Epichloë 内生真菌对禾草种子萌发影响研究进展
2022-03-26李春杰郎鸣晓陈振江陈泰祥刘静金媛媛魏学凯
李春杰,郎鸣晓,陈振江,陈泰祥,刘静,金媛媛,魏学凯
(1. 兰州大学草地农业生态系统国家重点实验室,兰州大学农业农村部草牧业创新重点实验室,兰州大学草地农业教育部工程研究中心,甘肃省西部草业技术创新中心,兰州大学草地微生物研究中心,兰州大学草地农业科技学院,甘肃 兰州 730020;2. 中国林业科学研究院国家林业和草原局草原研究中心,北京 100091)
禾草内生真菌(grass endophyte)是指能在宿主禾草体内完成其全部或部分生命周期而宿主本身不表现外部病症的一类真菌[1]。本研究中出现的内生真菌仅指子囊菌门(Ascomycota)麦角菌科(Clavicipitaceae)的Epichloë属。Epichloë属内生真菌与宿主禾草在协同进化过程中逐步形成互利共生关系[2-3],其在宿主抗旱[4-5]、耐盐[6]、耐寒[7-8]、耐重金属[9]等非生物胁迫方面及抗虫[10]、抗病[11]、抗线虫[12]等生物胁迫方面均发挥着积极作用。宿主禾草与Epichloë内生真菌共生可促进植物激素和次生代谢物生成,影响其自身生长发育[13],并减少家畜采食[14]。此外,禾草内生真菌能够提高宿主在植物群落中的竞争能力[15],扩大其分布范围。正是禾草内生真菌在自然系统中产生的积极影响,才引起国内外学者的广泛研究(图1)。

图1 国内外禾草内生真菌研究现状(1977-2021 年)Fig. 1 Current research status of grass endophyte at China and abroad(1977-2021)
种子萌发(seed germination)是植物生长发育的第一阶段,是有活力的种子从吸水进行呼吸、物质合成等生命代谢活动,到种胚突破种皮露出胚根的一个阶段[16]。种子萌发能力与种子自身内部生理状态及外部环境因素有关,只有在其自身具有生命活力且环境条件适宜时才能正常萌发[17]。在这一时期,种子对外界环境适应能力最差[18],其对胁迫的响应尤为明显。种子活力与萌发成苗关乎植物进入自然和农业生态系统的时间,直接影响农作物产量,对经济和生态发展具有重要作用[19]。从质量来看,种子萌发质量不仅对其自身播种品质和环境适应能力等方面有至关重要的作用,在植物整个生长发育过程[20]、物种繁殖、种群维持、植被分布和动态变化方面也具有重要意义。
Epichloë内生真菌在禾草体内的传播方式有无性型垂直传播和有性型水平传播[21-22],且以无性型垂直传播为主。在共生体形成种子的过程中,真菌菌丝先侵入花序,并与禾草同步生长,菌丝逐渐伸入种子胚胎,进而侵染胚珠、心皮等部位,胚形成后,菌丝逐渐定殖于胚芽鞘原基及叶原基[14,24]。在这期间,菌丝一直存在于禾草种子胚轴及盾片中,直至种子萌发[23]。种子内菌丝密度最高,主要存在于糊粉层[24]。有研究发现,内生真菌能够促进种子萌发、提高禾草种子对其伴生植物的化感作用及宿主禾草的建植速率,进而使宿主禾草占据有利生长空间与竞争优势[25]。种子萌发需要多种植物激素共同调节,其中赤霉素(gibberellin,GA)和生长素(auxin,IAA)在种子萌发过程中起主要促进作用,已有研究发现在体外培养的高羊茅内生真菌中可以产生吲哚乙酸[26]。另外,内生真菌能够促进禾草种子在胁迫环境下萌发,其原因可能是带菌禾草在收到胁迫信号时可诱导细胞间隙的真菌做出某种响应传递给植物,产生的内源信号物质可诱导植物激素水平提高,进而抵抗外界不利因素[27-29]。
本研究将对不同生境及处理下Epichloë内生真菌在禾草种子萌发方面的影响进行扼要总结,为禾草内生真菌共生体抗性机制研究,内生真菌资源开发利用和优良草地建植、畜牧业良性发展提供理论支持。
1 正常萌发条件
内生真菌垂直传播特性在宿主生长阶段发挥作用,因此共生体种子的萌发也必将受到内生真菌影响[30]。White 等[31]指出,内生真菌通过产生生长素和具有相似功能的生物碱,或调节植物体内激素水平,进而促进植物生长。研究发现,带菌(E+)黑麦草(Lolium perenne)和高羊茅(Festuca arundinacea)种子在适宜萌发条件下其发芽率比不带菌(E-)种子高约10%,且种子饱满度更高[32]。周景乐[33]通过比较不同养分供应下E+和E-野大麦(Hordeum brevisubulatum)种子发芽指标,发现E+野大麦种子萌发情况始终优于E-种子。但也有研究指出,禾草内生真菌对黑麦草[30]和高羊茅[34]种子发芽率影响不显著,却能缩短发芽所需时间,推测其原因可能是内生真菌与宿主种子竞争营养而迫使种子提前发芽。内生真菌对禾本科种子的萌发影响虽存在差异,但总的来说其对禾草种子萌发具有积极作用。
2 胁迫
2.1 非生物胁迫
2.1.1 干旱及渗透胁迫 种子萌发需要充足的水分以活化相关酶、分解贮藏有机物、完成新陈代谢。近年来,人口增长和社会发展加剧了温室气体排放和全球气候变化进程,使得农田干旱趋势日益加重[35]。加大灌溉开发与发展力度是缓解干旱农业生产和发展所面临困境的主要措施。然而在我国,农业灌溉用水严重不足,长此以往,将会改变禾草形态结构和生理功能,导致禾草细胞内环境紊乱,蛋白质结构及功能破坏。内生真菌的侵染可增强宿主禾草耐旱性能已被多次证实,其通过增强宿主植物根系、叶片发育,调节宿主叶片气孔活动及渗透势以抵抗干旱胁迫[13]。因此,利用内生真菌提高禾草自身抗旱性或耐旱性是应对干旱胁迫的一项行之有效的手段。
PEG模拟干旱实验发现,内生真菌能显著提高鹅观草(Roegneria kamoji)[36]、醉马草(Achnatherum inebrians)[37]的发芽率,且在干旱初期,内生真菌通过促进胚芽、胚根生长及提高幼苗含水量,以抵抗外界不利因素,保证种子正常萌发[36]。另有研究表明,内生真菌与黑麦草[30,38]共生除了能够在干旱水平下提高种子的发芽率和促进胚根生长,还可以缩短种子发芽所需时间。然而在高羊茅[39]种子萌发实验中,E+种子的耐旱能力低于E-,这可能与内生真菌的侵染在种子萌发过程中产生自毒物质有关,也可能是在种子萌发初期,内生真菌的生长消耗了种子自身有限的营养物质而使E+种子处于劣势。在许多研究中发现,内生真菌往往对胚根生长有显著影响,而对胚芽生长影响较小[33,40],可能的原因是内生真菌能够调节宿主种子中营养元素分配或激素生成,通过促进植物根系发育,吸收更多水分来保证宿主种子在干旱胁迫下成活[41]。Epichloë内生真菌对不同共生体组合间影响的差异与共生体基因型、生活史及生存环境条件相关。建议通过转录组学对耐旱常用模式禾草内生真菌共生体进行深入分子机制研究,挖掘耐旱相关基因,为今后禾草内生真菌耐旱种质创新提供依据。
2.1.2 酸碱胁迫 酸碱环境对禾草种子萌发的影响是建立盐碱化禾草草地的关键问题之一[6]。土壤酸化会降低土壤缓冲体系的缓冲能力,施用石灰等碱性物质是改良土壤酸化的传统手段,但长期施用会使土壤养分失衡,且在复酸化过程中,土壤会由于碱性物质消耗而再度酸化,以至于加剧其酸化程度。土壤理化性质对植物自身理化性质有关键影响,植物体内环境pH 对其自身生长发育和细胞内多种生理生化代谢起重要作用[42],且直接影响植物吸收养分和水分的能力。不适宜环境将会导致植物对营养元素吸收不佳,进而导致植株发育不良,根系不发达[43]。有研究发现Epichloë内生真菌能缓解酸碱胁迫环境对禾草的影响[44],因此利用内生真菌提高禾草宿主的酸碱耐性成为一种理想禾草育种手段。
彭清青等[44]发现,在偏中性(pH 6~11)及酸性(pH 4~5)环境中,E+醉马草、中华羊茅(Festuca sinensis)和野大麦种子的发芽势及发芽率总体上均大于E-,且在酸碱环境下E+种子异状发芽率小于E-。对不同pH 砖红壤浸提液处理的多年生黑麦草种子进行萌发实验,发现在一定范围内随着酸胁迫的增强,E+种子各项萌发指标均高于相应的E-种子,说明内生真菌能促进多年生黑麦草在酸胁迫环境中的萌发,同时扩大种子对酸胁迫的耐受范围,增强其对酸胁迫的抵抗力[45]。关于碱胁迫,万志文[46]发现,E+醉马草幼苗的株高、根长在一定范围内均随pH 升高而增加,且醉马草内生真菌共生体对强碱的耐受性大于强酸。分析这些结果可以得出,内生真菌能够扩展宿主对酸碱环境适应性的范围,进而提高其生存的竞争优势。而对于烈性毒草醉马草而言,宽泛的酸碱适应性是其侵害北方退化草地的原因之一[44]。因此,找到一种有效防除醉马草-内生真菌共生体的方法是草地生态修复行之有效的手段。过高的pH 环境通过抑制根系发育和ATP 合成进而影响植物生长[46]。先前的研究证明内生真菌能够促进胚根发育,增强根系活力[47],内生真菌的增益作用是否体现在萌发期缓减胚根的酸碱毒害有待进一步研究。
2.1.3 盐胁迫 土壤盐渍化造成的资源和环境生态问题是全世界面临的一大挑战,据报道,全球接近20%的耕地和50%的灌溉地受盐分影响[48]。土壤中过量盐离子会抑制种子萌发[49],对植物造成离子毒害、渗透胁迫和氧化胁迫[50]。盐渍化土壤的改良主要有工程、化学、物理、生物4 种措施,其中生物措施最为环保健康,尤以耐盐植物对土壤改良最为理想科学[50]。许多研究表明[6,51],Epichloë内生真菌能提高多种禾草耐盐性,因此,利用内生真菌进行耐盐禾草的研究与育种成为许多科学家的研究重点。
种子在萌发时对盐表现敏感。有学者通过比较E+与E-种子萌发指标,发现德兰臭草(Melica transsilvanica)[51]、醉马草[52]、中华羊茅[53]和野大麦[6]E+种子在盐胁迫下的发芽率均高于E-,特别在300 和400 mmol·L-1NaCl 浓度下,E+野大麦种子发芽率显著高于E-,同时,内生真菌能促进宿主在高盐环境中胚芽和胚根的生长。另外,盐胁迫抑制了黑麦草[54]、中华羊茅[53]种子发芽率,但E+种子发芽率下降趋势更缓,且低盐环境对E+种子萌发抑制不显著[54]。内生真菌提高禾草耐盐机理研究已取得一定进展,其中光合能力、氮代谢、氮利用、叶片相对含水量、可溶性糖、脯氨酸含量、过氧化物酶活性及离子比率均参与到禾草植株耐盐活动中[51-55],但关于内生真菌对禾草种子耐盐萌发机理影响的研究较少,种子生理状态不同于植株,内生真菌对种子耐盐机理是否与其产生生物碱或刺激宿主自身合成生物碱及其他次生代谢物有关,是否通过形态和生理两方面进行调控有待进一步研究。
2.1.4 重金属胁迫 随着现代社会发展,尾气排放,农药、化肥[9]的大量使用,导致重金属污染问题日益严峻。重金属离子的排放严重破坏了土壤和水质,当土壤中重金属含量大于植物对其自净能力时,则会表现出毒害症状,这对植物的生长发育与代谢活动造成严重的影响,并通过食物链进入人体,对人类健康构成威胁[56-57]。研究发现,内生真菌能够提高宿主禾草对某些重金属的耐受性,因此利用禾草-内生真菌共生体进行土壤重金属修复是生态修复的理想手段。
Epichloë内生真菌能在适宜重金属浓度下增加宿主禾草对营养物质的吸收,进而促进植物生长[56],并在一定范围内对过量重金属如铜(Cu)、锌(Zn)、铝(Al)、铅(Pb)等造成的植物体内水平衡紊乱、细胞膜透性降低、细胞损伤,胚乳蛋白破坏起到缓解作用[57-58],进而缓减重金属胁迫对带菌禾草种子的萌发抑制作用。研究发现,E+醉马草种子在锰(Mn2+)胁迫处理下,其发芽率与发芽指数均显著大于E-种子[57]。在Pb[58]、镉(Cd)[59]胁迫下,E+德兰臭草种子的发芽率、发芽势、活力指数均显著高于E-,特别是在Pb 浓度大于800 mg·L-1时,内生真菌能够降低德兰臭草幼苗中Pb 含量。植物在低浓度重金属胁迫中,通过排斥、固定、抗氧化酶、络合等方式以达到解毒的目的,同时,重金属离子能够通过刺激金属硫蛋白和植物螯合态合成而促进胁迫蛋白的积累。然而,当重金属浓度超过一定阈值时,植物中蛋白酶和淀粉酶受到抑制而影响植物正常生理代谢[58]。有报道称,内生真菌缓解重金属离子毒害的机制可能与其产生的酚类物质与重金属螯合而使重金属活性降低有关[58],而内生真菌的侵染是否参与及如何调节植物胁迫蛋白合成、调控重金属在宿主种子中的分配;内生真菌是否通过影响种子内部生理变化拓宽宿主植物对重金属胁迫耐受范围还需进一步探究。
2.1.5 温度胁迫 温度在种子萌发阶段起关键作用,种子的正常萌发和出苗均与温度息息相关[60]。种子通过萌发时对环境温度变化的感知以检测植被冠层和土壤埋藏深度是否适合后续植物生长[61]。当前,全球气候变化及过度放牧,使得天然草地的生长受到威胁,因此,研究如何提高不同生境下禾草种子萌发适应性对草地修复及草种扩繁具有重要意义。
Epichloë内生真菌可提高野大麦在高温(30 ℃)和低温(10 ℃)胁迫下种子发芽率和发芽指数,而在极限高温(35 ℃)和低温(5 ℃)下,内生真菌并不能促进种子萌发[41],可能的解释是极限温度抑制了种子自身的萌发或内生真菌在该温度下失活甚至死亡,内生真菌的侵染能够使宿主种子有更低的基点温度和更高的上限温度。有学者发现,内生真菌能够促进中华羊茅[62]、醉马草[63]种子在不同低温胁迫下种子的萌发及胚根、胚芽生长。通过进一步探究E+、E-种子萌发中的差异表达基因,发现这些基因与生物碱合成、脂肪酸代谢等途径有关。生物碱是内生真菌提高禾草抗逆性能的重要物质,而不饱和脂肪酸含量的增加能够使膜脂相变温度降低,进而保证低温胁迫下的膜流动性,此两者均可能是内生真菌提高宿主在低温胁迫下萌发的重要机制。通过代谢组学分析,发现E+醉马草种子中逆境防御蛋白、胁迫应激蛋白和一些参与代谢途径的相关酶大量表达[63]。说明内生真菌除了能够激活宿主种子自身防御应激反应外,还能加速能量代谢与消耗,以抵消低温胁迫对种子萌发带来的负面效应。E+种子高发芽率和发芽速度能够提高禾草建植速度,使禾草占据有利的生长空间和竞争优势,这对干旱、寒冷及热带地区草地建植有积极影响。因此,禾草-内生真菌共生体耐寒耐热功能基因的挖掘、分子机理的探究与验证及生理生化机制将对未来禾草育种具有重要意义。
2.2 生物胁迫
在自然环境中,植物的萌发生长过程不可避免地受到细菌、真菌、病毒和害虫等生物侵害,提高宿主植物耐受生物胁迫的方法成为各国科学家的研究热点。内生真菌通过产生麦角生物碱(ergot alkoloids,ergonovine 和ergine 为代表)、吲哚双萜类(indolditerpene,lolitrem B 为代表)、吡咯并吡嗪类(pyrrolopyrazine,peramine 为代表)及饱和吡咯类化合物(pyrrolizidine,loline 为代表)4 大类生物碱[64-65],对禾谷缢管蚜(Rhopalosiphum padi)、朱砂叶螨(Tetranychus cinnabarinus)[4]、亚洲小车蝗(Oedaleus decorus)和针毛收获蚁(Messor aciculatus)[10]等昆虫产生毒性,或是降低禾草的适口性,进而提高宿主禾草对昆虫的抗性[66]。然而,由于昆虫主要采食禾草植株,其对禾草种子萌发阶段的负面效应尚不明确,关于带菌禾草的萌发质量对后续植物生长及产生生物碱致毒剂量等抗虫机理有待进一步研究验证。
Li 等[67]发现在多年生老芒麦(Elymus sibiricus)种子萌发阶段,Epichloë内生真菌能够提高宿主对细交链孢(Alternaria alternata)、小 孢 壳 二 孢(Ascochyta leptospora)、离 蠕 孢(Bipolaris sorokinianum)及 燕 麦 镰 孢(Fusarium avenaceum)的抗性,同时,带菌醉马草幼苗对离蠕孢、新月弯孢(Curvularia lunata)、锐顶镰孢(Fusarium acuminatum)和细交链孢有不同程度的抑制。该结论与张永雯[55]的研究结果一致,即在接种细交链孢菌、密实链格孢菌(Alteruaria compacta)、半裸镰刀菌(Fusarium incarnatum)等13 种种带病原真菌后,E+野大麦种子萌发的各项指标均大于E-种子。种带病原真菌通过释放某些代谢产物或有毒物质来降低一些酶的活性,抑制某些氨基酸的合成,进而对种子萌发造成影响。蛋白组学分析表明,内生真菌侵染的宿主中,三萜类物质蛋白表达量明显提高,三萜类物质作为植物中的一种次生代谢产物,其在植物抗病抗虫方面具有重要作用[63,68]。内生真菌与植物的共生可增强宿主对病原菌的抗性,除了保护性生物碱的作用外,也与内生真菌在植物组织上形成菌丝网而造成对病原菌的“生态位排斥”有关。另外,有研究指出内生真菌作为一种外源生物因素,在与植物共生过程中可能被识别为病原真菌而提前诱发出植物的防御反应[67,69],因此在病原真菌入侵时,带菌植物已经适应或已具备较为成熟的防御体系。这些研究结果说明内生真菌提高种子在生物胁迫下的萌发能力,不仅与其诱导植物产生相关防御反应及代谢产物有关,还可能与内生真菌自身菌丝的定殖与生长有关。而关于内生真菌在禾草种子萌发及成苗阶段对病原菌侵入、扩展以及共生体拮抗物质生成的调控等抗病机理还需进一步探究。
综上所述,在不同胁迫处理下,内生真菌对宿主种子萌发各项指标均产生积极影响,说明内生真菌可提高宿主植物在萌发阶段非生物和生物胁迫抗性。但同时也存在内生真菌对宿主种子萌发有抑制作用和无显著影响的研究结果,其原因可能是内生真菌参与宿主禾草的抗性机制与植物基因型、内生真菌基因型、共生体适应性及生长环境密切相关。带菌种子在胁迫下萌发能力更强可能是由于其作为一种生物异源物质,在宿主内部形成长期胁迫,因此植物提前适应或提高了对外界环境的敏感度[70]。

表1 内生真菌对宿主在不同胁迫下种子萌发影响Table 1 Effects of grass endophyte on seed germination of hosts under different stresses
3 化感作用
化感作用(allelopathy)是指植物向其周围环境释放的某些化学物质对周围植物或微生物造成有利或不利影响的作用[72-74]。植物间的化感作用在种子萌发、幼苗存活、植物物种分布和种群结构及演替方面都发挥着作用[73]。同时,其与植物种类、生育期、生长部位、生态型、内生真菌侵染及化感物质浓度等有关[75]。研究发现内生真菌通过提高宿主植物的化感作用,进而提高宿主在种间及种内的竞争优势,以获取更多的营养和生长空间[75]。有些化感作用在种子萌发、幼苗存活、植物物种分布及种群结构和演替方面也发挥着积极作用[74]。草粉法[75]和浸提液法[76]是研究植物通过化感作用对其他植物种子萌发产生影响的常用方法,除此以外发酵液法和挥发油法也逐渐应用于化感作用的研究(表2)。
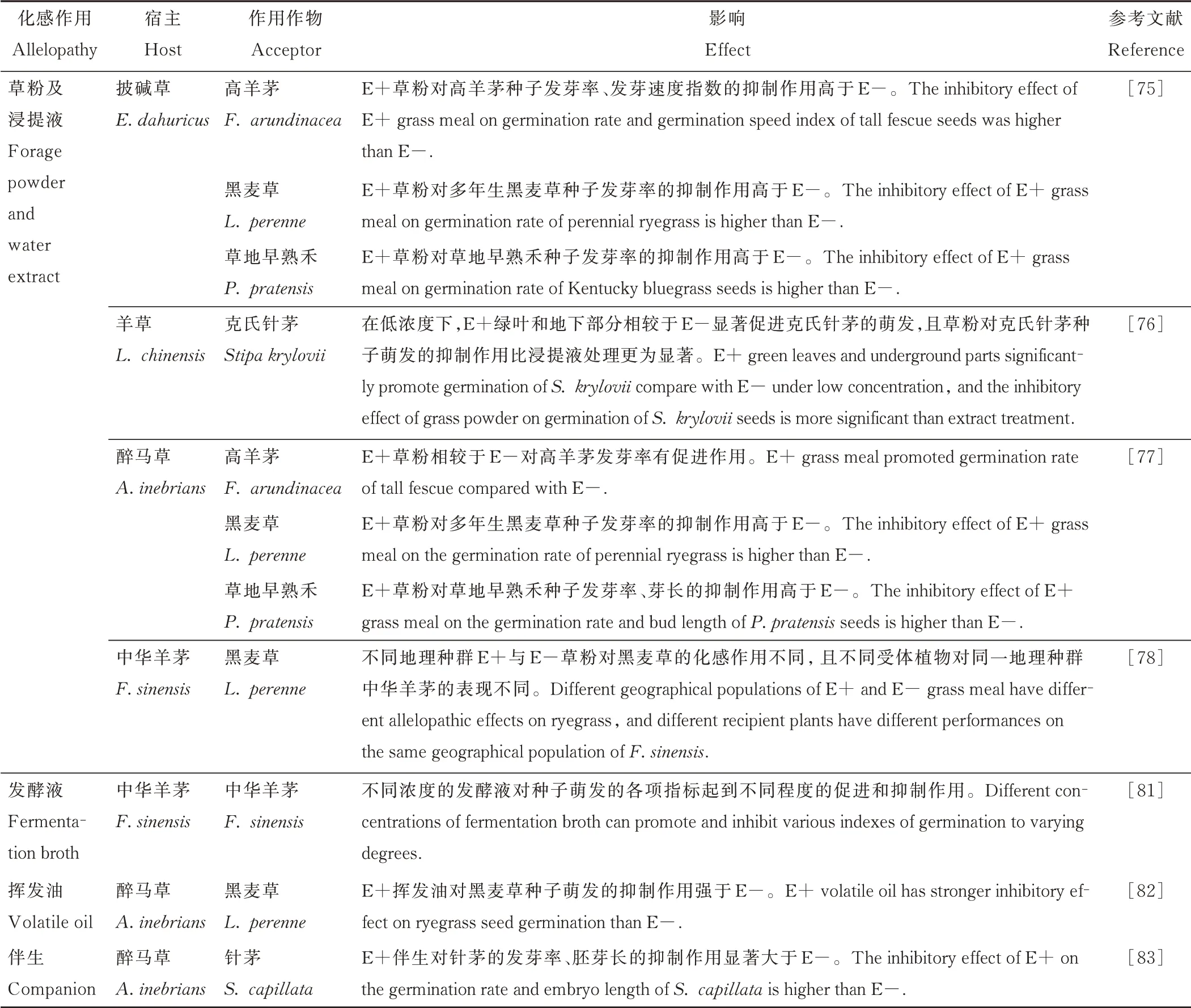
表2 内生真菌引起的不同化感作用对禾草种子萌发的影响Table 2 Effects of different allelopathy caused by grass endophyte on germination of grass seeds
3.1 草粉及浸提液
研究者通过将E+羊草(Leymus chinensis)制作成草粉并配制成一定浓度的稀释液,探究克氏针茅(Stipa krylovii)种子在不同浓度草粉液下种子萌发情况,发现相比于E-羊草草粉浸提液,E+羊草草粉浸提液整体上呈现出低浓度促进,高浓度抑制的效应[76]。杨松等[75]通过滤纸萌发试验表明,与E-相比,E+披碱草草粉浸提液能够抑制高羊茅、黑麦草、草地早熟禾(Poa pratensis)种子的发芽率及高羊茅种子的发芽速度;E+醉马草草粉浸提液可降低多年生黑麦草和草地早熟禾的发芽率,却可提高高羊茅种子的发芽率[77],但无论带菌与否,草粉液处理均会抑制此三者的萌发。然而,这些结果与盆栽试验结果并不一致。通过将采集自6 个不同地理种群中华羊茅制作成的草粉浸提液处理黑麦草种子,发现不同地理种群及带菌与否对黑麦草种子发芽率、发芽势及发芽指数影响有所不同,且不同受体种子对同一地理种群的中华羊茅效应也不同,但总体上表现为抑制趋势[78]。可见,禾草内生真菌共生体对不同禾草的化感作用存在差异,这种差异不仅与不同种禾草及内生真菌基因型有关,还可能与草粉浓度、草粉采集部位、采集生育期等因素有关。内生真菌与植物共生能在宿主植物中产生多种化感物质,这些化感物质通过调节植物自身离子平衡、水势和呼吸等机制以调节植物生理生化过程[76]。
3.2 发酵液
通过将黑麦草种子浸泡于分离自醉马草的甘肃内生真菌(Epichloë gansuensis)、分离自披碱草的内生真菌(E. bromicola)和分离自中华羊茅的中国内生真菌(E. sinensis)制成的发酵液并附加干旱处理,发现不同发酵液浸种后的黑麦草种子无论是否处于干旱胁迫下,其发芽率均可显著提高,而对于发芽势和发芽指数,不同内生真菌发酵液在不同干旱处理下对黑麦草种子萌发的效应不一致[79]。另外,内生真菌发酵液的浓度对其化感效应影响极大[80-81]。中华羊茅内生真菌发酵液稀释倍数对中华羊茅种子的萌发有不同的促进和抑制作用,其发芽率、发芽势在40 倍发酵液稀释下能显著提高,而在其他浓度下均表现为抑制作用[81]。关于内生真菌发酵液对禾草种子萌发影响研究较多,但仅有少量针对Epichloë属。内生真菌纯培养能产生一些次生代谢产物,通过优良菌株的筛选和微生物发酵[65],或许能够为生防资源研究提供新的思路。另外,内生真菌发酵液是否在多种附加处理下发挥作用尚不明确。已有的研究表明Epichloë内生真菌发酵液倍数及种类影响其化感作用,这为禾草内生真菌发酵液的开发利用提供实践基础。
3.3 挥发油
挥发油作为一种新的化感作用研究材料,通过将草粉在水蒸气蒸馏装置中蒸馏得到挥发性物质,经萃取、干燥得到粗提物。通过将E+、E-醉马草挥发油粗提物作用于黑麦草种子,发现相较于E-,E+醉马草挥发油粗提物对黑麦草种子萌发和幼苗生长有显著的抑制作用,即内生真菌增强了醉马草挥发油的化感抑制作用,同时随着挥发油浓度的增加,对黑麦草萌发指标的抑制作用也随之增加,且其幼苗生长差异也越来越大[82]。关于内生真菌挥发油的作用在多科、属植物上均有研究,这为我们研究禾草-内生真菌共生体对禾本科植物的化感作用机理以及利用优良禾草-内生真菌共生体对其他科属植物进行生长调节研究提供了一个新的思路。
3.4 伴生
同一群落中,各植物间对生态资源的竞争不可避免,故一种群落中某种植物的生长势必会对其伴生种产生影响。研究发现,带菌醉马草除自毒性以外,释放的化感物质对其伴生种也有影响。黄玺等[83]研究表明,带菌醉马草可抑制其常见伴生种针茅(Stipa capillata)及硬质早熟禾(Poa sphondylodes)种子萌发和幼苗生长。内生真菌的存在能够提高醉马草对营养物质的利用率,在促进自身生长的同时抑制其竞争者,从而表现出强的竞争力[40]。内生真菌侵染可增强禾草本身对其伴生植物的化感抑制作用,通过对禾草内生真菌共生体及其伴生植物资源调查,根据其生活史及生态位定义种群之间资源利用的相似与竞争等关系,进而判断在生态系统中种群及群落的演化方向,并对其可能出现的问题进行有针对性的研究。
异株相克,即一种生物释放出对另一种生物有抑制或促进作用的物质[84],是植物间化感作用基本原理,植物生长环境受到竞争、压力及干扰的综合影响,其中,竞争在决定群落性质方面扮演重要角色[85]。植物的竞争策略多样,种间差异、幼苗出土时间、空间分布、相邻植物体型、竞争期时间、生物因素、地上地下竞争、土壤养分、水分等,其中生物因素包括真菌和食草动物的作用是影响竞争的重要因子[83],内生真菌侵染能有效增强宿主植物的化感作用。然而,化感作用的方向、强度与内生真菌、宿主及受体植物基因型、二者生存环境以及化感物质的浓度和形式有关。植物自身含有萜类、黄酮、酚和酯等活性成分,这些成分在植物与其他物种竞争方面可能发挥着积极作用。然而,先前的研究结论得出的带菌植物对受体禾草种子萌发表现出“低浓度促进,高浓度抑制”的作用规律,这些作用的改变可能是由于内生真菌在宿主体内产生的生物碱、多酚类物质、有机酸、脂肪族类和醌类等活性物质参与到了植物的化感作用当中[82],进而调节或改变了宿主植物在植物各方面竞争当中的表现。
4 其他
4.1 CO2浓度
随着现代社会对化石燃料的大量消耗及对土地的不合理开发,大气中CO2浓度逐年升高,CO2是植物光合作用的原料,其浓度增加能够在一定程度上促进植物生长发育及增加干物质积累[86],而对于其在种子萌发过程中的作 用 报 道 存 在 差 异[87]。陈 薇 等[34]和 师 志 冰 等[87]分 别 通 过CO2和 内 生 真 菌 在 高 羊 茅 和 羽 茅(Achnatherum sibiricum)种子上的交互作用试验发现,内生真菌在羽茅种子发芽以及高羊茅幼苗长和幼根长的增益作用在高浓度CO2条件下更为显著;总的来说,CO2浓度的增加加大了E+和E-种子之间的生长差异[34]。CO2主要通过增加植物光合作用提高对内生真菌的光合供给,进而对禾草-内生真菌共生体产生影响,因此内生真菌与CO2互作的增益效应在萌发后才可能显现出来。通过分析CO2浓度对禾草-内生真菌共生体萌发的影响,有助于通过种子萌发情况预测全球气候变化,同时,也能够预测在全球气候变化中禾草-内生真菌共生体的竞争能力甚至是种群分布与优势物种[87]。因此,探究内生真菌与CO2互作机理及综合反应机制对将来禾草内生真菌共生体育种也有积极意义。
4.2 保存时间
内生真菌在种子中的活力变化受到种子储藏时间、温度及湿度的影响,随着种子储藏年限的加长,内生真菌在种子中的活力随之降低,当内生真菌完全丧失活力时,种子萌发将不再受到内生真菌的影响[88]。金文进等[89]研究发现,在同等贮藏年限下,E+中华羊茅种子的发芽指标与苗长、根长整体上显著优于E-,而对于过长的保存年限,内生真菌的增益效应并未体现。另外,有研究发现,在具有更高发芽率的草甸羊茅(Festuca pratensis)种子中,内生真菌活力也更高[90]。有研究对不同保存年限下醉马草种子进行萌发试验,发现内生真菌的侵染能缓解贮存年限对种子萌发的负效应。醉马草种子在贮藏1~2 年后,其发芽率、发芽指数、根长及苗长均显著降低,但E+种子萌发力始终强于E-。通过对其活力指标测定,发现内生真菌可能通过在宿主植物种子中的代谢活动,缓解质膜受损程度,提高种子脱氢酶和酸性磷酸酯酶活性,以延缓种子自然衰老进程[88]。内生真菌与宿主植物共生,二者共同消耗种子中的营养物质,随着贮藏年限的增加,内生真菌获得的营养物质逐渐减少,进而失去活力。因此可以得出结论,内生真菌对种子的增益效应是有限的。农事作业中,种子的贮藏不可避免,探究关于内生真菌在宿主种子中代谢活动的生理生化机制,并有针对性地进行育种工作,或许对今后农事生产具有重要价值。
5 展望
我国幅员辽阔,禾草种质资源丰富,有超过225 个属的禾本科植物,目前,已在早熟禾属、羊茅属、披碱草属等70 多种禾草中发现有内生真菌共生[14]。近年来,随着我国居民生活水平的提高及对环境绿化的重视,草坪业发展迅速,应用范围扩大,逐渐发展为人类生态系统的重要组成部分之一[91]。随着对内生真菌与宿主禾草共生关系及互作响应研究的加深,内生真菌对宿主禾草在生长发育及抗性生理上的增益作用日益体现,内生真菌对种子萌发的影响也越来越受到广大研究者的关注。有研究表明,与石南科植物共生的内生真菌可以通过菌丝侵入破壁进而促进种子萌发[92],能否以此为参考,发掘出促进禾草种子萌发的内生真菌,研究其多样性、专一性、共生及对环境胁迫的抗性萌发机制,对利用内生真菌开展禾草植物种质创新、物种保护及繁育研究等有重大意义。
种子萌发在禾草农业发展中扮演重要角色,通过了解种子萌发特征才能进一步对植物进行深入的研究与开发。种子萌发需要适宜的水分、温度等环境因素,这些环境因素协同作用,共同调节种子萌发。然而近年来,全球环境逐渐恶化,生物赖以生存的自然环境资源面临各种威胁,其中以盐胁迫和重金属胁迫问题最为突出,我国盐渍土面积约1×108hm2,其分布范围广,类型多[93],同时每年因重金属污染造成的直接经济损失高达200 多亿元[94],因此,提高禾草种子在不同环境胁迫下的萌发率是草地禾草建植的关键问题之一,也是禾草能否在胁迫环境中生存的基础。内生真菌与禾草共生能促进禾草种子在胁迫环境下萌发,对于如何提高禾草种子在不良条件下的萌发指标,除了改良利用现有土地和其他环境资源以及采用对应的工程措施外,利用内生真菌资源对禾草种子进行种质创新,通过分子生物学、生理生化机制以及基因改良手段,深入研究在胁迫环境中,蛋白质分子和基因在不同时空条件的选择性表达,从根本上探究内生真菌提高禾草萌发抗性能力,以达到有效利用内生真菌资源和改善胁迫环境的目的。
种子休眠(seed dormancy)是指有生活力的种子在适宜萌发环境下不能萌发的现象[95]。就植物而言,种子休眠能够使其规避地震、沙尘暴等自然灾害及抵御旱涝、冷热等特殊气候情况,以保证物种延续,是植物生态适应的特征,但这却阻碍了禾草育种的发展,其对野生物种的人工繁育阻碍尤为明显。种子休眠是由遗传和环境共同影响,国际社会在破除禾草种子休眠研究方面有一定基础,但种子的萌发与休眠是一个复杂的生理过程。通过研究禾草种子解剖结构发现,种子附属物,如稃和种皮是影响种子萌发的重要因素,其可能对胚根胚芽的伸长产生机械阻碍,抑制水分和气体交换,阻碍种子内部抑制物的外漏或调整种子对光的响应[95],在石南科植物种子中发现,带内生真菌的种子菌丝能够突破外种皮,侵入到种皮与胚之间,胚径向伸长,根分化且被菌丝覆盖,在胚胎发育成熟期,菌丝进入细胞表皮外切向壁以进行营养交换,而未与内生真菌共生的种子其种皮厚且存在包被[92]。内生真菌菌丝在禾草种子中较多分布于内表皮,其在种子萌发进程中除了起到促进植物激素产生的作用,在突破种皮和通过菌丝吸收营养物质方面是否发挥作用还需进一步研究。内生真菌共生能够提高宿主的化感作用,但作用效果与方向和植物种类、化感物质浓度有关,研究作物及牧草不同品种间的化感作用,进一步研究化感作用在种间差异机制,通过选育合适的禾草-内生真菌共生体作为绿肥作物或覆盖作物,以在草坪或农田中抑制杂草种子萌发、防治杂草危害和促进优质草坪建植。
内生真菌对禾草种子在不同生境下萌发影响的分子机理是今后研究的方向之一。结合传统育种方法和现代分子生物学技术,明确内生真菌对宿主禾草萌发有利影响的生理生化机制,充分利用禾草内生真菌对特定宿主的优良特性,对禾草进行定向改良[96],建立针对不同生境及需求的禾草内生真菌共生体种质资源库,对内生真菌在禾草育种作用方面有重大意义。研究发现内生真菌基因组中存在能够使家畜中毒的基因[97],这为利用内生真菌进行牧草育种带来新的挑战,内生真菌对禾草种子萌发的影响不应仅关注于良好萌发指标这一浅层研究,通过对内生真菌在禾草种子中的分布及其基因组中产碱基因、抗性基因和共生基因特征及表达的研究,分析其在禾草种子萌发过程中发挥的作用、萌发后对环境适应性及对采食者甚至人类的潜在危险进行风险评估,以期得到有利于动物健康及生态建设的目标共生体。现有的内生真菌对宿主禾草种子萌发影响多为胁迫环境下生理响应、生长指标及生态位竞争等宏观方面,而对造成这些差异的分子机理分析还存在欠缺,因此,针对微观机理的研究具有较大潜力。目前,随着内生真菌共生对不同科种子萌发影响及内生真菌对宿主植物抗性影响的分子机制研究的不断深入,为选育具有优良特性的禾草种质提供了更多启示。因此,应结合宏观生理指标与微观分子机制针对禾草植株的需求特性、物种特异性和内生真菌功能基因多样性,分别选育出满足不同需求的禾草内生真菌共生体,以将内生真菌资源充分利用。
