水杨酸缺失型突变体NahG和野生型番茄果实抗冷性的比较
2022-04-01赵瑞瑞孙存一生吉萍
丁 洋,赵瑞瑞,孙存一,申 琳,生吉萍
(1.北京物资学院物流学院,北京市物流系统与技术重点实验室,北京 101149;2.中国农业大学食品科学与营养工程学院,北京 100083;3.中国人民大学农业与农村发展学院,北京 100872)
随着居民生活水平的提高和网络购物的普及,我国生鲜电商行业和冷链物流市场发展迅速[1]。虽然冷链物流能够延缓果蔬采后成熟、抑制病原菌生长、保持果实贮藏品质,但许多热带、亚热带果蔬对低温却相当敏感,若温度控制不当,极易发生冷害[2]。冷害是指果蔬在其冰点以上的不适低温环境中受到的生理伤害,是冷敏型果蔬在逆境胁迫下的一种不良反应[3]。冷敏型果蔬在不适低温下并不会立即产生冷害症状,只有将这些在低温下贮藏的果蔬产品转移至较温暖的环境(20~25 ℃)之后冷害症状才会显现出来[4],而此时的果蔬产品已进入终端消费者手中,严重地影响采后果蔬的食用品质和产品价值,影响到消费者再次购买的意愿。我国是果蔬生产和消费的大国,销售的果蔬中大约有50%是冷敏型果蔬,目前我国每年采后经济损失高达上千亿元,其中由冷害造成的损失占总损失的1/3以上[4]。因此,采后冷害仍是制约我国果蔬产业和生鲜电商平台发展的重大难题。
水杨酸(salicylic acid,SA)作为一种重要的植物内源激素,参与植物体内多种生理生化过程和抗性调控过程[5-6]。Zhang Xinhua等[7]研究发现,外源水杨酸甲酯(methyl salicylate,MeSA)处理樱桃番茄果实可促进内源多胺积累,使冷害症状减轻。任邦来等[8]研究发现,外源SA处理能减少果蔬水分蒸发,降低果实质量损失率,对保持果实硬度和可溶性固形物、VC、可滴定酸含量有较好作用。朱璇等[9]发现,SA处理可以维持细胞器和膜系统的完整性,从而提高杏果实冷藏期间的抗冷性。由此可见,SA在采后果蔬的贮藏保鲜和抗性诱导中均发挥着重要作用。张春光等[10]将细菌中编码水杨酸羟化酶的nahG基因转入烟草和拟南芥后发现,病原物侵染后这两种转基因植物中SA的积累受到了抑制,从而减弱了植株限制病原物扩展和产生系统获得性抗性(system acquired resistance,SAR)的能力。与野生型拟南芥植株相比,NahG突变体更易受臭氧伤害[11]。
番茄作为典型的冷敏型果蔬,是采后果实冷害研究的重要模式材料[7]。目前内源SA在果实抗冷调控中的作用机制及其相关信号网络研究尚未完全清楚,特别是关于低温对SA缺失型番茄果实抗冷性影响的研究甚少。本实验采用NahG(SA缺失型)番茄果实和野生型(wild type,WT)番茄果实为实验材料,于4 ℃下贮藏28 d,分析冷害指数、游离脯氨酸含量及磷脂酶C(phospholipase C,PLC)、磷脂酶D(phospholipase D,PLD)、脂氧合酶(lipoxygenase,LOX)、多聚半乳糖醛酸酶(polygalacturonase,PG)和纤维素酶活力的变化情况以及木质素、纤维素、总酚和儿茶素水平,比较研究冷藏过程中WT和NahG番茄果实的抗冷性变化,为掌握水杨酸在果实抗冷过程中的作用提供理论依据。
1 材料与方法
1.1 材料与试剂
可稳定遗传的NahG番茄(体内不能积累高水平的SA)和野生型(WT)番茄(Solanum lycopersicumcv.moneymaker)种子来源于美国加州大学戴维斯分校的C.M.Rick番茄遗传资源中心。将上述番茄种子播种于自北京金六环农业生态园,于开花后对花期进行标记,绿熟期挑选大小基本相同、果型饱满、无病虫害、无机械伤的果实作为实验材料。
常用试剂和药品均为国产分析纯。
1.2 仪器与设备
TU-1901双光束紫外-可见分光光度计 北京普析通用有限责任公司;HH-4数显恒温水浴锅 金坛市荣华仪器有限责任公司;GL-20G-II型高速离心机 上海安亭科学仪器厂;s210 pH酸度计 瑞士梅特勒-托利多公司;ANKOM 2000i全自动纤维分析仪 北京安科博瑞科技有限公司。
1.3 方法
1.3.1 原料处理
将采摘后的番茄先运至冷库(15 ℃)预冷过夜后,用体积分数3%次氯酸钠溶液表面消毒2 min,然后清水洗净2 次,自然晾干。整个贮藏期间,果实均贮藏于冷库中((4±1)℃、相对湿度85%~90%)。对照组(WT果实)和实验组(NahG果实)各固定90 个果实用于统计冷害指数:分别于14、21、28 d从冷库中各取30 个样品,于室温下放置2 d,用于测定冷害指数。其余果实分别在0、3、7、14、21、28 d取样,每次样品取5 个,取样部位为果实赤道部位1 cm宽的外果皮和果肉,用液氮速冻后放入-80 ℃冰箱中冷冻,待样品全部取完后,用冷冻研磨机研磨,用于测定PLC、PLD、LOX、PG和纤维素酶的活力以及脯氨酸含量、木质素和纤维素质量分数、总酚和儿茶素含量。
1.3.2 冷害指数测定
统计采用Zhao Danying等[12]的方法,每次从各组中随机抽取8 个果实统计冷害级别:0级,冷害面积占比为0;1级,冷害面积占比为0~25%;2级,冷害面积占比为25%~50%;3级,冷害面积占比为50%~75%;4级,冷害面积占比为75%~100%。统计各冷害级别果实个数后,按式(1)计算冷害指数。

1.3.3 游离脯氨酸含量测定
游离脯氨酸含量测定参考曹建康等[13]的方法。实验重复3 次。
1.3.4 LOX活力测定
LOX活力测定参考Minguez-Mosquera等[14]的方法。实验重复3 次。
1.3.5 PLC、PLD活力测定
PLC、PLD活力测定分别参考Mao Linchun[15]和Kurioka[16]等的方法。实验重复3 次。
1.3.6 纤维素酶活力测定
纤维素酶活力测定参考曹建康等[13]的方法。实验重复3 次。
1.3.7 PG活力测定
PG活力测定参考Gayathri等[17]的方法,并略有改动。称取5 g 样品,加入10 mL预冷的体积分数95%乙醇溶液研磨,离心后弃上清液,沉淀中加入5 mL预冷的体积分数80%乙醇溶液,离心后去上清液,沉淀中加入5 mL预冷的提取缓冲液,离心后收集上清液,为酶提取液。加入1.0 mL 50 mmol/L pH 5.5醋酸钠缓冲液和0.5 mL 10 g/L多聚半乳糖醛酸,加入0.5 mL酶提取液,以加入0.5 mL 经煮沸5 min的酶提取液作为对照,37 ℃保温1 h后加入1.5 mL 3,5-二硝基水杨酸试剂,沸水浴5 min,以蒸馏水稀释至25 mL,测定540 nm波长处吸光度。以每克鲜样品每小时分解多聚半乳糖醛酸产生1 mg半乳糖醛酸定义为1 个酶活力单位。实验重复3 次。
1.3.8 木质素质量分数测定
木质素质量分数测定参考范鹏程等[18]的方法,略有改动。取0.1 g样品加入10 mL体积分数1%乙酸溶液,离心后弃上清液,加入5 mL体积分数1%乙酸溶液洗涤沉淀。再加入4 mL乙醇和乙醚混合液(体积比1∶1)浸泡沉淀3 min,弃上清液,重复3 次,沸水中蒸干沉淀,加入3 mL质量分数72%硫酸溶液,静置16 h以溶解全部纤维素,加入10 mL蒸馏水,沸水加热5 min,冷却后加入5 mL蒸馏水和0.5 mL质量分数10%氯化钡溶液,摇匀离心后弃上清液。沉淀中加入10 mL体积分数10%硫酸溶液和10 mL 0.025 mol/L重铬酸钾溶液,沸水加热15 min。冷却后转入烧杯,用20 mL蒸馏水洗涤残余部分后也转入烧杯。在烧杯中先加入5 mL质量分数20%碘化钾溶液和1 mL质量分数1%淀粉溶液,再用0.2 mol/L硫代硫酸钠溶液滴定,记录颜色变化时所用的硫代硫酸钠用量以计算木质素含量。实验重复3 次。
1.3.9 纤维素质量分数测定
纤维素质量分数测定采用全自动纤维分析系统。称取样品1 g(m/g)放置在坩埚中热浸提5 min,结束后关掉加热开关,用样品夹转移到冷浸提装置。将坩埚在(105±2)℃下处理5 h,冷却坩埚到室温(最好放在干燥器中)称质量(m1/g,精确到0.1 mg),灰化样品在(525±10)℃条件下处理3 h;冷却坩埚到室温,称质量(m2/g,精确到0.1 mg)。实验重复3 次。按式(2)计算纤维素质量分数。
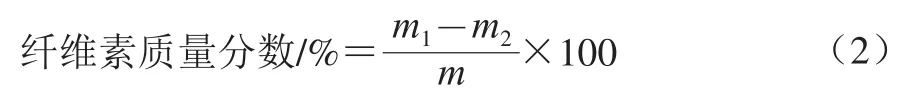
1.3.10 总酚和儿茶素含量测定
总酚含量测定参考Pirie等[19]的方法。儿茶素含量测定参考胡凤等[20]的方法。实验重复3 次。
1.4 数据处理与统计分析
采用Excel 2010软件对数据进行统计分析,采用Origin 10.0软件制图。采用SPSS 17.0软件进行差异显著性分析,具体采用独立样本T检验,当P<0.05时,认为两对比组之间具有显著性差异。
2 结果与分析
2.1 NahG和WT番茄果实的冷害情况分析
如图1A所示,NahG果实和WT果实在4 ℃下贮藏14 d并转移至室温下放置2 d后均出现冷害症状。之后,随着贮藏时间延长,两组果实冷害指数均呈上升趋势。到贮藏28 d时,NahG果实冷害指数显著低于WT果实,比WT果实降低了23.53%(P<0.05)。如图1B所示,冷藏28 d后,NahG果实和WT果实移至室温放置2 d,NahG果实的冷害症状比WT果实明显减轻。
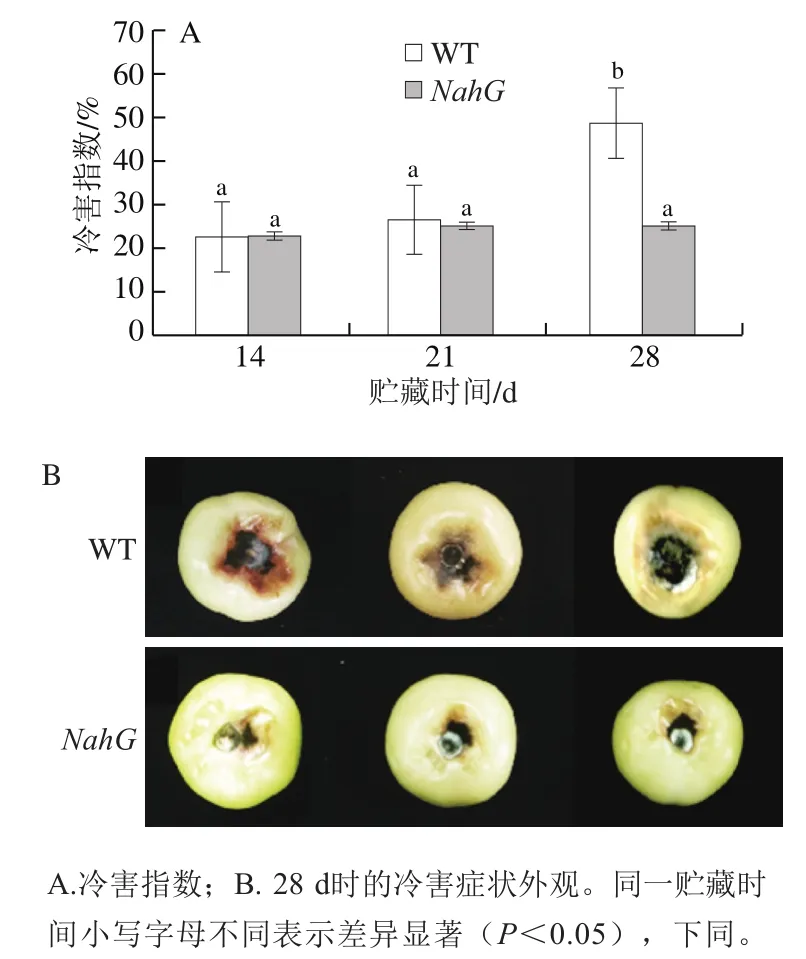
图1 冷藏过程中NahG和WT番茄果实冷害指数及冷害症状的变化Fig. 1 Changes in chilling injury index and symptoms in NahG and WT tomato fruits during cold storage
2.2 NahG和WT番茄果实的游离脯氨酸含量比较
受到外界环境胁迫时,植物组织中游离脯氨酸含量会明显升高,这可以增强细胞的渗透性、稳定原生质胶体及组织内的代谢、降低冰点,对提高低温胁迫下果蔬组织的抗逆性、避免细胞遭到氧化胁迫的伤害等方面有十分重要的作用[21]。如图2所示,在贮藏期间WT与NahG果实中游离脯氨酸含量不断增加,NahG果实中游离脯氨酸含量显著高于WT果实(P<0.05)。贮藏14 d后,NahG果实游离脯氨酸含量快速升高,到28 d达到最大值(22.15 μg/g),而WT果实仅为8.06 μg/g。这说明NahG果实抗冷能力比WT果实更强。
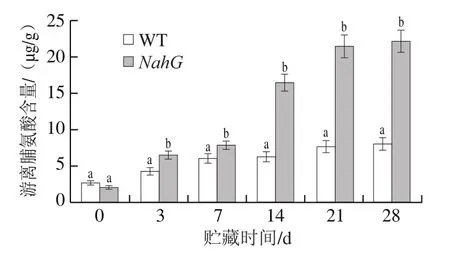
图2 冷藏过程中NahG和WT番茄果实游离脯氨酸含量的变化Fig. 2 Changes in proline content in NahG and WT tomato fruits during cold storage
2.3 NahG和WT番茄果实的细胞膜相关酶活力比较
脂类是生物膜的重要组成成分,其不饱和程度与生物膜的稳定性及流动性密切相关,不饱和脂肪酸的水平是影响膜流动性的主要因素[22]。PLC和PLD为磷酸二脂酶,能够水解磷酸二脂键,催化多不饱和脂肪酸的过氧化,对细胞的结构和稳定性有非常重要的作用,是造成膜损伤和发生冷害的主要物质[23]。由图3A可以看出,WT果实中PLC活力在整个贮藏期间都显著高于NahG果实(P<0.05)。在贮藏期间WT果实中PLC活力呈先下降后增加的变化趋势,而NahG果实PLC活力是先下降后上升再下降,到贮藏第7天达到最大值(127 U/kg),此时WT果实PLC活力仍显著高于NahG果实。由图3B看出,NahG与WT果实PLD活力变化趋势一致,均在第7天时达到最小值,除第7天外,其余时间WT果实PLD活力均显著高于NahG果实(P<0.05)。以上结果说明,在低温胁迫下,NahG果实PLC和PLD活力的增加受到明显抑制。
LOX是酶促脂质过氧化的重要因子,能催化游离的不饱和脂肪酸产生脂质过氧化自由基,这些自由基能直接作用于膜磷脂中的结合态不饱和脂肪酸,导致细胞膜磷脂双分子层破坏,与后熟衰老过程的启动和逆境胁迫、伤诱导、病原信号的产生和识别等关系密切[23]。如图3C所示,在贮藏过程中,NahG和WT果实LOX活力呈先上升后降低趋势。贮藏14 d时,NahG果实LOX活力显著低于WT果实(P<0.05)。贮藏第3天,NahG果实LOX活力达到最大值(4.24 U/(g·min)),WT果实在第14天时LOX活力达到最大值(7.59 U/(g·min)),比同期NahG果实高86.95%。实验结果表明,NahG果实冷害症状的发展被延缓,同时PLC、PLD和LOX的活力受到抑制,保持了细胞膜的相对稳定。
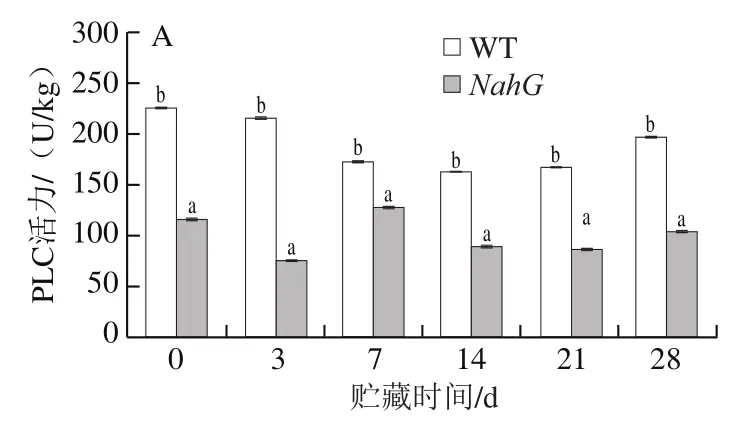
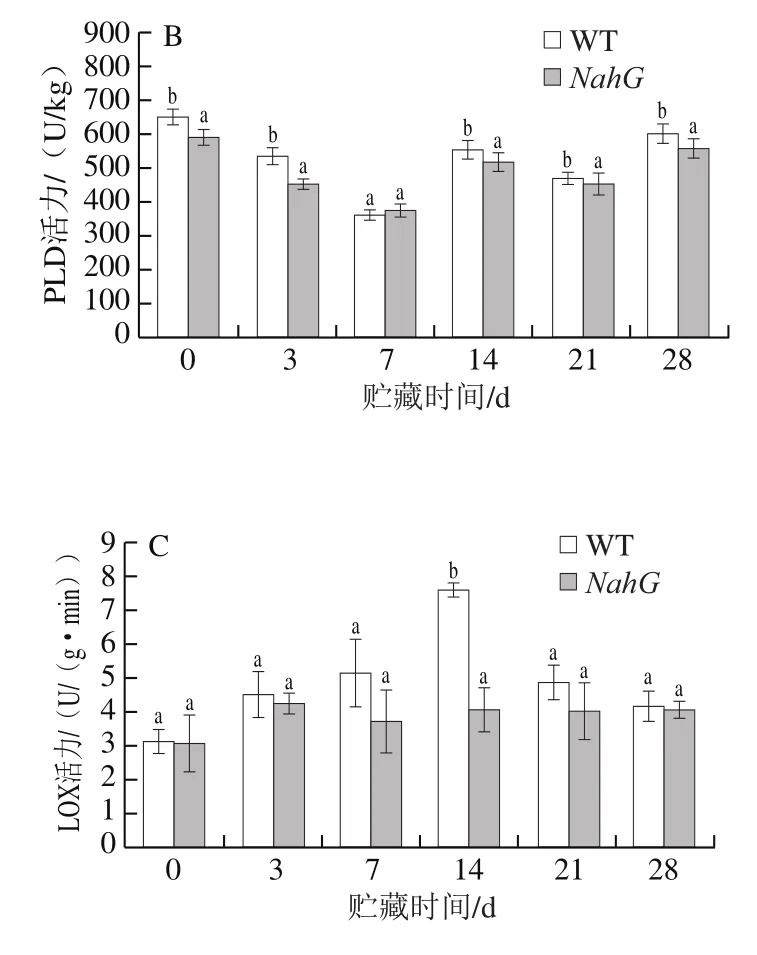
图3 冷藏过程中NahG 和WT番茄果实PLC(A)、PLD(B)和LOX(C)活力的变化Fig. 3 Changes in activities of PLC (A), PLD (B) and LOX (C) in NahG and WT tomato fruits during cold storage
2.4 NahG和WT番茄果实的细胞壁相关酶活力比较
在低温胁迫下,采后果实中的PG、纤维素酶、半乳糖苷酶等一系列水解酶会将细胞壁中的纤维素、半纤维素、原果胶、木质素等物质分解,使细胞壁解体,果蔬组织变软,导致细胞壁代谢异常,发生冷害[24]。如图4A所示,在贮藏期间除了0、14 d和28 d外,NahG果实PG活力显著低于WT果实(P<0.05)。WT果实的PG活力在贮藏第7天迅速增加到13.53 mg/(kg·h),而NahG果实此时仅为3.18 mg/(kg·h)。到贮藏第14天,NahG果实PG活力达到最大。如图4B所示,WT果实中纤维素酶活力在贮藏14 d时达到最大值(6.32 mg/(kg·h)),此时可能番茄果实细胞壁的结构已经发生改变,而NahG果实中纤维素酶活力在28 d时才达到最大(4.42 mg/(kg·h)),在整个贮藏期,除第28天外,WT果实中纤维素酶活力显著高于NahG果实(P<0.05)。实验结果表明,与WT果实相比,NahG果实中不但PG的活力显著降低,且PG和纤维素酶活力峰值出现时间也被推迟。
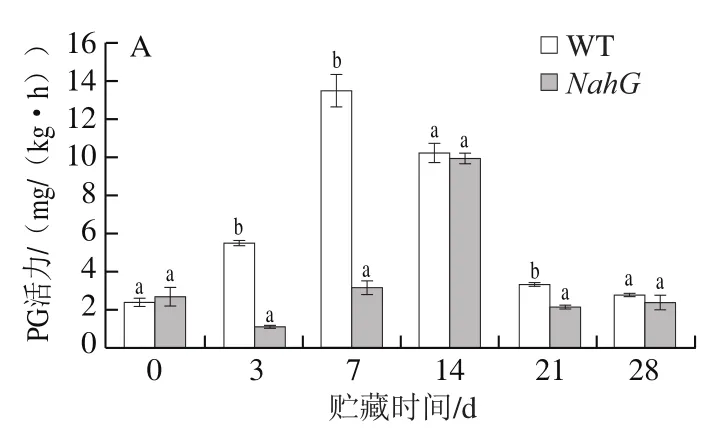
2.5 NahG和WT番茄果实的木质素和纤维素质量分数比较
木质素是植物细胞壁的主要成分之一,与纤维素协同作用可增强果实的表皮结构,提高植物抗病性[25]。如图5A所示,在贮藏第3天和第21天,NahG果实中的木质素质量分数显著高于WT果实(P<0.05)。WT果实中木质素质量分数在贮藏14 d之前变化幅度不大,到21 d时降低到2.80%;而NahG果实木质素质量分数在贮藏第3天就增加到最大(4.18%),后期贮藏过程中变化幅度不大。与WT果实相比,NahG果实的木质素质量分数更高,可推迟果实软化。
植物细胞壁结构的改变与纤维素、半纤维素的结构变化和果胶物质的降解有关。纤维素可以支持和保护细胞骨架,防止植物组织变软[26]。由图5B可以看出,在整个贮藏期间,NahG果实与WT果实纤维素质量分数变化趋势相似,均在贮藏第14天时达到最大值,此后均呈降低趋势,结合冷害指数的实验结果,可以得出结论:贮藏14 d以后番茄果实出现冷害症状,随时间推移,冷害发展严重,品质下降。与WT果实相比,NahG果实能有效提高细胞壁的稳定性。
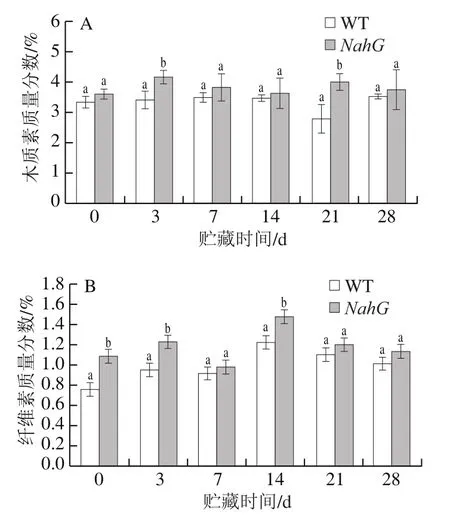
图5 冷藏过程中NahG和WT番茄果实木质素(A)和纤维素(B)质量分数的变化Fig. 5 Changes in contents of lignin (A) and cellulose (B) in NahG and WT tomato fruits during cold storage
2.6 NahG和WT番茄果实的总酚和儿茶素含量比较
酚类化合物是植物重要的次级代谢产物之一。果蔬色泽、品质和味道的形成、成熟与衰老进程、组织色变、抗逆性抗病性代谢等都与酚类有很大的关系[27]。由图6A可以看出,在贮藏初期,两组番茄果实中的总酚含量相差不大。第7天时,NahG果实中总酚含量迅速增加,从贮藏第7天开始,NahG果实中的总酚含量显著高于WT果实(P<0.05)。在贮藏0~21 d,两组番茄果实中总酚含量在贮藏期间不断增加,而21~28 d总酚含量下降,可能到第28天时两组番茄果实都发生严重冷害,导致果实中总酚物质含量减少,但NahG果实中总酚含量仍高于WT果实。实验结果表明,NahG果实在低温胁迫下能够保持酚类物质含量处于较高水平,抑制酚类物质进一步氧化,从而减轻番茄组织褐变等冷害症状。
儿茶素可以清除自由基,具有强抗氧化作用,并能够调控植物的抗性[28]。如图6B所示,在贮藏期间,NahG果实中的儿茶素含量显著高于WT果实(P<0.05)。在21 d前,两组儿茶素的含量不断增加,到21 d时达到最大值,此时NahG果实儿茶素含量为8.29 μg/g,比WT果实高27.93%。实验结果提示,NahG番茄果实内的抗氧化物质总酚和儿茶素的含量均高于WT果实,表明NahG果实抗冷和清除自由基的能力增强。
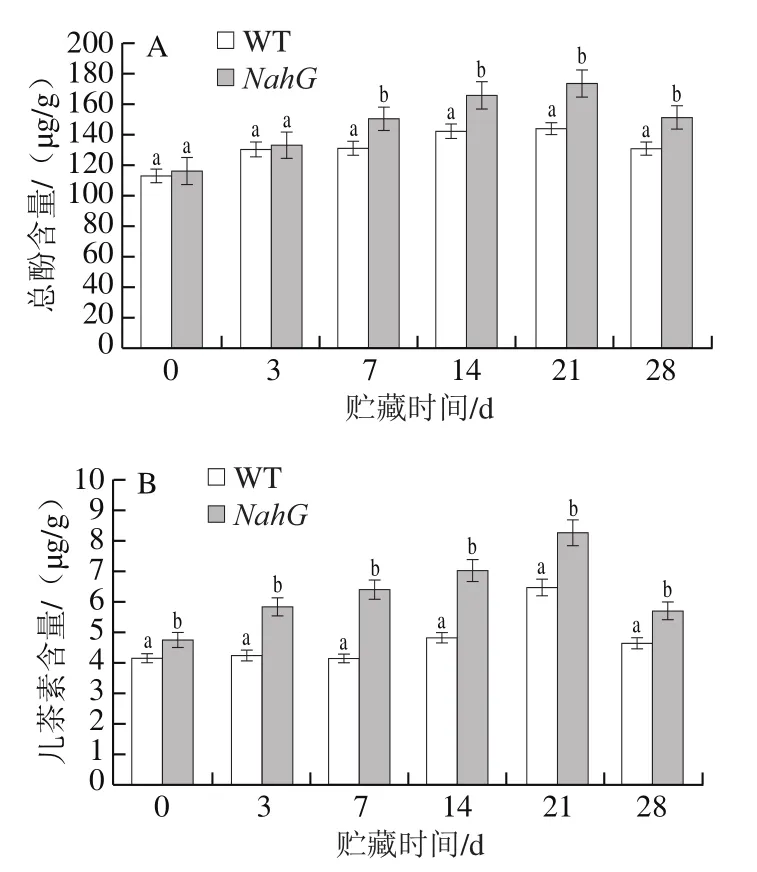
图6 冷藏过程中NahG和WT番茄果实总酚(A)和儿茶素(B)含量的变化Fig. 6 Changes in contents of total phenolic compounds (A) and catechin (B) in NahG and WT tomato fruits during cold storage
3 讨 论
SA是植物中非常重要的内源激素类物质和信号分子,不仅参与植物的抗病过程,还参与植物的抗冷反应。在番茄果实中施用低浓度的MeSA可以减轻低温贮藏过程中的冷害和腐烂发生率[29]。NahG番茄是从细菌中转入了一个编码SA羟化酶的NahG基因[30]。该酶能分解植物体内产生的SA,产生儿茶素,使体内不能积累高水平的SA。本实验中的NahG突变体番茄果实中儿茶素的含量显著高于野生型(P<0.05),其抗冷能力也显著提高。这与拟南芥NahG突变体植株抗冷能力高于野生型植株[31]的结果一致。在低温胁迫下,SA积累过高促进了拟南芥的膜脂过氧化,而不能积累高水平SA的NahG突变体生长状况却明显好于野生型,表明SA高浓度积累不利于拟南芥对低温胁迫的耐受[32]。在果实采后贮藏中的研究表明,外源SA诱导的抗冷性存在浓度依赖效应。陈国品等[33]分别用0.04、0.36、0.72、1.08、1.44 mmol/L的SA溶液喷洒巨峰葡萄叶片,在自然低温条件下(<10 ℃),1.08 mmol/L的SA对冬葡萄果实的抗冷性有更好的促进作用。蔡琰[34]采用1.0 mmol/L SA处理水蜜桃,比0.7 mmol/L和1.3 mmol/L SA处理在0 ℃下贮藏能更好地抑制后熟软化和果肉褐变,显著抑制冷害的发生。尽管外源SA处理能够有效提高采后果实的抗冷性[7,9,33-34],但关于内源SA的水平对采后果实抗冷性影响的研究甚少且机制尚未明确。
低温通过改变细胞内丙二醛和活性氧含量对植物造成严重损伤[35]。低温最先损伤的是细胞膜,而细胞膜结构和功能的完整性对于低温下植物维持细胞代谢和生长必不可少。脂质成分的改变与否决定了低温下细胞膜是否受损。在许多植物中,细胞膜中脂肪酸不饱和度和不饱和磷脂酰胆碱比例的增加对提高植物抗冷能力起至关重要的作用[36]。磷脂是生物膜的骨架成分,PLC和PLD参与磷脂分子的水解反应[37],在持续低温条件下,植物中PLD会大量水解磷脂,引起细胞膜不可逆的损伤[38]。LOX具有膜脂过氧化的作用,其活性的升高会引起质膜的降解和过氧化物的积累[39]。PG和纤维素酶分别调节果胶和纤维素的降解,是引起细胞壁降解并最终导致果实软化的主要参与酶类[40]。在本实验中,NahG突变体番茄果实的PLC、PLD、PG和纤维素酶活性低于WT果实,木质素和纤维素质量分数高于WT果实,进一步说明转入NahG基因有效抑制了细胞内磷脂、果胶和纤维素的降解,保护了细胞膜和细胞壁结构,防止了果实的软化,进而提高了果实自身的抗冷能力。nahG编码的SA羟化酶能够水解SA,形成具有抗氧化能力的儿茶素。NahG突变体果实中总酚和儿茶素含量的增加使得细胞内抗氧物的提高,从而抑制细胞成分的氧化,对提高果实的抗冷性起到促进作用。
4 结 论
本实验将NahG突变体和野生型番茄果实进行4 ℃下28 d的低温贮藏,对贮藏期间果实的冷害症状、冷害指数,细胞膜和细胞壁相关酶活力,以及游离脯氨酸、总酚、儿茶素含量及木质素、纤维素质量分数进行综合分析。结果表明,NahG番茄果实具有更强的抗冷性,其原因可能是SA水解影响低温下果实中抗氧化物质总酚和儿茶素的水平以及细胞膜和细胞壁降解酶系统活性,促使番茄果实的抗冷能力增强。
