调整林分密度而改善林分冠层结构,或构建针阔异龄混交林,形成不同树龄、多层次、近自然化的混交林群落,从而提高凋落物的数量和质量,丰富土壤中的腐殖质层和恢复土壤肥力,为更多林下植物物种的侵入、定居、繁殖和更新提供更加丰富和多元化的生境,增加林下生物多样性,促进树木的持续健康生长,以提高人工林生态系统的稳定性和功能的多样性。
