MiR-181a-5p 通过靶向KLF6 信号调控低氧诱导的肺动脉平滑肌细胞的增殖和迁移的机制研究
2022-04-04陈妮赵梅
陈妮,赵梅
1.海南医学院第二附属医院药学部,海南海口 570311;2.三亚中心医院(海南省第三人民医院)药学部,海南三亚 572000
肺动脉高压(pulmonary arterial hypertension, PAH)是一种罕见的疾病,其特征是小肺动脉进行性闭塞,导致肺动脉压升高和右心衰竭[1-2]。在过去的几十年中,对该疾病的病理生理学的深入了解导致开发了针对内皮功能障碍的有效疗法(依前列醇及其衍生物、内皮素受体拮抗剂和5 型磷酸二酯酶抑制剂)[3]。这些药物可以改善临床、功能和血流动力学。尽管如此,PAH 尚无治愈方法,预后仍然很差。MicroRNAs(miRNAs, miRs)是一类内源性的小型非编码RNA,其通过靶向mRNA 的3'-非翻译区(3’-untranslated region, UTR)来负调控基因表达,以进行基因降解或翻译抑制[4]。miRNA 具有多种细胞功能,包括调节细胞代谢、发育、增殖和死亡。研究发现多种miRNA 在PAH 的病理生理中起到关键的调控作用[5]。最近的研究发现miR-181a-5p 能够通过抑制炎症反应来缓解大鼠的肺动脉症状,提示miR-181a-5p 在PAH 的病理生理中起到关键的作用[6]。然而miR-181a-5p 调控PAH,特别是调控肺动脉血管平滑肌细胞增殖、迁移等分子机制尚未明确。因此,本研究2021 年11 月—2022 年5月进行,探讨了miR-181a-5p 对肺动脉血管平滑肌细胞增殖以及迁移的影响,并探索了相应的分子调控通路。现报道如下。
1 材料与方法
1.1 材料来源
DMEM 培 养 基、FBS 胎 牛 血 清、Lipofectamine 2000 转染试剂购于美国Thermo Fisher Scientific 公司;肺动脉平滑肌细胞购于美国ATCC 公司;miR-181a-5p 模拟 物,模拟物对照,miR-181a-5p 抑制剂,抑制剂对照,pcDNA3.1 以及pcDNA3.1-KLF6 购于广州锐博生物科技有限公司;萤光素酶报告载体以及萤光素酶检测试剂盒购于美国Promega 公司;CCK-8 检测试剂盒购于北京碧云天生物科技有限公司。实时定量PCR 试剂盒以及Trizol 试剂购于日本Takara 公司。
1.2 方法
1.2.1 细胞培养将肺动脉平滑肌细胞用含10%胎牛血清的不含抗生素的DMEM 培养基悬浮后,置于条件为37℃、5%的CO2的恒温细胞培养箱中培养。每天在镜下观察细胞生长情况,并根据情况进行换液,当细胞融合度达80%以上时可进行消化传代。取对数生长期细胞进行后续实验。
1.2.2 细胞转染当肺动脉平滑肌细胞细胞贴壁后,使用Lipofeetamine 2000 试剂根据使用说明对细胞进行相应的质粒和miRNA 进行转染,转染后24 h,进行相关的试验。
1.2.3 处理分组第1 组实验细胞分别用常氧(21%O2和5%CO2)或 者 低 氧(3%O2、5%CO2和92%N2),分别处理12、24 h 或者48 h 后,收集细胞进行相关实验;第2 组实验:细胞分别用常氧(21%O2和5%CO2)、低氧(3%O2、5%CO2和92%N2)或者低氧(3%O2、5%CO2和92%N2)联合miR-181a-5p 模拟物转染处理人肺动脉平滑肌细胞,24 h 后,收集细胞进行相关实验。第3 组实验:人肺动脉平滑肌细胞利用miR-181a-5p 抑制剂或者抑制剂对照转染,24 h 后,收集细胞进行相关实验。第4 组实验:细胞分别用低氧(3%O2、5%CO2和92%N2)、低氧(3%O2、5%CO2和92%N2)联合miR-181a-5p 模拟物转 染、低 氧(3%O2、5%CO2和92%N2)联 合miR-181a-5p 模拟物以及pcDNA3.1-KLF6 处理人肺动脉平滑肌细胞,24 h 后,收集细胞进行相关实验。
1.2.4 实时定量PCR使用Trizol 试剂盒从细胞系中分离出包括miRNA 的总RNA。分别使用RNA 反转录试剂盒和PrimeScript RT 试剂盒进行反转录。使用SYBR 预混Ex Taq 在ABI 7500 实时PCR 系统 上进行实时荧光定量PCR。U6 作为miR-181a-5p 的表达内参,GAPDH 作为PCNA 的内参;用2-ΔΔCt法计算相关基因的相对表达水平。
1.2.5 CCK-8 试验使用CCK-8 检测试剂盒检测细胞活性。收集经过不同处理的细胞,以每孔1×103 个细胞的密度接种到96 孔板中,24 h 后,将CCK-8 溶液加入各孔中,于37℃下孵育2 h,450 nm 处测定吸光度,计算细胞活性。
1.2.6 双荧光素酶试验采用TargetScan 预测发现,miR-181a-5p 和KLF6 3'-UTR 有连续的结合位点。按照说明书将KLF6 3‘UTR 野生型或突变型的荧光素酶报告基因质粒载体和模拟物对照或者miR-181a-5p 模拟物分别或同时转入肺动脉平滑肌细胞。培养48 h 后小心吸弃培养液,洗涤细胞后加入细胞裂解液裂解细胞5~10 min,3 000 r/min 离心5 min,取上清,利用双荧光素酶报道试剂盒对荧光素酶活性进行测定。
1.3 统计方法
采用GraphPad Prism 7.0 软件分析数据,计量资料符合正态分布,以(±s)表示,两组间比较采用t检验,多组间比较采用单因素方差分析,多组间两两比较采用Bonferroni's 校正检验。P<0.05 为差异有统计学意义。
2 结果
2.1 低氧诱导增强肺动脉平滑肌细胞的活性,下调肺动脉平滑肌细胞中的miR-181a-5p 相对表达水平
CCK-8 结果显示,低氧处理肺动脉平滑肌细胞0 h 组(100.00±15.10),12 h 组(110.3±10.1),12 h 后未增强肺动脉平滑肌细胞的活性,差异无统计学意义(t=0.985,P>0.05),见 图1A。 0 h 组(100.00±15.10),24 h(145.7±10.2),差异有统计学意义(t=4.353,P<0.05);0 h 组(100.00±15.10),48 h 组(147.7±15.3),差 异 有 统 计 学 意 义(t=4.545,P<0.05),24、48 h 低氧处理能够增加肺动脉平滑肌细胞的活性,见图1A。实时定量PCR 检测结果显示,0 h 组(1.00±0.06),12 h 组(0.87±0.053),差异无统计学意义(t=1.561,P>0.05),12 h 低氧处理对肺动脉平滑肌细胞中miR-181a-5p 的相对表达水平未影响,见图1B。0 h 组(1.00±0.06),24 h 组(0.49±0.15),差异有统计学意义(t=4.545,P<0.001);0 h 组(1.00±0.06),48 h 组(0.43±0.12),差异有统计学意义(t=6.882,P<0.001),24、48 h 低氧处理能够显著下调肺动脉平滑肌细胞中miR-181a-5p 的相对表达水平,见图1B。
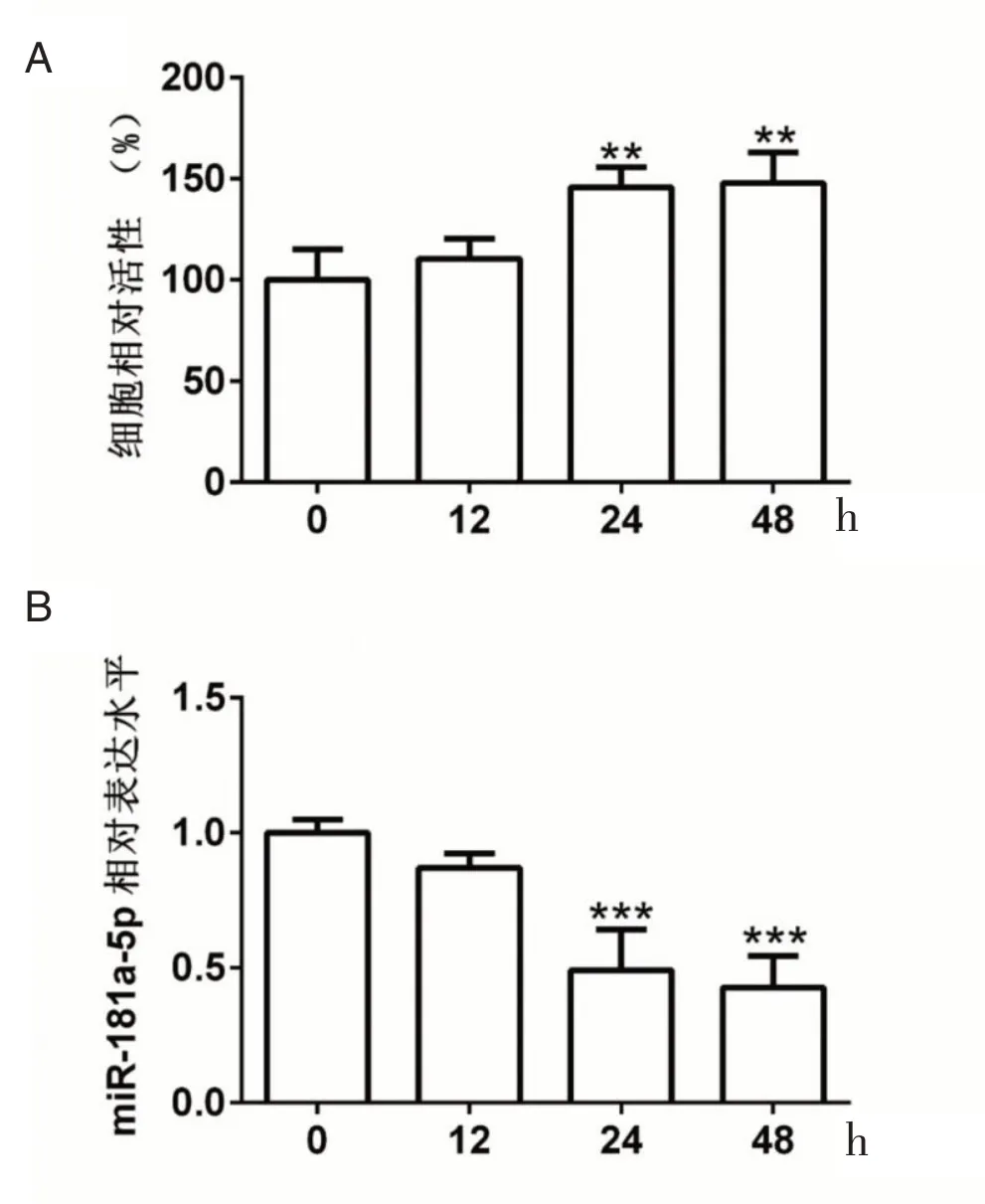
图1 低氧诱导增强肺动脉平滑肌细胞的活性,下调肺动脉平滑肌细胞中的miR-181a-5p 相对表达水平
2.2 miR-181a-5p 过表达抑制低氧诱导的肺动脉平滑肌细胞活性以及迁移
miR-181a-5p 模拟物转染后,miR-181a-5p 模拟物组(9.20±1.95),模拟物对照组(1.00±0.12),24 h 能够显著提高肺动脉平滑肌细胞中的miR-181a-5p相对表达水平,差异有统计学意义(t=7.282,P<0.05),见图2A。低氧 组(154.30±7.60),对照组(100.00±26.10),24 h 低氧处理能够增强肺动脉平滑肌细胞的活性,差异有统计学意义(t=4.166,P<0.05);上调PCNA 的表达水平,低氧组(2.06±0.29),对照组(1.00±0.11),差异有统计学意义(t=5.919,P<0.05),见图2B、图2C。低氧+miR-181a-5p 模拟物组(107.00±5.60),低氧组(154.30±7.60),miR-181a-5p 过表达能够显著下调低氧诱导的肺动脉平滑肌细胞活性,差异有统计学意义(t=3.630,P<0.05),见图2B。PCNA 的表达水平,低氧+miR-181a-5p 模拟物组(1.33±0.25),低氧组(2.06±0.29),差异有统计学意义(t=3.929,P<0.05),见图2C。进一步的划痕修 复 试 验 发 现,低 氧 组(71.67±6.51),对 照 组(46.67±6.66),24 h 低氧处理能够促进肺动脉平滑肌细胞的迁移,差异有统计学意义(t=4.782,P<0.05),见 图2D。低 氧+miR-181a-5p 模 拟 物 组(51.67±6.03),低氧组(71.67±6.51),差异有统计学意义(t=3.825,P<0.05),miR-181a-5p 过表达能够显著下调低氧诱导的肺动脉平滑肌细胞迁移能力,见图2D。

图2 miR-181a-5p 过表达抑制低氧诱导的肺动脉平滑肌细胞增殖以及迁移
2.3 miR-181a-5p 敲减促进低氧诱导的肺动脉平滑肌细胞增殖以及迁移
通过转染试验,miR-181a-5p 抑制剂组(0.43±0.07),抑制剂对照组(1.00±0.28),miR-181a-5p 抑制剂转染后24 h 能够显著下调肺动脉平滑肌细胞中的miR-181a-5p 相对表达水平,差异有统计学意义(t=3.410,P<0.05),见图3A。CCK-8 试验检测,miR-181a-5p 抑制剂组(152.70±10.40),抑制剂对照组(100.00±21.90),miR-181a-5p 抑制剂转染能够显著增强肺动脉平滑肌细胞的活性,差异有统计学意义(t=3.758,P<0.05),见图3B。实时定量PCR 检测,miR-181a-5p 抑制剂组(1.60±0.12),抑制剂对照组(1.00±0.04),miR-181a-5p 抑制剂转染能够显著上调肺动脉平滑肌细胞中PCNA 的表达水平,差异有统计学意义(t=8.202,P<0.05),见图3C。划痕修复试验,miR-181a-5p 抑制剂组(68.67±6.81),抑制剂对照组(47.67±8.33),miR-181a-5p 抑制剂转染能够显著增强肺动脉平滑肌细胞的迁移能力,差异有统计学意义(t=3.382,P<0.05),见图3D。
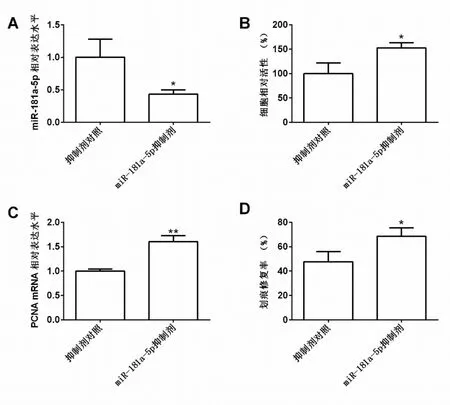
图3 miR-181a-5p 敲减促进低氧诱导的肺动脉平滑肌细胞增殖以及迁移
2.4 MiR-181a-5p 负向调控肺动脉平滑肌细胞中KLF6 的表达
通过TargetScan 在线生信分析工具,发现miR-181a-5p 与KLF6 3’UTR 区域存在相互结合的区域,见图4A。通过萤光素酶报告试验检测发现,miR-181a-5p 模 拟 物 组(0.26±0.089)模 拟 物 对 照 组(1.00±0.14),miR-181a-5p 模拟物转染能够显著降低野生型KLF6 3’UTR 萤光素酶报告载体的萤光素酶活性,差异有统计学意义(t=7.729,P<0.05),见图4B。miR-181a-5p 模拟物组(1.04±0.12),模拟物对照组(1.00±0.13),miR-181a-5p 模拟物转染对突变型KLF6 3’UTR 萤光素酶报告载体的萤光素酶活性没有影响,差异无统计学意义(t=0.421,P>0.05),见图4C。实时定量PCR 检测结果显示,miR-181a-5p模拟物组(0.36±0.11),模拟物对照组(1.00±0.03),miR-181a-5p 模拟物转染能够显著下调肺动脉平滑肌细胞中的KLF6 mRNA 表达水平,差异有统计学意义(t=9.439,P<0.05),见图4D。低氧组(1.58±0.18),对照组(1.00±0.17),24 h 低氧处理显著下调肺动脉平滑肌细胞中的KLF6 mRNA 表达水平,差异有统计学意义(t=4.108,P<0.05),见图4E。
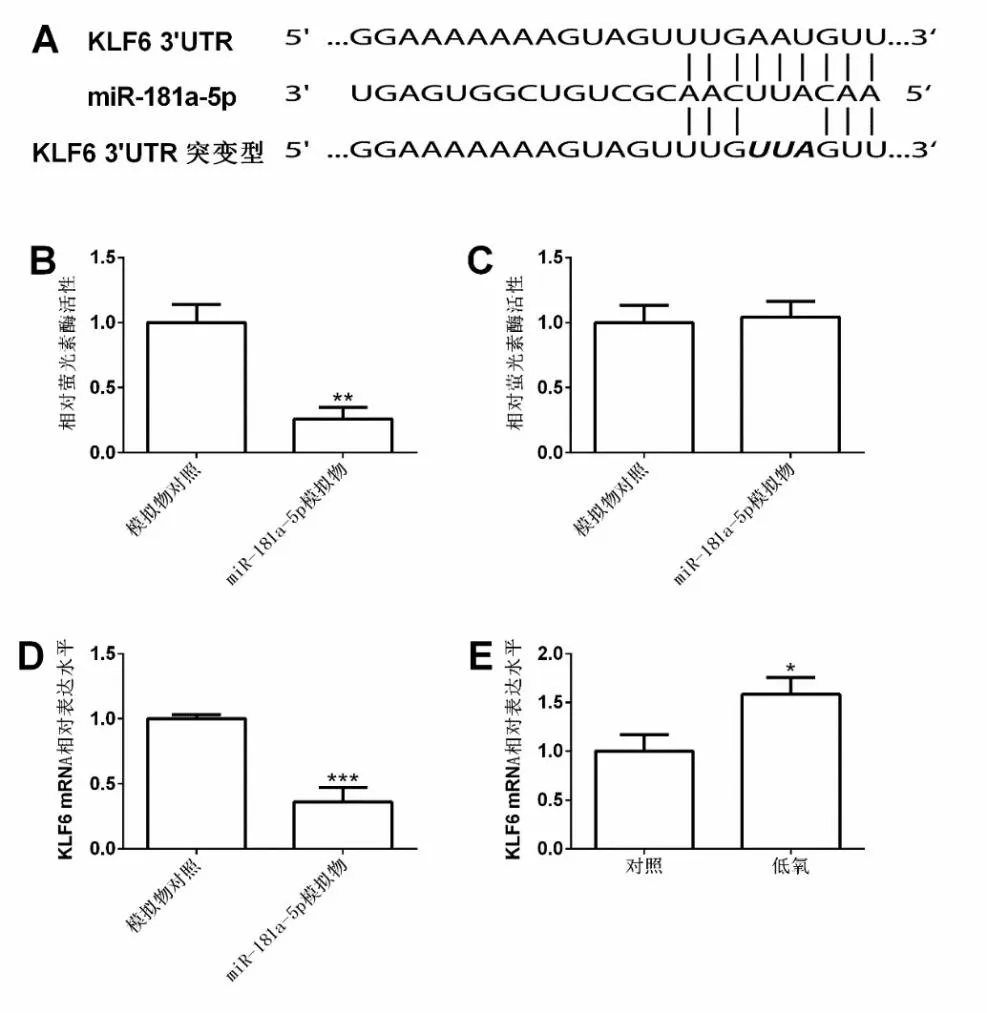
图4 miR-181a-5p 负向调控肺动脉平滑肌细胞中KLF6 的表达
2.5 KLF6 过表达能够拮抗miR-181a-5p 对低氧诱导的肺动脉平滑肌细胞增殖和迁移的抑制作用
pcDNA3.1-KLF6 载体转染能够显著上调肺动脉平滑肌细胞中KLF6 mRNA 的表达水平pcDNA3.1-KLF6 组(3.53±1.43),pcDNA3.1 组(1.00±0.17),差异有统计学意义(t=3.055,P<0.05),见图5A。miR-181a-5p 过表达能够显著下调低氧诱导的肺动脉平滑肌细胞活性,低氧+miR-181a-5p 模拟物组(61.1±4.0),低氧组(100.00±14.50),差异有统计学意义(t=5.408,P<0.05),见图5B;和PCNA 的表达水平,低氧+miR-181a-5p 模拟物组(0.47±0.078),低氧组(1.00±0.12),差异有统计学意义(t=6.645,P<0.05),见图5C,而pcDNA3.1-KLF6 转 染能 够拮抗miR-181a-5p 过表达对低氧诱导的肺动脉平滑肌细胞活性,低氧+miR-181a-5p 模拟物+pcDNA3.1-KLF6 组(86.0±2.6),低氧+miR-181a-5p 模拟物组(61.10±4.00),差 异 有 统 计 学 意 义(t=3.467,P<0.05),见图5B;和PCNA 的表达抑制的影响,低氧+miR-181a-5p 模 拟 物+pcDNA3.1-KLF6 组(0.83±0.081),低氧+miR-181a-5p 模拟物组(0.47±0.078),差异有统计学意义(t=4.542,P<0.05),见图5C。miR-181a-5p 过表达能够显著下调低氧诱导的肺动脉平滑肌细胞迁移能力,低氧+miR-181a-5p 模拟物组(46.33±4.16),低氧组(70.33±6.11),差异有统计学 意 义(t=4.323,P<0.05),见 图5D。pcDNA3.1-KLF6 转染能够拮抗miR-181a-5p 过表达对低氧诱导的肺动脉平滑肌细胞迁移抑制的影响,低氧+miR-181a-5p 模拟物+pcDNA3.1-KLF6 组(66.00±9.17),低氧+miR-181a-5p 模拟物组(46.33±4.16),差异有统计学意义(t=3.543,P<0.05),见图5D。

图5 KLF6 过表达能够拮抗miR-181a-5p 对低氧诱导的肺动脉平滑肌细胞增殖和迁移的抑制作用
3 讨论
本研究发现miR-181a-5p 在低氧诱导的肺动脉血管平滑肌细胞中表达下调;进一步的功能试验发现miR-181a-5p 过表达能够部分反转低氧诱导的肺动脉血管平滑肌细胞的增殖和迁移;miR-181a-5p 敲减能够促进肺动脉血管平滑肌细胞的增殖和迁移。通过进一步的生物信息学预测试验发现miR-181a-5p 能够与KLF6 的3‘UTR 区域结合;功能验证试验进一步发现miR-181a-5p 能够负向调控KLF6 在肺动脉平滑肌细胞的表达,而低氧诱导能够上调KLF6 的表达水平。营救试验发现KLF6 过表达能够反转miR-181a-5p 过表达对低氧诱导的肺动脉血管平滑肌细胞的增殖和迁移的抑制作用。
研究发现miR-181a-5p 在调控心血管细胞的生物学功能方面起到非常关键的作用。相关研究发现miR-181a-5p 能够抑制血管平滑肌细胞的增殖、迁移,同时促进其凋亡,进而调控粥状动脉硬化的病理进程[7]。另有研究表明miR-181a-5p 通过靶向PDGFRA 来抑制血管内皮细胞的血管生成[8]。Deng L 等[9]研究发现miR-181a-5p 能够抑制癌细胞的迁移和血管生成。也有研究发现miR-181a-5p过表达能够抑制网络新生血管,其是通过调控endocan 和ERK1/2 信号通路来实现的[10]。在对肺动脉高压(pulmonaryarterial hypertension, PAH)的研究中,发现miR-181a-5p 通过靶向endocan 改善野百合碱诱导的PAH 的炎症反应[11]。最新的研究发现KLF2 诱导的外泌体miR-181a-5p 可以减弱PAH 的血管重塑[12]。本研究发现低氧诱导能够抑制miR-181a-5p 的表达,同时miR-181a-5p 过表达抑制低氧诱导的肺动脉平滑肌细胞增殖以及迁移,而miR-181a-5p 敲减促进低氧诱导的肺动脉平滑肌细胞增殖以及迁移。以上这些证据提示miR-181a-5p可能是通过抑制肺动脉平滑肌细胞的增殖和迁移,从而来缓解PAH 的病理进展。
由于miRNA 一般是通过靶向mRNA 的3'UTR来负调控基因表达,本研究通过生物信息学预测发现miR-181a-5p 能够与KLF6 3’UTR 区域结合,通过进一步的试验发现miR-181a-5p 能够负向调控肺动脉平滑肌细胞中的KLF6 的表达水平。Krüppel 样因子6(KLF6)是一种转录调节因子;它属于锌指类DNA 结合转录因子的Krüppel 样因子(KLFs)家族,可调节包括细胞增殖、分化、信号转导、肿瘤发生和细胞死亡在内的整体功能。研究发现KLF6 能够介导大鼠实验性肝肺综合征中的肺血管生成[13]。Kim GD 等[14]发现KLF6 能够促进巨噬细胞的炎症反应和低氧反应。研究也发现KLF6 信号通路激活参与糖尿病肺纤维化中的上皮-间质转化[15]。本研究发现KLF6 过表达能够反转miR-181a-5p 过表达对低氧诱导的肺动脉血管平滑肌细胞的增殖和迁移的抑制作用。以上这些结果提示,miR-181a-5p 通过靶向抑制KLF6 的表达,使肺动脉平滑肌细胞增殖和迁移,缓解PAH 的病理进展。本研究仅仅验证KLF6 为miR-181a-5p的一个下游靶点,未对其他靶点进行相关的验证。因此将来有必要对其他的靶点进行相关验证,从而更全面了解miR-181a-5p 在PAH 中的分子调控机制。
综上所述,本研究结果提示miR-181a-5p 能够抑制低氧诱导下的肺动脉平滑肌细胞增殖和迁移,其作用机制可能是通过靶向抑制KLF6 的表达来实现。由于本研究仅仅在细胞层面进行了初步探索,将来需要进一步通过体内试验来验证miR-181a-5p调控PAH 的机制。
