内源异黄酮对紫花苜蓿结瘤固氮及氮效率的调控研究
2022-03-26童长春刘晓静吴勇赵雅姣王静
童长春,刘晓静,吴勇,赵雅姣,王静
(甘肃农业大学草业学院,草业生态系统教育部重点实验室,甘肃省草业工程实验室,中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
黄酮类化合物是广泛存在于自然界的一大类化合物,具有较强的生物活性和生理作用,按结构可将黄酮类化合物分为黄酮类、黄酮醇类、二氢黄酮类、二氢黄酮醇类、查尔酮类、双黄酮类、异黄酮类以及其他黄酮类等,其中异黄酮(isoflavonoes)是植物苯丙氨酸代谢过程中的一种以C6-C3-C6结构为母核的杂环多酚类次生代谢产物,在植物界集中分布在蝶型花科的一些物种中[1]。研究表明,异黄酮在植物中主要行使两方面功能,一种是作为植物保护素,用于抵御病原物和疾病,来增强植物的抗性[2]。植物在低肥、金属毒害等逆境胁迫或环境的改变时能够诱导异黄酮基因的表达,并通过自身的调节机制来增强对外界各种恶劣环境的适应性[3]。另一种功能是异黄酮作为豆科植物与根瘤菌之间的信号分子:有研究表明,根瘤菌和豆科植物的共生固氮过程中,植物一方面通过分泌黄酮类和异黄酮类化合物至根际,吸引固氮菌和其他微生物靠近植物的根部,诱导植物根圈根瘤菌结瘤因子的形成[4-5];另一方面通过增加体内异黄酮含量,可以抑制植物生长素的运输,导致植物生长素在结瘤起始部位积聚,从而刺激细胞分裂和结瘤器官发生[6]。因此,异黄酮对豆科植物的氮素利用至关重要。
对植物氮素营养特性的不断探索发现,同一种植物的不同基因型品种对氮素的敏感度和响应程度存在一定差异,即氮效率差异[7]。氮高效作物品种能够积累更多的氮素,从而促进光合作用以及提高碳、氮同化效率,最终得到高产[8-9]。由于豆科根瘤的固氮作用,其氮素营养特性与其他植物不同,外源氮素添加会对豆科植物结瘤固氮产生复杂的影响[10],但同属的豆科植物不同基因型品种间对氮素的反应也会不尽相同,并且可以根据其氮含量、氮素积累量和产量等将品种划分为不同的氮效率类型,同一生长条件下,氮高效型品种的氮含量、氮素积累量和产量均显著高于氮低效型品种[11]。紫花苜蓿(Medicago sativa)作为优质蛋白饲草,在畜牧业发展中的重要性日益突显,并且紫花苜蓿还能肥田沃土,随着我国种植业结构的调整,其栽培面积逐年增加[12-13]。本团队前期的研究发现:不同品种紫花苜蓿的氮素吸收、累积、运输和再利用能力在品种间均具有差异[14],从而影响产量的高低[15]。
近年来,提高植物氮肥利用率受到国内外学者的广泛关注,充分挖掘植物对氮素利用的遗传潜力成为提高作物产量和品质以及合理利用资源、提高氮素利用效率的理想途径。培育氮高效紫花苜蓿品种能满足我国草食家畜和奶牛饲养业快速发展的需求[16],对紫花苜蓿体内异黄酮与结瘤固氮的研究将是实现这一目标的突破口。然而目前为止,对于有关异黄酮和结瘤的研究主要集中在豆科植物结瘤过程中异黄酮的必要性上[17],而在不同水平外源氮素供应时,异黄酮是否对豆科植物的结瘤固氮及氮效率具有一定的调控作用,并且如何参与这一过程仍有待明确。因此,本研究通过对不同氮效率紫花苜蓿的异黄酮含量和结瘤固氮以及异黄酮合酶(isoflavone synthase,IFS)和结瘤信号传递通路相关基因(nod)表达的研究,从而探讨紫花苜蓿体内异黄酮与结瘤固氮的相关性并揭示紫花苜蓿体内异黄酮对其氮效率的调控机制,为今后培育氮高效型紫花苜蓿品种提供理论基础。
1 材料与方法
1.1 试验材料
供试紫花苜蓿品种为“LW6010”(M. sativacv. Longwei 6010)和陇东苜蓿(M. sativacv. Longdong),来源于甘肃农业大学草业学院。郝凤[18]研究发现“LW6010”紫花苜蓿对氮素的吸收利用效率较高,陇东苜蓿对氮素的吸收利用效率较低。
1.2 试验设计
试验采用营养液砂培,在培养室内进行(光照28 ℃/14 h,黑暗20 ℃/10 h,光照强度260~350 μmol·m-2·s-1,相对湿度60%~70%)。选用饱满均一的紫花苜蓿种子,经消毒后于2019 年3 月1 日播种至直径9 cm、高12 cm装有灭菌砂的培养杯中,每6 杯放入1 个水培盒。设置5 个氮素(N)水平:21(N21)、105(N105)、210(N210)、315(N315)、420(N420)mg·L-1(氮素水平的设定参考前期相关试验[19]),共10 个处理,每个处理重复3 次。用蒸馏水培养至幼苗3 cm 高时进行间苗,每杯保留20 株健壮幼苗。长至2 叶1 心时,以Hoagland-Arnon 营养液[20]为基本营养液,NO3--N∶NH4+-N=1∶1 为氮源,调节营养液pH 为7,每个水培盒浇入1 L 营养液,进行不同水平的供氮处理。之后每周淋洗1 次防止盐分积累,再重新浇入新的氮营养液,期间每天补蒸馏水至第一次加营养液的高度。长至3 片复叶时,每杯接种吸光值为0.6 的根瘤菌悬液10 mL。培养60 d 后各处理间生长性状表现明显差异时取样进行相关指标的测定。
1.3 测定指标及方法
1.3.1 异黄酮含量的测定 采用高效液相色谱法(high performance liquid chromatography,HPLC)测定不同氮效率紫花苜蓿的异黄酮含量:称取干草样品1 g(粉碎过1 mm 筛),以料液比1∶20(m:v)加入色谱级甲醇20 mL,浸泡4 h 后,超声提取30 min,再加入甲醇20 mL 超声30 min,提取2 次,合并2 次提取液,30 ℃下真空旋干,将粗提物溶于甲醇至10 mL,用0.22 μm PTFE 滤膜过滤后可用于高效液相色谱分析,色谱条件:分离柱为C18 反相柱;进样量10 μL;流动相为乙腈∶0.3% H3PO4=20%∶80%(0~5 min)→40%∶60%(5~15 min)→60%∶40%(15~25 min)→80%∶20%(25~30 min);流速0.8 mL·min-1;柱温为室温;分析时间30 min;检测波长260 nm。异黄酮标样见图1。

图1 异黄酮标样色谱图Fig.1 Chromatogram of isoflavone standard sample
1.3.2 相关基因的表达 于NCBI(national center for biotechnology information)网站查找筛选与蒺藜苜蓿(Medicago truncatula)异黄酮合酶(IFS)及结瘤信号传递通路相关的外显子基因(nod)序列,利用Premier 5.0 进行结瘤及异黄酮相关基因引物设计,序列信息如表1,引物送由上海生物工程公司合成。

表1 定量PCR 引物序列信息Table 1 Information of primer sequences of quantitative real-time PCR
紫花苜蓿RNA 提取采用RNA prep Pure 试剂盒,试剂盒由天根生化科技有限公司提供,提取步骤参照说明书。采用1%琼脂糖凝胶电泳法和超微量紫外分光光度计(NanoPro 2010,北京鼎昊源科技有限公司)来进行浓度和纯度的检测。
利用设计的目标基因引物对提取的紫花苜蓿RNA 进行反转录合成cDNA,对合成的cDNA 模板进行PCR 扩增,PCR 扩增体系及反应条件均参考试剂盒,通过荧光定量对目标基因表达差异进行分析。试剂盒均由南京诺唯赞生物科技有限公司提供。
1.3.3 根瘤参数的测定 将植株冲洗干净后统计所有根瘤数量,计算单株根瘤总数;每株紫花苜蓿粉色及浅粉色根瘤(主要是豆血红蛋白的颜色)数量为有效根瘤数[21];统计完所有根瘤数量后用分析天平进行称重,计算单株根瘤总鲜重(mg·plant-1);单根瘤重(mg)为单株紫花苜蓿根瘤重与总根瘤数的比;采用乙炔还原法测定固氮酶活性(μmol·g-1·h-1)[22];单株固氮潜力(μmol·h-1)为固氮酶活性与单株总根瘤重的乘积。
1.3.4 氮积累量的测定 采用凯氏定氮法测定含氮量[23]。按以下公式进行相关指标计算:

1.4 数据统计与分析
采用Excel 2010 软件整理数据,应用SPSS 19.0 软件对数据进行统计分析。采用2-⊿⊿CT法[24]进行荧光定量PCR 数据的处理与分析。
2 结果与分析
2.1 紫花苜蓿的异黄酮含量及相关基因的表达
2.1.1 异黄酮含量 紫花苜蓿根系中的异黄酮含量高于茎叶,其中染料木素和大豆苷元的含量高于木犀草素和刺芒柄花素(表2)。随着氮素水平的升高,“LW6010”紫花苜蓿茎叶和根系中的异黄酮含量均表现出先下降后上升的趋势,除了茎叶中的N21和N420处理差异不显著外(P>0.05),茎叶和根系中的总异黄酮含量均在N21处理下具有最大值。陇东苜蓿茎叶中,除了木犀草素含量随着氮素水平的升高表现出先下降后上升的趋势外其他异黄酮含量的变化规律不明显,其中总异黄酮含量在N21处理下具有最大值,但和N105与N210处理差异不显著(P>0.05)。陇东苜蓿根系中,木犀草素、大豆苷元以及刺芒柄花素含量随着氮素水平的升高表现出先下降后上升的趋势,并且在N420处理下具有最大值,但木犀草素和大豆苷元含量在N21和N420处理下差异不显著(P>0.05),总异黄酮含量在N21处理下具有最大值,但和N210处理差异不显著(P>0.05)。从品种间来看,不同品种紫花苜蓿的异黄酮含量也不同,茎叶中,“LW6010”的异黄酮含量在氮素水平低于N210处理时显著高于陇东苜蓿(P<0.05);根系中,各氮素处理下“LW6010”紫花苜蓿的异黄酮含量均显著高于陇东苜蓿(P<0.05)。

表2 紫花苜蓿的异黄酮含量Table 2 Isoflavone content of alfalfa(µg·g-1)
2.1.2 异黄酮相关基因的表达 “LW6010”在氮素水平低于N210时,茎叶中的IFS基因以及根系中IFS-2和IFS-3基因的表达均显著上调(P<0.05)(图2),在氮素水平高于N210时,除了根系中IFS-3在N210和N315处理下差异不显著外(P>0.05),茎叶和根系中IFS-2和IFS-3基因的表达量均显著上调(P<0.05);陇东苜蓿在氮素水平低于N210时,只有IFS-2基因的表达显著上调(P<0.05),在氮素水平高于N210时,除了茎叶中IFS-2 在N210和N315处理下差异不显著外(P>0.05),茎叶和根系中的IFS-2和IFS-3基因的表达量均显著上调(P<0.05)。整体来看,“LW6010”在氮素水平低于和高于N210时比陇东苜蓿上调的IFS基因更多,并且表达量更高。

图2 紫花苜蓿异黄酮基因的相对表达量Fig.2 Relative expression level of isoflavone genes in alfalfa
2.2 紫花苜蓿的氮素固定
2.2.1 根瘤数 随着氮素水平的升高,“LW6010”的单株总根瘤数呈先下降后上升再趋于平缓的趋势(表3),单株有效根瘤数表现出先下降后趋于平缓的趋势,有效根瘤数/总根瘤数的变化规律则不明显。除了有效根瘤数/总根瘤数在N21和N210处理下的差异不显著外(P>0.05),“LW6010”的单株总根瘤数、单株有效根瘤数以及有效根瘤数/总根瘤数均在N21处理下具有最大值。陇东苜蓿除了N21处理下单株总根瘤数显著大于其他处理而有效根瘤数/总根瘤数显著小于其他处理外(P<0.05),其他氮处理下的单株总根瘤数、单株有效根瘤数以及有效根瘤数/总根瘤数均无明显变化。整体来看,各氮水平处理下,“LW6010”的单株总根瘤数、单株有效根瘤数以及有效根瘤数/总根瘤数均显著高于陇东苜蓿(P<0.05)。
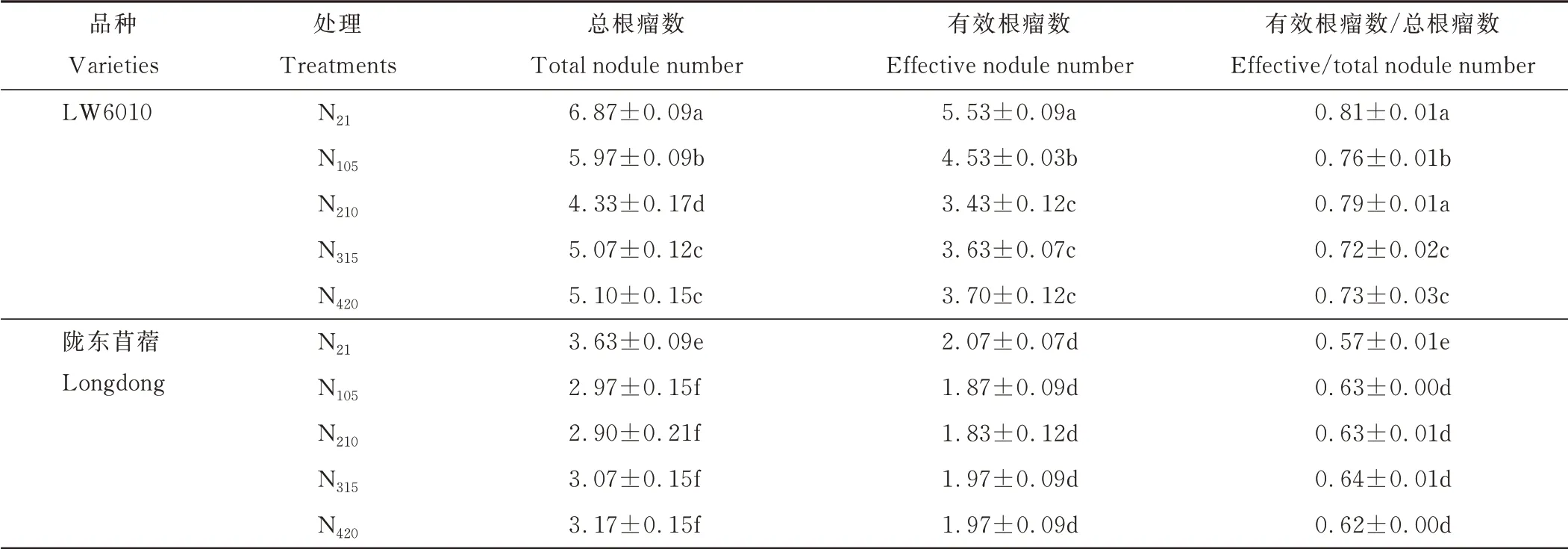
表3 单株紫花苜蓿的根瘤数Table 3 Number of nodules of single alfalfa
2.2.2 根瘤重 随着氮素水平的升高,“LW6010”的单株总根瘤重表现出先下降后上升再趋于平缓的趋势(图3),并且在N21处理下具有最大值。陇东苜蓿的单株总根瘤重除了N21处理下显著高于其他处理外(P<0.05),其他氮处理之间则无明显变化。从品种间来看,除了N210处理,其他氮处理下“LW6010”的单株总根瘤重均显著高于陇东苜蓿(P<0.05)。随着氮素水平的升高,“LW6010”和陇东苜蓿的单根瘤重均无明显变化,从品种间来看,除了N315处理,其他氮处理下陇东苜蓿的单根瘤重均显著高于“LW6010”(P<0.05)。

图3 紫花苜蓿的根瘤重Fig.3 The weight of nodules of alfalfa
2.2.3 固氮酶活性与单株固氮潜力 随着氮素水平的升高,“LW6010”和陇东苜蓿的固氮酶活性均表现为先升高后降低的趋势(图4),“LW6010”在N210处理下最大,但和N315处理差异不显著(P>0.05),陇东苜蓿在N315处理下最大,但和N105、N210处理差异不显著(P>0.05)。“LW6010”和陇东苜蓿的单株固氮潜力对氮素的响应表现出明显差异。随着氮素水平的升高,氮高效型的“LW6010”紫花苜蓿的单株固氮潜力表现出先下降后上升再趋于平缓的趋势,陇东苜蓿的单株固氮潜力表现出下降的趋势,“LW6010”和陇东苜蓿均在N21处理下具有最大单株固氮潜力。从品种间来看,“LW6010”的单株固氮潜力在各氮素水平下整体显著高于陇东苜蓿(P<0.05)。

图4 紫花苜蓿的固氮酶活性与单株固氮潜力Fig.4 Nitrogenase activity and nitrogen fixation potential of alfalfa
2.2.4 氮积累量 随着氮素水平的升高,“LW6010”和陇东苜蓿的地上和地下氮积累量均表现为先升高后降低的趋势(图5),并且均在N210处理下最大。从品种间来看,除了N420处理下“LW6010”和陇东苜蓿的地下氮积累量差异不显著(P>0.05)外,“LW6010”的地上和地下氮积累量在各氮素水平下整体显著高于陇东苜蓿(P<0.05)。

图5 紫花苜蓿的氮积累量Fig.5 Nitrogen accumulation of alfalfa
2.2.5 结瘤相关基因的表达 “LW6010”紫花苜蓿在氮素水平低于N210时,除了茎叶中nod-2在N105和N210处理下差异不显著外(P>0.05),茎叶中的nod基因和根系中nod-1基因的表达均表现出显著上调(P<0.05)(图6),在氮素水平高于N210时,除了根系中nod-1在N210和N315处理下差异不显著外(P>0.05),茎叶中的nod基因和根系中nod-1基因的表达也均显著上调(P<0.05);陇东苜蓿茎叶中,只有nod-1基因的表达在氮素水平为N21时表现出显著上调(P<0.05),根系中,nod-1基因表达在氮素水平为N105和N420时显著上调(P<0.05),nod-2基因表达在氮素水平为N21时显著上调(P<0.05)。整体来看,“LW6010”紫花苜蓿在氮素水平低于和高于N210时比陇东苜蓿表达量上调的nod基因数量更多,并且表达量更高。

图6 紫花苜蓿结瘤基因的相对表达量Fig.6 Relative expression level of nodulation genes in alfalfa
2.3 紫花苜蓿的异黄酮与氮素固定的相关性
2.3.1 异黄酮含量与氮素固定的相关性 紫花苜蓿除了茎叶中的木犀草素与单株有效根瘤数、有效根瘤数/总根瘤数、单株固氮潜力以及刺芒柄花素与有效根瘤数/总根瘤数的相关性不显著外,其异黄酮含量与单株总根瘤数、单株有效根瘤数、有效根瘤数/总根瘤数、单株总根瘤重以及单株固氮潜力均具有显著或极显著正相关关系(表4)。另外,紫花苜蓿茎叶和根系中的大豆苷元、总异黄酮含量以及根系中的刺芒柄花素与单根瘤重呈显著或极显著负相关,可见紫花苜蓿的异黄酮含量主要通过提高其根瘤数量从而促进固氮潜力,但不利于单根瘤重的增加。
2.3.2 异黄酮基因与结瘤基因表达的相关性 紫花苜蓿茎叶中IFS基因的表达与茎叶和根系中nod-1基因的表达具有显著或极显著正相关关系(表5),并且茎叶中IFS-2基因的表达还与茎叶中nod-2基因的表达具有极显著正相关关系;根系中,除了IFS-3基因的表达与茎叶中nod-1基因表达的相关性不显著外,IFS-2、IFS-3基因的表达与茎叶中nod基因以及根系中nod-1基因的表达均呈现显著或极显著正相关。可见,紫花苜蓿主要通过茎叶中IFS基因以及根系中IFS-2、IFS-3基因的表达来共同促进nod基因的表达。

表5 紫花苜蓿异黄酮基因与结瘤基因表达的相关性Table 5 Correlation between isoflavone gene and nodulation gene expression in alfalfa
3 讨论
3.1 紫花苜蓿体内异黄酮与结瘤固氮
豆科植物在与根瘤菌的共生互作过程中,能够从共生体系中获得化合态的氮素维持生命活动[25],因此结瘤固氮对豆科植物至关重要。根瘤菌与豆科植物的共生固氮可分为两个过程:一个是结瘤过程,另一个是固氮过程,前者是后者的先决条件。在结瘤的起始阶段,当植物萌发和根尖穿过土壤时,根际就开始了根与细菌的信号交换,土壤细菌被存在于根分泌物中的多种化合物所吸引,而向根部迁移,相关研究表明绝大多数诱导物属于黄酮类化合物[26]。本研究中,在氮素水平低于210 mg·L-1时紫花苜蓿的异黄酮含量以及根瘤数、根瘤重和单株固氮潜力均表现出上升的趋势,并且相关性分析表明紫花苜蓿的异黄酮含量与其单株总根瘤数、单株有效根瘤数、有效根瘤数/总根瘤数、单株总根瘤重以及单株固氮潜力均具有显著或极显著正相关关系。有研究表明,在根瘤菌感染豆科植物的过程中,植物也可以通过增加体内异黄酮含量来抑制植物生长素的运输使其在结瘤起始部位积聚,从而刺激细胞分裂和结瘤器官发生[6]。可见,紫花苜蓿的结瘤固氮受异黄酮含量的调节,在外源氮素缺乏时紫花苜蓿能通过提高体内异黄酮含量来促进结瘤固氮,从而维持正常的生长发育。另外,本研究还发现,紫花苜蓿的异黄酮含量虽然可以提高总的结瘤数量以及总根瘤重,但是对单根瘤重和固氮酶活性没有明显的提高效果。因此,异黄酮只是通过促进紫花苜蓿的结瘤来提高单株固氮潜力,从而达到改善氮利用效率的效果。
植物异黄酮生物合成途径中的关键酶包括异黄酮合酶(isoflavone synthase,IFS)、查尔酮还原酶(chalcone reductase,CHR)、查尔酮合酶(chalcone synthase,CHS)、查尔酮异构酶(chalcone isomerase,CHI)和苯丙氨酸解氨酶(phenylalanine ammonia lyase,PAL)等,这些酶能够在转录水平对异黄酮的合成进行调控[27-29]。IFS 作为催化大豆(Glycine max)异黄酮合成的第1 个酶,本研究发现,在氮素水平为21 和420 mg·L-1时,紫花苜蓿会通过上调茎叶和根系中的IFS基因来加快异黄酮的生物合成。在根瘤菌侵染豆科植物的过程中,结瘤因子引起宿主植物的根毛卷曲,根瘤菌从根表皮弯曲部分侵入根毛细胞,形成胞内侵染线并逐渐伸长进入皮层细胞[30],结瘤因子信号传导途径,如Ca2+信号传导途径,通过结瘤信号传递通路蛋白(nod)和乙烯应答结合因子(ERFs)、起始结瘤蛋白(nodulation initiation protein,NIN)等转录调控因子激活某些早期结瘤基因的表达以促进侵染的起始[31]。有报道指出结瘤基因的表达受异黄酮类物质的诱导[32],也有研究人员利用RNA 干扰技术抑制植物异黄酮合成路径中IFS等几个重要酶基因的表达,结果导致了豆科植物不能结瘤[33]。本研究中,相关性分析表明紫花苜蓿IFS基因与nod基因的表达具有显著正相关性,其中,茎叶中nod-1基因与IFS基因表达的相关性更强,根系中IFS-2和IFS-3基因与nod基因表达的相关性更强。可见,紫花苜蓿能通过茎叶和根系中不同IFS基因的表达进而刺激nod基因表达来促进结瘤固氮。
综上,紫花苜蓿的结瘤固氮与异黄酮含量显著正相关,在外源氮素浓度改变时,紫花苜蓿能通过茎叶和根系中不同IFS基因的表达提高体内异黄酮含量,从而刺激nod基因表达来促进结瘤和提高单株固氮潜力。
3.2 紫花苜蓿体内异黄酮与其氮效率
氮高效型作物能够在低氮肥投入的情况下达到高产优质的效果,并且有利于环境保护以及农业的可持续发展[34-35]。研究表明,根瘤菌-豆科植物共生系统的固氮量约占生物固氮总量的65%[25]。李凯等[36]通过对大豆的研究也发现,接种根瘤菌后可以显著增加大豆产量和籽粒含氮量。因此,紫花苜蓿的结瘤固氮特性对其氮效率的高低影响重大。本研究中,同一氮素水平下“LW6010”紫花苜蓿的单株总根瘤数、总根瘤重、固氮潜力以及氮积累量均大于陇东苜蓿,这也是“LW6010”紫花苜蓿氮效率较高的直接原因。本研究还发现,不同氮效率紫花苜蓿的结瘤固氮受异黄酮影响的机制不同。氮高效的“LW6010”紫花苜蓿在氮素水平为21 和420 mg·L-1时,茎叶和根系中的木犀草素、染料木素、大豆苷元、刺芒柄花素以及总异黄酮含量均上升,促进了总根瘤数和总根瘤重,从而提高了单株固氮潜力;氮低效的陇东苜蓿结瘤固氮主要受茎叶中总异黄酮含量和根系中木犀草素、大豆苷元、刺芒柄花素以及总含量的影响,在氮素水平为21 mg·L-1时,紫花苜蓿的异黄酮含量明显升高,从而提高了其总根瘤数、总根瘤重以及单株固氮潜力。宋冰等[37]对大豆的研究也表明品种间总异黄酮和各组分的含量具有显著差异。可见,紫花苜蓿体内各异黄酮含量的差异影响了其结瘤固氮能力的高低,是导致其氮效率在品种间表现差异的重要原因。
豆科结瘤和生物固氮过程均由复杂的基因系统所控制[38]。内源性异黄酮不仅可以调节根瘤菌结瘤因子形成和结瘤基因的转录表达[39],而且可以改变根瘤菌产生的胞外多糖的组成和分子量分布,在形成结瘤和调节结瘤比例中起关键作用[40-41]。本研究发现,不同氮效率紫花苜蓿的结瘤固氮受IFS基因间接调控的方式不同。氮效率较高的“LW6010”紫花苜蓿在氮素水平低于210 mg·L-1时可上调茎叶IFS基因以及根系IFS-2和IFS-3基因,在氮素水平高于210 mg·L-1时可上调茎叶和根系中IFS-2和IFS-3基因,从而提高异黄酮含量来促进结瘤固氮;氮效率较低的陇东苜蓿在氮素水平低于210 mg·L-1时,通过上调茎叶和根系中IFS-2基因而下调茎叶IFS-1、IFS-4基因,在氮素水平高于210 mg·L-1时,通过上调茎叶和根系IFS-2、IFS-3基因而下调茎叶IFS-1、IFS-4基因,从而改变异黄酮含量,进而调控结瘤固氮。有研究表明,异黄酮类物质对豆科植物结瘤基因的表达调控与外源氮素水平相关[42]。本研究发现,无论氮素水平低于或高于210 mg·L-1,氮高效的“LW6010”紫花苜蓿其IFS和nod基因比氮低效的陇东苜蓿上调幅度更大。可见,氮高效紫花苜蓿在氮胁迫时可通过IFS基因表达的协同上调提高异黄酮含量促进结瘤固氮从而改善氮效率,氮低效紫花苜蓿在氮胁迫时只有部分IFS基因上调,改变异黄酮含量来调控结瘤固氮,从而对氮效率进行调控。
由此可见,紫花苜蓿体内异黄酮含量的差异影响了其结瘤固氮能力的高低,这是导致其氮效率在品种间表现差异的重要原因。氮高效紫花苜蓿在氮胁迫时通过多个IFS基因的高表达促进异黄酮合成,进而刺激结瘤固氮,从而提高氮效率;氮低效型紫花苜蓿在氮胁迫时只有部分IFS基因上调,因此其氮效率较低。
4 结论
1)紫花苜蓿的结瘤固氮与异黄酮含量显著正相关,在外源氮素浓度改变时,紫花苜蓿能通过茎叶和根系中不同IFS基因的表达提高体内异黄酮含量,从而刺激nod基因表达来促进结瘤和提高单株固氮潜力。
2)紫花苜蓿体内异黄酮含量的差异是决定其氮效率高低的重要原因,氮高效紫花苜蓿在氮胁迫时通过多个IFS基因的高表达促进异黄酮合成,进而刺激结瘤固氮,从而表现出氮高效。
