70 份紫云英种质资源表型多样性及其在豫南地区的结实特征
2022-03-26张梦史鹏飞李本银刘春增郑春风张成兰郭晓彦张丽霞吕玉虎何春梅曹卫东
张梦,史鹏飞,李本银,刘春增*,郑春风,张成兰,郭晓彦,张丽霞,吕玉虎,何春梅,曹卫东
(1. 河南省农业科学院植物营养与资源环境研究所,河南 郑州 450002;2. 信阳市农业科学院,河南 信阳 464000;3. 中国农业科学院农业资源与农业区划研究所,北京 100081;4. 河南省作物分子育种研究院,河南 郑州 450002;5. 福建省农业科学院土壤肥料研究所,福建 福州 350013)
紫云英(Astragalus sinicus)是我国南方稻田重要的冬季绿肥,具有培肥地力以及降低化肥依赖、提升稻米产量和品质的作用[1]。随着我国农业供给侧结构性改革和绿色发展理念的逐步深入,紫云英推广利用面积逐年增加,种子需求逐年增大[2]。进一步提高紫云英种子产量和质量,保障市场稳定供给,已成为绿肥行业发展亟待解决的重要课题。目前,由于气候条件、土壤类型、耕作制度的差异,我国各地区对紫云英生长和生理特性的需求有所不同,使紫云英品种选育和扩繁生产具有明显的地域性特征[3-5]。然而,对于江南和华南地区,梅雨季与紫云英开花结实同期,低温寡日照严重影响蜜蜂授粉、小花败育率增加、结实率降低,成为紫云英种子产量形成的限制因素[2,6-7],种子成本的年际波动限制了紫云英绿肥的推广应用。因此,若能在全国范围内选择自然气候和社会经济条件适宜的区域,实现紫云英不同品种集中高产高效繁育,将极大推动绿肥生产应用的产业化发展。
豫南地区位于我国南北气候过渡带,以单季稻(Oryza sativa)-紫云英/油菜(Brassica napus)轮作为主要种植制度。由于自然气候条件适宜且存在大面积的冬闲稻田,该区曾于20 世纪90 年代被认定为国家级紫云英繁育基地。据报道,江西紫云英种子产量普遍为150~540 kg·hm-2[8-9],福建为270~345 kg·hm-2[5],湖南为330~750 kg·hm-2[5,10],湖北为600~750 kg·hm-2[11],安徽为540~705 kg·hm-2[12-13],而豫南地区紫云英种子产量可达675~1200 kg·hm-2[14-15],具有紫云英繁育的比较优势。但是,由于各地种植品种不同,分枝结实特性存在差异[4-5],上述比较优势不能排除紫云英的品种差异。同时,豫南地区在我国水稻-紫云英种植区的最北端,南方紫云英品种的越冬性、光周期限制以及收获期与当地水稻生产茬口衔接等,亦成为豫南地区全面发展紫云英种子产业的不确定因素[7]。目前,紫云英相关研究主要集中于其养分特征和生态效益[1,16-17],而对紫云英功能性状与其种子产量关系的研究较少,缺乏指导紫云英种子扩繁的功能性状相关理论基础,限制了对紫云英不同种质资源扩繁潜力的综合评估。因此,本研究通过对比54 份外地紫云英种质资源和16 份当地品种(品系)在豫南地区的生长特征和结实性状,分析紫云英不同种质资源生长、结实性状相关性,划分特征聚类并明确不同紫云英在豫南地区的繁种潜力,以期为紫云英种子产业化发展提供理论支撑。
1 材料与方法
1.1 供试资源
参试紫云英种质资源共70 份,其中53 份(编号1~53)由国家绿肥作物种质资源库提供,1 份(编号54)由福建省农业科学院土壤肥料研究所提供,15 份(编号55~69)为近5 年在信阳各地收集获得的地方紫云英种质资源,1份(编号70)为豫南地区主推紫云英品种。研究中将编号1~54 种质作为引入资源,编号55~70 作为当地资源(表1)。
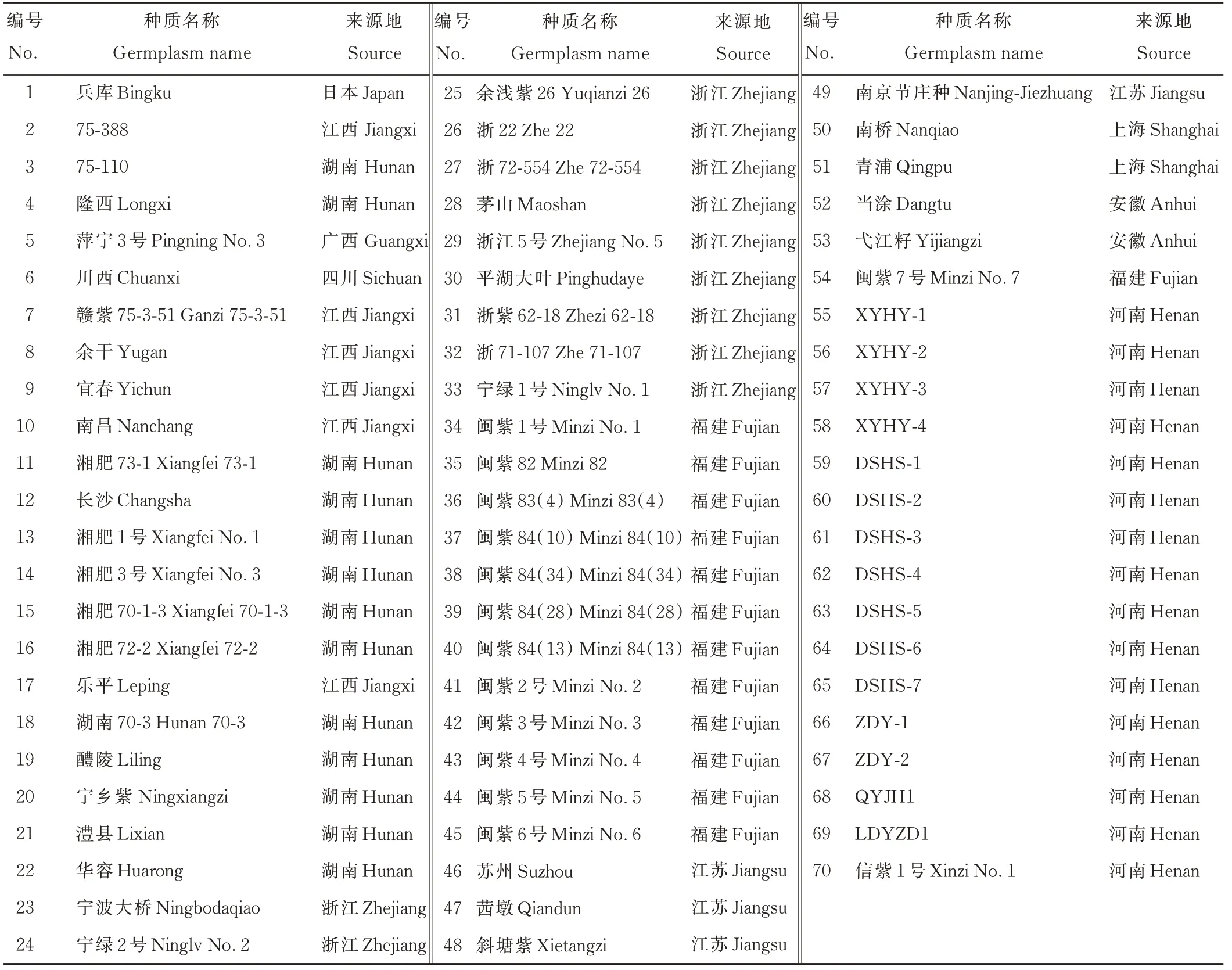
表1 70 份紫云英种质资源Table 1 Materials of 70 A.sinicus germplasm lines
1.2 试验地概况
2018 年9 月-2019 年6 月,试验在河南省信阳市罗山县周党镇桂店村进行(114°31′6.50″E,31°52′34.51″N),供试土壤为水稻土,pH 值为6.7,有机质为25.8 g·kg-1,碱解氮为84.3 mg·kg-1,有效磷16.8 mg·kg-1,速效钾79.1 mg·kg-1。该地区属亚热带向暖温带过渡区,日照充足,年均日照1900~2000 h,年平均气温15 ℃,降水量1060 mm。试验前茬作物为水稻。试验期间,无极端气象灾害发生,但12 月至次年2 月日照时长低于常年同期,5月降水量和2 月积温偏低(图1)。
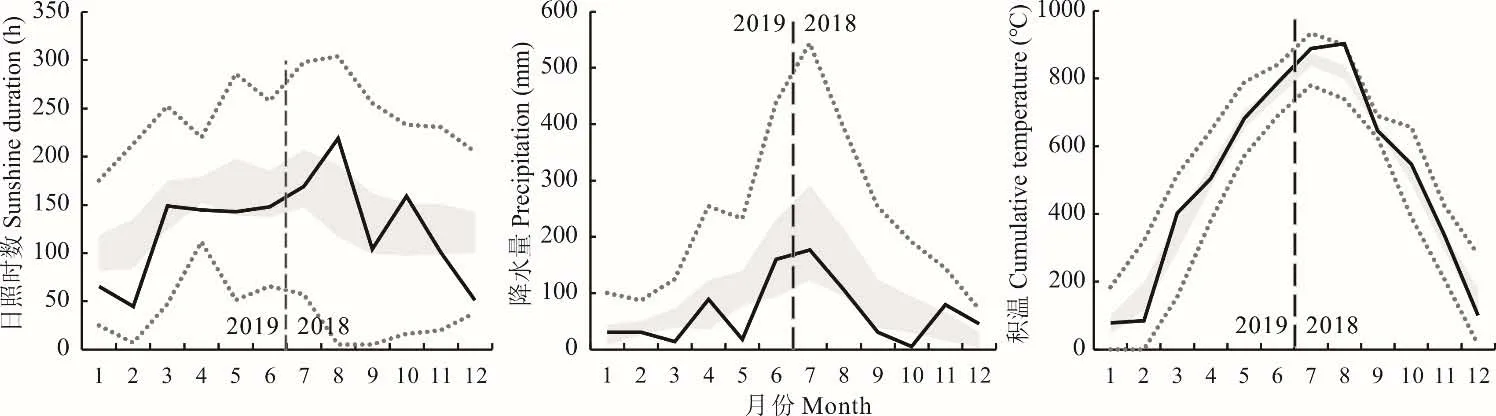
图1 试验区气候条件Fig.1 Weather condition of the experimental site
1.3 试验设计
于2018 年9 月28 日采用点播方式统一人工播种,株距10 cm×10 cm,每个资源播种于1.5 m×1.5 m 的小区中,重复3 次。播种前7 d,配合整地开沟,基施过磷酸钙225 kg·hm-2。由于供试紫云英均为原种,种质资源珍贵,为达观测品种性状的同时扩繁种子目的,在现蕾初期以孔径0.25 mm 的白色尼龙网进行资源隔离,防止窜粉影响种子纯度,盛花期进行人工授粉。
1.4 测定指标及方法
根据《绿肥种质资源描述规范和数据标准》[18],分别记录供试资源出苗期、返青期、盛花期和成熟期时间。其中,出苗期为小区内50%以上种子扎根、2 片子叶展开的日期,返青期为小区内75%以上植株产生新枝新叶的日期,盛花期为小区内75%以上的茎枝开花的日期,成熟期为小区内75%以上的荚果转为黑色的日期。
每小区随机选取5 株,测定株高(返青期和盛花期)、一级分枝数和二级分枝数(盛花期)、冠层面积(苗期、越冬期和返青期),记录叶片和主茎颜色(现蕾期)。其中,分别于2018 年11 月30 日(苗期)、2019 年2 月28 日(越冬期)和2019 年3 月27 日(返青期),通过数码相机对植株进行冠层垂直拍照(图2a),图像经SpyderCheckr 软件校色,采用Lightroom 软件校正镜头畸变,采用Photoshop 软件剔除杂质,采用Imagej 软件计算面积获得冠层面积;叶片和主茎颜色则为质量性状,即根据1-全部为绿色、2-绿色带有少许红色、3-绿色与红色相当、4-红色带少许绿色、5-全部为红色,分别定性描述并赋值记录。
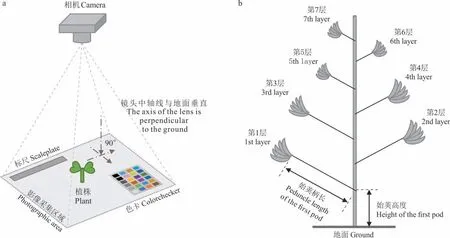
图2 图像信息采集(a)和结实特征调查方法(b)Fig.2 Image information acquisition(a)and podding characteristics survey method(b)
每小区随机选取5 株,于盛花期调查生殖分枝数和总分枝数,于成熟期调查始荚高度、始荚柄长、结荚层数(图2b)。小区内植株成熟收获后进行脱粒,测定种子产量和千粒重。由于资源较珍贵,仅在成熟期采集植株样品,60 ℃烘干后粉碎,测定全氮、全磷、全钾含量[19]。
1.5 数据处理
采用Microsoft Excel 2013 和SPSS 18 软件进行数据录入和统计分析,采用Python 的seaborn 包进行相关分析和分层聚类,采用matplotlib 包进行数据可视化。检验方差齐性、样本独立性和正态性后,对引入资源与当地资源进行单因素方差分析(One-way ANOVA)。为消除量纲对聚类关系的影响,数据通过z值标准化后进行分层聚类(hierarchical clustering)。相关分析(pearson’s correlation)以双尾(two-tailed)检验显著性。
z值标准化计算公式如下:
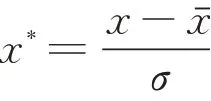
式中:x*为标准化数值;x为原数值;为样本均值;σ 为样本标准差。
根据生殖分枝数(reproductive branch number,RB)和总分枝数(total branch number,TB),计算生殖分枝比率(reproductive branch ratio,RR):
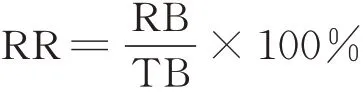
2 结果与分析
2.1 物候期特征
供试紫云英生育期存在差异(表2)。统一播种后,当地紫云英于10 月15 日-10 月18 日进入出苗期,而引入资源则在10 月13 日-10 月20 日进入出苗期,说明不同紫云英种子萌发对气候响应存在差异。越冬后,供试紫云英分别于3 月下旬进入返青期,4 月上中旬进入盛花期,5 月中下旬成熟,但当地紫云英生育期总体早于引入紫云英。出苗期至盛花期,当地紫云英平均为173 d,显著低于引入紫云英(176 d);出苗期至成熟期,当地紫云英平均为217 d,亦显著低于引入资源(220 d),说明豫南地区当地紫云英主要为早熟资源。值得注意的是,虽然引入资源生育期总体上长于当地紫云英资源,但两个群体中均存在特早熟或特晚熟的特异性种质资源,最早熟资源与最晚熟资源相差10~11 d。
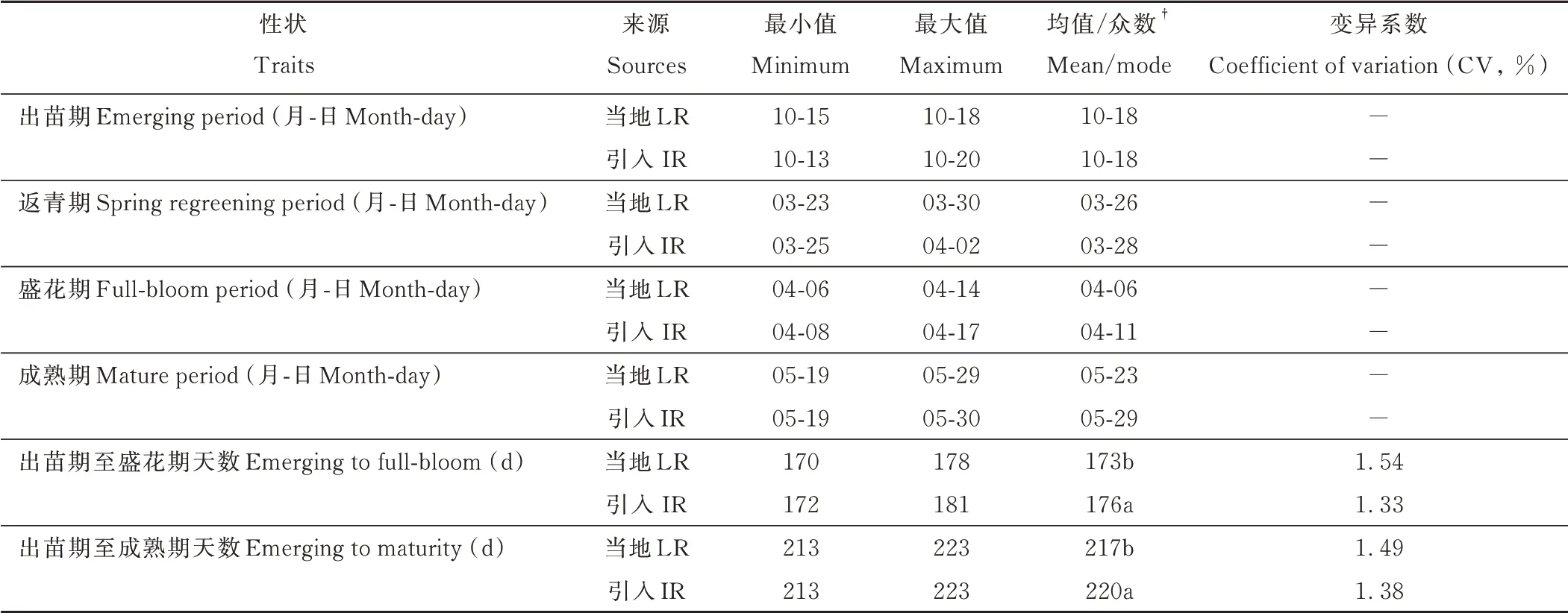
表2 不同紫云英的生育期情况Table 2 Fertility period of different A.sinicus germplasm resources
2.2 农艺性状及养分含量特征
当地紫云英和引入紫云英在返青期株高、盛花期株高以及苗期、越冬期、返青期冠层面积等性状间无显著差异,但引入资源性状的变异系数高于当地资源(表3)。在分枝数方面,引入紫云英资源的一级分枝数、二级分枝数显著高于当地资源。引入紫云英资源与当地资源茎秆颜色均以红色或红色带少许绿色为主,但当地紫云英存在全红色叶片的种质资源。值得注意的是,苗期、越冬期、返青期冠层面积的变异系数为45.9%~73.2%,说明在紫云英资源间越冬前后冠层生长差异极大。图像采集当天,南桥、弋江籽、信紫1 号以及当地种质资源XYHY-1、XYHY-2、XYHY-3、XYHY-4 和DSHS-1 叶片呈紫红色,有别于其他资源,但至现蕾期时南桥和弋江籽叶片转为绿色或绿色带少许红色(图3)。在养分含量方面,引入紫云英植株全氮和全钾含量显著高于当地紫云英,全磷含量无显著差异,但引入紫云英和当地紫云英全磷含量的变异系数分别为26.5%和26.3%,变异性较大。

图3 紫云英种质资源冠层图像Fig.3 The canopy images of the A. sinicus germplasm lines
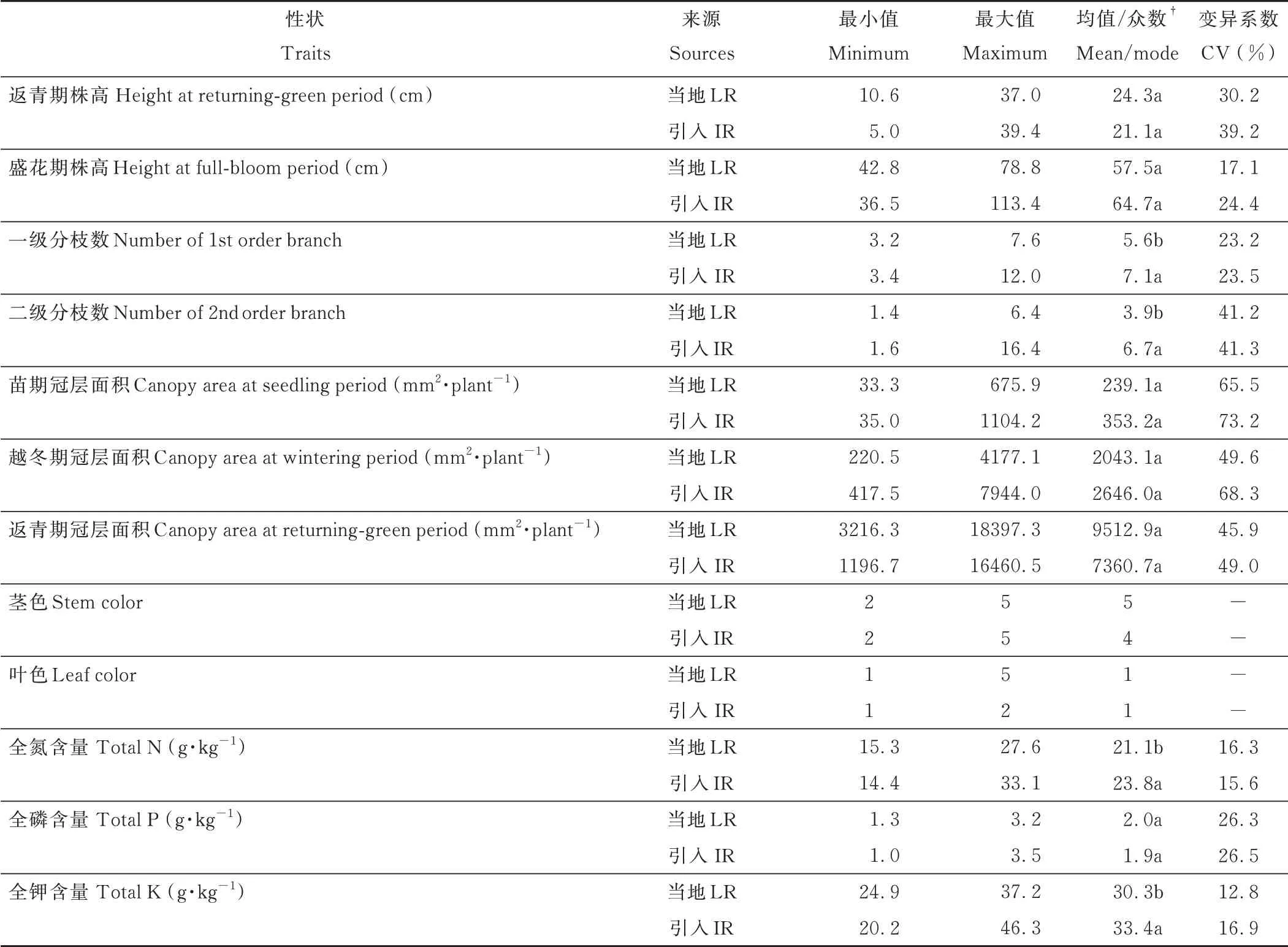
表3 不同紫云英的农艺性状及养分含量Table 3 Main agronomic characters and nutrient content of A.sinicus germplasm resources
2.3 结实特征
引入紫云英和当地紫云英的结实特征无显著差异,但性状变异性较高(表4)。当地紫云英始荚高度和始荚柄长分别为5.0~22.0 cm 和12.6~18.0 cm,引入紫云英则为4.7~41.2 cm 和10.4~19.2 cm,引入资源的变异系数高于当地资源。结荚层数方面,当地和引入紫云英最小值分别为6.8 和5.3,最高值为13.2 和14.0,不同资源结荚层数相差2~3 倍。当地和引入紫云英的生殖分枝数分别为0.8~4.0 和0.4~5.6,生殖分枝比率分别为19.2%~87.5%和11.8%~100.0%,种子千粒重分别为1.9~3.6 g 和2.6~3.7 g,不同紫云英资源的结实特征变异性较高。成熟期当地紫云英种子产量为562.5~1644.0 kg·hm-2,而引入紫云英为346.5~1941.0 kg·hm-2,显示出不同紫云英种质资源在豫南地区繁种能力的差异。在植株个体结实性状中,始荚高度、生殖分枝数、生殖分枝比率的变异系数较高,可能是造成最终种子产量差异的重要因素。
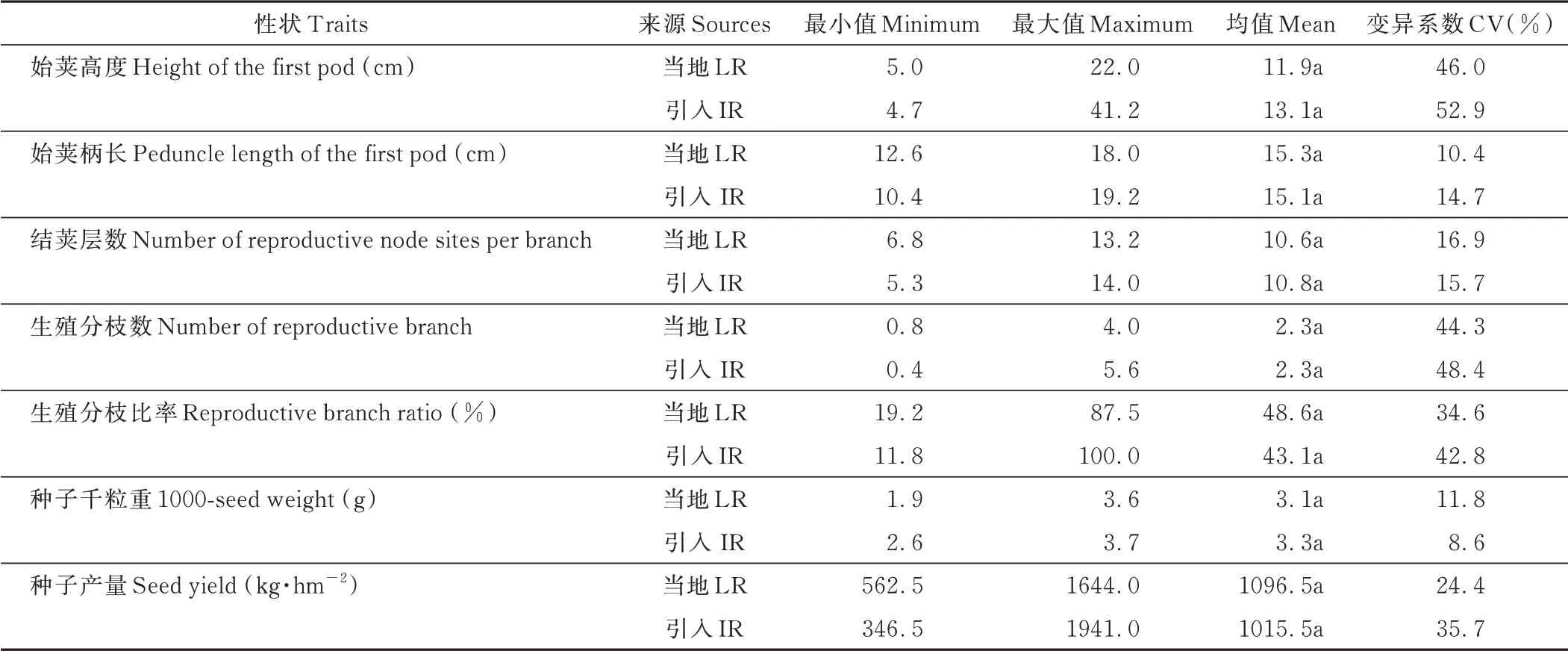
表4 不同紫云英的结实特征Table 4 Podding characters of different A.sinicus germplasm resources
2.4 功能性状与结实特征的相关性
生育期天数、农艺性状、养分含量以及结实性状之间存在相关性。如图4 所示,种子产量与返青期株高、盛花期株高、越冬期冠层面积以及始荚高度、结荚层数、种子千粒重显著正相关;种子千粒重则与返青期株高、盛花期株高、越冬期冠层面积以及始荚高度、结荚层数、生殖分枝比率显著正相关,与出苗至盛花期天数、全氮和全磷含量显著负相关。对于生殖分枝数、生殖分枝比率和结荚层数这3 个紫云英种子产量的构成因素,生育期天数、植株全氮和全磷含量、一级和二级分枝数均与其呈负相关,而返青期株高、盛花期株高、越冬期冠层面积、返青期冠层面积均与其呈正相关。始荚高度和始荚柄长反映了紫云英初始结荚的空间信息,不仅与返青期、盛花期株高以及苗期、越冬期、返青期冠层面积显著正相关,同时与植株全钾含量显著正相关。叶片颜色与出苗至盛花期、出苗至成熟期的天数以及一级和二级分枝数显著负相关,说明叶色越红,则生育期越短、分枝数越少。总体观之,具备生育期短、返青期和盛花期株高较高、越冬期和返青期冠层面积较大特征的紫云英,其种子产量和种子千粒重较高。
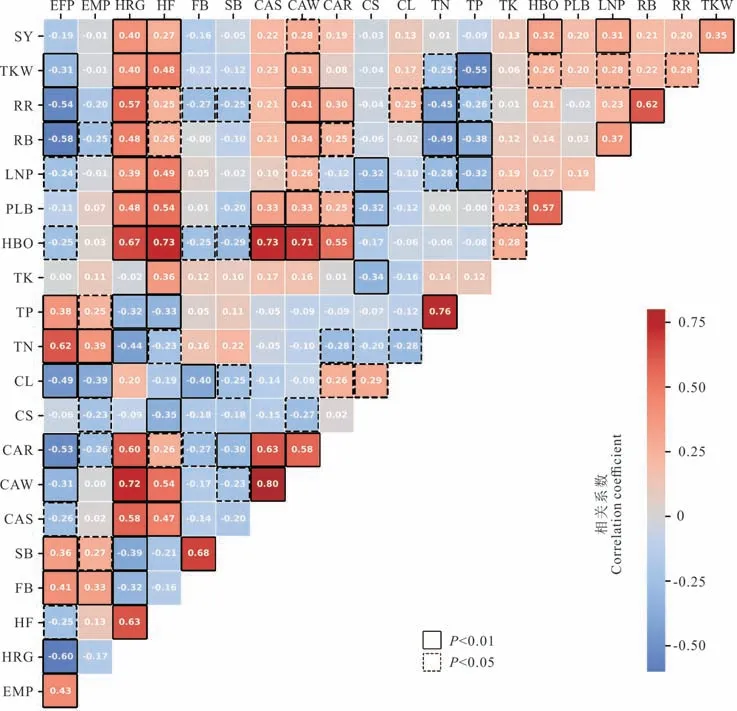
图4 紫云英主要性状间的相关性Fig.4 The relationships between traits of the A. sinicus germplasm lines
2.5 不同紫云英种质资源的聚类关系
21 个性状指标可分为4 个性状聚类(图5),其中一级和二级分枝数、全氮和全磷含量、出苗至盛花期天数以及出苗至成熟期天数为一类,反映紫云英作为绿肥的综合利用价值;苗期、越冬期、返青期冠层大小以及返青期、盛花期株高、始荚高度和始荚柄长为一类,反映紫云英的越冬能力及其结实空间特征;茎叶颜色为一类,反映紫云英的感官特征;结实分枝数、结实分枝比率、全钾含量、结荚层数、种子千粒重和产量为一类,反映紫云英的结实能力。基于此,70 个紫云英资源分为10 个种质聚类。其中,聚类Ⅰ结实能力较弱,聚类Ⅱ氮磷含量较高、生育期较长、结实能力较弱,聚类Ⅲ生育期较长,聚类Ⅳ分枝性能较高,聚类Ⅴ养分含量和种子产量均较高,聚类Ⅵ叶片红色、生育期短,聚类Ⅶ越冬前后冠幅大、结实空间较大、种子产量高,聚类Ⅷ氮磷养分含量低、生育期短、生殖分枝数和比率较高,聚类Ⅸ越冬后冠幅较大、结实空间较大、结实能力较强,聚类Ⅹ结实空间较大。比较引入紫云英和当地紫云英资源在不同聚类的分布发现,现蕾期叶片全为红色的聚类Ⅵ均为当地紫云英资源,聚类Ⅳ、Ⅴ、Ⅸ仅包含引入紫云英资源。值得注意的是,结实能力较弱的聚类Ⅰ和Ⅱ为引入资源和当地资源的混合聚类,说明本地和引入紫云英均存在繁种能力差的种质资源;聚类Ⅳ、Ⅵ、Ⅶ和Ⅷ中具有特异性状(分枝能力、叶色、始荚高度、生殖分枝数)的资源,显示出紫云英种质资源的遗传多样性。
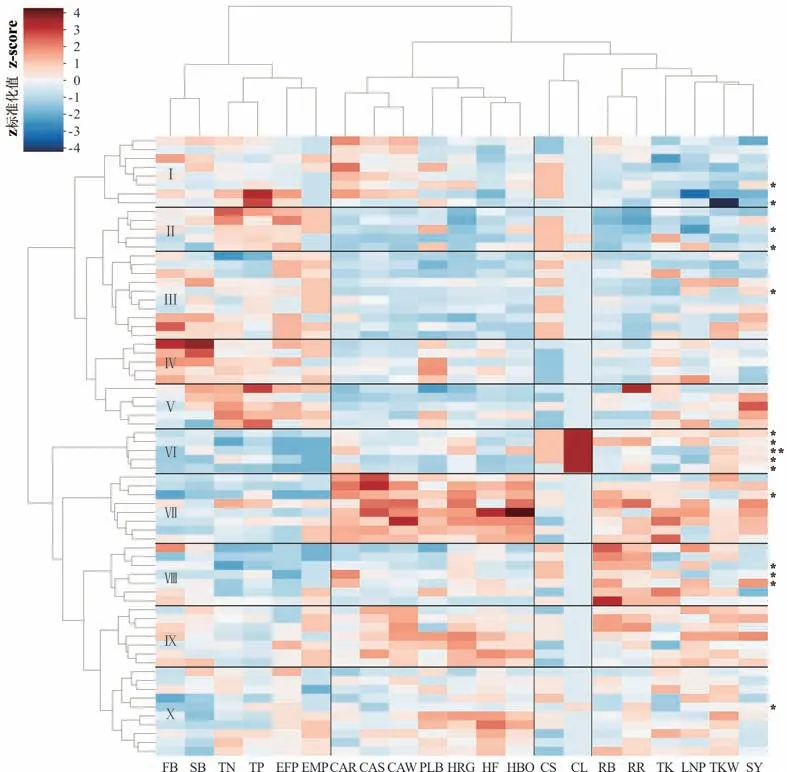
图5 紫云英种质资源及性状特征聚类关系Fig.5 Clustering relations between different traits and germplasm lines of A. sinicus
3 讨论
3.1 紫云英种质资源的遗传多样性
作物的表型性状变异是生物遗传多样性在形态水平上的表现,反映了自身基因与外界生活环境互相作用的结果。何春梅等[5]对10 个紫云英地方品种研究发现,盛花期不同紫云英单株分枝数为2.6~4.8、株高为40.0~81.1 cm、叶面积为1.03~2.68 cm2,变异系数分别为21.1%、21.9%和32.7%。本研究中不同紫云英资源各性状的变异系数为1.3%~73.2%,其中苗期、越冬期、返青期冠层面积的变异系数较高,为45.9%~73.2%,始荚高度为46.0%~52.9%、生殖分枝数和生殖分枝比率为34.6%~48.4%、二级分枝数为41.2%~41.3%、返青期株高为30.2%~39.2%、种子产量为24.4%~35.7%,是调查21 个数量性状中变异性较高的9 个性状(表3~4)。同时,供试紫云英中存在特异性资源。南桥、弋江籽、信紫1 号以及当地种质资源XYHY-1、XYHY-2、XYHY-3、XYHY-4 和DSHS-1 在返青期叶片呈现紫红色,显著区别于其他资源(图3),南桥和弋江籽至现蕾期时叶片转为绿色或绿色带少许红色(图3),而当地紫云英资源叶片红色一直保持至成熟期。研究发现,花青素苷在植物抵抗干旱、低温、盐胁迫、低氮等非生物胁迫中具有重要的作用[20]。张彬等[21]对谷子(Setaria italica)花青素累积与低温胁迫响应研究中发现,低温可诱导紫叶品种叶肉细胞和表皮细胞中花青素累积,并与脯氨酸和可溶性糖参与植物低温胁迫响应。Zhu 等[22]对冬季肖蒲桃(Acmena acuminatissima)的幼嫩叶片研究发现,花青素在幼嫩叶片的累积可以作为光衰减器,降低低温条件下光照对光合系统的损伤。本研究中返青期和现蕾期呈现完全紫红色叶片的紫云英均为豫南地区当地资源(表3 和图2),推测在长期适应性进化过程中,当地紫云英资源保留或获得了通过大量累积花青素抵御北方低温胁迫的抗逆生理机制。然而,目前对于紫云英应对非生物胁迫的基因与环境互作的分子机制尚无报道,有待深入研究。
3.2 紫云英功能性状对结实能力的影响
植物功能性状间存在普遍的权衡关系,这种关系是经过自然筛选和进化形成的性状组合[23]。Westoby[24]曾提出“叶片-株高-种子(leaf-height-seed,LHS)”的性状权衡维度理论,用以评价植物的生存策略。紫云英虽然是我国传统的绿肥作物,但其生长和繁殖仍具有明显的未驯化或待驯化特征[25],功能性状间仍保留了较普遍的关联(图4)。本研究中种子产量和种子千粒重与返青期株高、盛花期株高、越冬期冠层面积显著正相关,符合LHS 性状权衡理论。秦自果等[11]研究发现,紫云英群体密度增加,株高、茎基部直径、单株重、单株一级分枝数和一级分枝花序数显著降低,说明紫云英功能性状对群体密度具有关联响应。本研究在种群密度固定条件下进行,生殖分枝数、生殖分枝比率和结荚层数与生育期天数、植株全氮和全磷含量、一级和二级分枝数呈负相关,与返青期株高、盛花期株高、越冬期冠层面积、返青期冠层面积呈正相关,说明紫云英营养/生殖性状存在一定组合联动关系,其中返青期株高、越冬期和返青期冠层面积、植株全磷含量对紫云英结实具有较大影响,应是紫云英种子繁育中人工调控的重要目标性状和指标参数。此外,苗期、越冬期和返青期冠层面积三者间存在极显著正相关性,说明苗期冠层生长量与紫云英越冬和返青冠层面积关联,印证了前人关于“促进苗期生长可提高紫云英越冬能力”的实践经验[7],同时也间接说明苗期紫云英生长状况间接影响紫云英结实和种子产量(图4),重视苗期管理对豫南地区紫云英种子繁育具有重要意义。
3.3 豫南地区紫云英种子产业发展的潜在价值
豫南地区为单季稻区,冬季普遍撂荒,部分区域种植油菜、小麦(Triticum aestivum)等冬季作物。对于撂荒地,除去播种用种和管理成本,即为紫云英繁种收益;对于油菜和小麦种植田地,若进行紫云英繁种则当年不能进行小麦和油菜规模生产。目前,豫南地区小麦平均产量为5100 kg·hm-2,产值10200 元·hm-2;油菜产量为1875 kg·hm-2,产值6750 元·hm-2[26]。近5 年来,全国紫云英种子平均收购价为14 元·kg-1。基于上述信息,若豫南地区紫云英种子产量达525 kg·hm-2,其收益高于油菜;产量达750 kg·hm-2,则收益高于小麦。本研究中,虽然越冬期日照和2 月积温均低于常年同期水平,但70 个紫云英种质资源中,66 个资源的种子产量高于525 kg·hm-2,54个资源种子产量高于750 kg·hm-2,分别占总资源数的94.3%和77.1%,说明绝大多数紫云英在豫南地区繁种具有较高的经济效益。何春梅等[5]在福建对10 个紫云英地方品种进行比较试验,种子最高产量为345 kg·hm-2;鲁艳红等[4]在湖南双季稻区对5 个紫云英品种进行比较试验,种子产量为336~738 kg·hm-2。上述研究的供试品种中弋江籽、湘肥3 号均在本研究中涉及,产量均高于引入资源产量平均值(1015.5 kg·hm-2,表4 和图5),显示出豫南地区发展紫云英种子产业的区域优势。此外,在越冬方面,本研究中供试紫云英未出现越冬期前后死苗现象。在以苗期、越冬期、返青期冠层大小表征不同紫云英的越冬能力上,聚类Ⅰ、Ⅶ、Ⅸ紫云英越冬前后冠层较大,聚类Ⅲ、Ⅵ、Ⅷ则在越冬过程中冠层大小排序逐渐升高(图5,颜色由蓝转红,说明在70 个资源中排序位置上升),说明这6 个聚类的紫云英越冬性较强。聚类Ⅴ的冠层大小排序逐渐降低(图5,颜色由红转蓝),虽然其种子产量仍然较高,但其越冬性不及其他资源。在茬口衔接方面,本研究中由于9 月积温和降水量较低(图1),紫云英出苗期整体延后5~7 d,在5 月中下旬成熟(表2),较之于常年成熟期推迟1 周。虽然紫云英成熟期可与该区水稻移栽期衔接,但错过水稻播种期[27],说明紫云英种子繁育在该区移栽稻田具有可行性,但在直播稻生产上存在茬口衔接的限制。针对这一问题,是否可以通过稻底套播方式实现生育期前移,以满足直播稻生产需要,尚不清楚。同时,紫云英开花结实与光周期变化的信号机制还未明确,但有研究显示,早播紫云英生物产量高,迟播紫云英种子产量高[28]。因此,如何协同紫云英品种、生育期和种子产量,解决超晚熟紫云英品种繁育与当地水稻生产茬口衔接问题,仍需进一步研究。
4 结论
19 个调查性状的变异系数(CV)为1.3%~73.2%,其中苗期、越冬期、返青期冠层面积(CV=45.9%~73.2%)、始荚高度(CV=46.0%~52.9%)、生殖分枝数和生殖分枝比率(CV=34.6%~48.4%)、二级分枝数(CV=41.2%~41.3%)、返青期株高(CV=30.2%~39.2%)、种子产量(CV=24.4%~35.7%)变异性较高。信紫1 号、XYHY-1、XYHY-2、XYHY-3、XYHY-4 和DSHS-1 这6 个当地资源在越冬期和返青期叶片呈现紫红色,明显区别于其他资源。返青期株高、越冬期和返青期冠层面积、植株全磷含量对紫云英结实具有较大影响,是紫云英种子繁育中人工调控的重要目标性状和指标参数。
70 份紫云英种质资源在豫南地区可以完成扩繁,可分为10 个聚类,反映绿肥综合利用能力、越冬能力及其结实空间特征、感官特征、结实能力的差异。引入资源平均种子产量为1015.5 kg·hm-2,当地资源平均种子产量为1096.5 kg·hm-2。94.3%的供试紫云英种子产量高于525.0 kg·hm-2,77.1%的供试紫云英种子产量高于750.0 kg·hm-2,紫云英在豫南地区繁种具有较高的经济效益,适合移栽稻接茬。
