不同物候期刈割对羊草再生和根形态及产量的影响
2022-03-26赵成振李强钟荣珍
赵成振,李强,钟荣珍*
(1. 中国科学院东北地理与农业生态研究所,吉林省草地畜牧重点实验室,吉林 长春 130102;2. 中国科学院大学,北京 100049)
草原是分布广泛的陆地生态系统,占全球陆地面积的26%[1]。在温带气候区,以草原为基础的乳品生产系统为牲畜提供了可持续的食物[2]。农民通常每年8 月下旬收获牧草[3]。尽管传统的草地利用方式满足了草地保护的需要,但是草地产量难以满足日益增加的牲畜的需求[4]。因此,传统的草原管理限制了牧草的生产和利用[5]。许多研究表明春季刈割可能影响牧草的再生和干物质(dry matter,DM)总产量[6]。比如4 月刈割比5 月刈割降低了“shimanoushie”全年的DM 总产量[7],然而也有研究表明5 月20 日刈割比6 月20 日刈割增加了柳枝稷(Panicum virgatum)的全年DM 总产量[8]。此外,有研究表明传统利用方式8 月中旬割1 次,牧草DM 产量更高[9-10]。
过早刈割可能会对羊草(Leymus chinensis)累积DM 产量产生负面影响[11]。在松嫩草地,从8 月15 日至10 月4 日期间每隔10 d 设置1 个刈割处理,结果表明刈割没有对羊草次年的DM 产量产生影响[3]。因此,刈割时间越晚,对草地的伤害越小。而最佳刈割期对草地生产至关重要,是获得高产的关键。羊草通常在8 月15 日(传统刈割日)之前经历伸长期、抽穗期和花期[12],这些时期的刈割管理可能有助于累积DM 和粗蛋白(crude protein,CP)产量。
刈割理论及试验结果表明,植物有能力通过形态和生理调整,如再生分蘖,产生更多的新叶,光合能力的增强,最后导致再生速率的增加[13-14]。在地下部分,根形态和根茎通常被认为是决定大针茅(Stipa grandis)和羊草再生性能的主要养分吸收器官和养分储存器官[15-16]。不同时期刈割对根茎水溶性碳水化合物(water soluble carbohydrate,WSC)储量和根系发育有强烈影响[17-18]。不同时期刈割,羊草的光合能力、营养物质在地上和地下器官之间的分配以及储存养分的有效性也存在差异[19]。因此,不同时期刈割会影响牧草的再生,进而影响其DM总产量。此外,有研究表明,轻度或中度刈割会显著影响根系结构,尤其是增加后续再生过程中的细根长度[20]。本研究通过对不同直径的根茎形态及根茎WSC 含量的研究,探究了拔节期、抽穗期和开花期刈割管理对羊草累积DM 产量、CP 产量和可持续的影响。
羊草是一种原生的多年生根茎型草本植物,以羊草为主的典型草原分布在欧亚温带草原带上[21]。羊草是大型草食动物的首选牧草,其品质优良,适口性和营养价值高[3]。本研究旨在研究羊草在不同物候期刈割的再生过程,并根据其地下根系形态和根茎WSC 含量探究羊草草地适宜的刈割期。研究结果为合理制定温带草地刈割制度提供了理论参考。
1 材料与方法
1.1 研究区概况
本研究在中国科学院东北地理与农业生态研究所长岭草地农牧生态研究站(44°33′N,123°31′E,海拔145 m)进行。研究区属温带大陆性季风气候,年平均气温5.9 ℃,平均最高和最低气温分别为6.4 和4.6 ℃,年平均生长期约为150 d,>10 ℃的年有效积温为2900~3000 ℃。年平均降水量为427 mm,70%的降水发生在6-9 月(图1)。根据世界土壤资源[22]数据,研究区土壤为盐碱型,土壤pH 值为8.7。研究区主要的植被类型为羊草,是一种多年生根茎型C3草本植物,其生物量占该地区地上总生物量的85%。试验区为0.3 hm2的平坦未退化草地,羊草分布均匀,近十几年仅少量放牧。
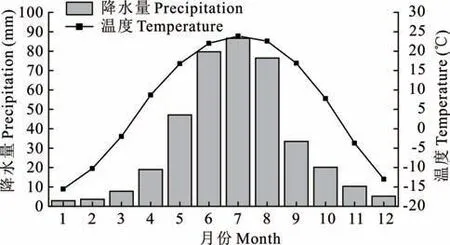
图1 松嫩平原2000-2016 年平均降水量和气温分布Fig. 1 Mean precipitation and temperature distribution of Songnen plain from 2000-2016
1.2 试验设计
试验在2018 年开始实施,包括对照组(control,CON)、刈割组包括拔节期刈割(elongation stage at May 15,ES)、抽穗期刈割(heading stage at June 1,HS)和开花期刈割(flowering stage at June 15,FS),留茬高度为5 cm。对照组不刈割,8 月15 日收获;刈割组起始刈割后,8 月15 日收获。试验采用随机区组设计,每个区组面积为4 m×35 m,3 次重复,共计12 个区组。为防止相互影响,区组之间设置1 m 宽的隔离带。每个区组内分成7 个小区,每个小区面积为4 m×5 m。
1.3 取样方法
1.3.1 地上生物量取样 CON 处理,取样时间从5 月15 日开始,每隔15 d 取样1 次,到8 月15 日(传统制作干草的收割日期)结束。对于刈割处理,起始刈割开始前进行第1 次取样,作为刈割开始前的羊草产量,起始刈割后每隔15 d 取样1 次,至8 月15 日收获后为最后1 次取样,作为刈割后羊草再生产量的动态变化。因此,在生长期内,CON、ES、HS 和FS 分别采样7、7、6 和5 次。每次取样时,每个处理每个区组中依次选择1 个小区(未被取过样),每个小区随机取3 个1 m×1 m 的样方框,采用手工方式剪草,每个样方框内的羊草装入信封中并放入105 ℃烘箱杀青30 min 后,再在65 ℃烘箱中干燥48 h,称重,研磨后过0.4 mm 筛,4 ℃保存,用于氮(nitrogen,N)的分析。
1.3.2 根系及单株取样 在8 月中旬进行根系及地上单株取样。每个区组随机选取3 个小区,分别挖出3 个20 cm×20 cm×30 cm 的土块,连同地上部分转移到实验室。然后将每个土块的地上羊草在茎基部剪断,从中随机选取10 株羊草个体分别按茎、叶、鞘装到穿孔纸袋中,并放入105 ℃烘箱杀青30 min 后,再在65 ℃烘箱中干燥48 h,分别按茎、叶、鞘称重。每个区组30 株羊草的茎、叶、鞘的平均值为1 个重复,采用3 个重复(3 个区组)进行数据分析。将各区组每个小区的土块上剩余植株样品收集按茎、叶、鞘冷冻干燥后研磨成细粉,过1 mm 筛,-20 ℃保存,用于测定WSC 含量。取每个区组3 个样品的茎、叶、鞘的WSC 含量的平均值为1 个重复,用3 个重复(3 个区组)进行数据分析。随后,将每个小区的土块放入水中,小心冲洗,将目标植株的根系与其他根系分离,置于0.2 mm 的筛网上。每株羊草完整的根中含有根状茎(将连通的羊草从其节间的中间切下)和须根。每个区组中每个小区保留10 个完整的根,并保存在自来水中,置于冰箱(8 ℃)中,直到进行进一步的分析。在平板扫描器(Epson Scan 700/750,加拿大)上,以400 dpi 的分辨率扫描每株羊草的根系,使用WinRhizo 2005c 软件(Regent Instruments,Quebec,加拿大)对图像进行分析,计算出每株羊草在不同根直径(root diameter classes,RDC)下(0.0~0.2 mm、0.2~0.5 mm、0.5~1.0 mm、1.0~2.0 mm 和>2.0 mm)的根长、根表面积及根体积。每个区组30 株羊草的根长、根表面积和根体积的平均值为1 个重复。收集每个小区的土块剩余的根系用于WSC 含量的分析,每个区组3 个样品的根WSC 含量的平均值为1 个重复。
1.3.3 光合特性的测定 叶片的净光合速率、气孔导度、胞间CO2浓度和蒸腾速率采用LI-6400 便携式光合系统(Licor,美国)测定。在空气温度25 ℃、CO2浓度为365×10-6、光合活性辐射(photosynthesis active radiation,PAR)为0~2000 μmol·m-2·s-1的条件下,使用LI-6400 的程序生成各叶片的光合响应曲线计算光合速率。为避免强烈的阳光和大风,测量于8 月15 日上午09:00-11:00 进行。每个区组随机选择3 个小区,每个小区随机选取3 株羊草测定其中间叶片代表整株羊草,每个区组9 株羊草的平均值作为1 个重复,用3 个重复(3 个区组)进行数据分析。
1.4 N、CP 和WSC 含量测定
用CHN 元素分析仪(EA1108,Series 1)分析总N 含量,CP 含量为N 含量×6.25。CON 的DM 总产量为8 月中旬羊草产量达到高峰时的草产量。刈割组DM 总产量为起始刈割DM 产量与收获DM 产量之和。CP 产量为DM 产量乘以CP 含量,总CP 产量计算方法同DM 总产量。WSC 含量的测定采用苯酚-硫酸比色法[23],每个样品中准确称量0.05 g,在玻璃管中加入20 mL 去离子水,静置30 min,然后加热至沸腾(100 ℃)保持1 h,离心后,取样品中的上清液用于茎、叶、鞘和根的测量分析,每个样品重复3 次。采用96 孔平底聚苯乙烯酶标仪(型号:354,Titertek Multiskan Ascent,德国),在波长490 nm 处记录吸光度,测定WSC 的含量。WSC 含量由以下公式得到:
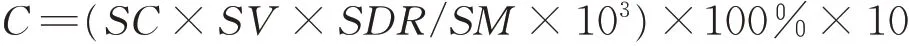
式中:C为WSC 含量(mg·g-1);SC(sample concentration)为样品浓度(mg·L-1);SV(solution volume)为溶液体积(mL);SDR(sample dilution multiple)为样品稀释比;SM(sample mass)为样品质量(mg)。
1.5 数据分析
采用SPSS 16.0 统计软件和Origin 2016 Pro 软件进行方差分析和绘图。采用t检验评价4 个处理下羊草植物组分中DM 产量、CP 产量、植物功能性状、光合作用、根系形态及其WSC 含量的显著差异。用最小二乘平均值,并以平均值±标准误表示。P<0.05 时差异有统计学意义。
2 结果与分析
2.1 羊草产量
总体而言,随着刈割从拔节期延迟至开花期,羊草再生DM 产量迅速增加,特别是ES 和HS 处理之间的 再 生DM 产 量 接 近(图2)。6 月1 日 至8 月15 日,CON 组羊草CP 产量为10.12~13.21 g·m-2。在整个再生季中,FS 处理羊草再生CP 产量从1.11 g·m-2稳定增长到17.45 g·m-2,ES 和HS 处理的再生CP 产量先增加,8 月1 日达到峰值后开始下降。
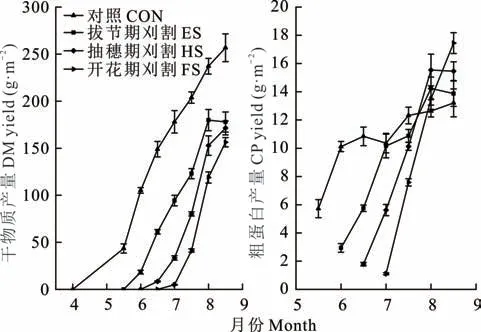
图2 不同物候期刈割羊草再生干物质产量和粗蛋白产量的变化Fig. 2 Changes of regrowth DM yield and CP yield of L.chinensis under different phenological growth stages mowing(mean±SE,n=3)
与CON 处理相比,ES 处理的累积DM 产量下降了12%(P<0.05),HS 和FS 处理的累积DM 产量分别增加了9%和29%(P<0.05)(图3)。各刈割处理的CP 累积产量均高于CON 处理(P<0.05)。随着刈割期的推迟,羊草累积CP 产量递增,HS 和FS 处理之间没有显著差异。
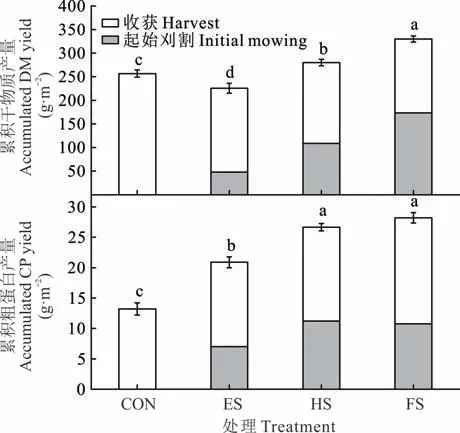
图3 不同物候期刈割羊草累积干物质产量和粗蛋白产量的变化Fig. 3 Changes of accumulated DM yield and CP yield of L.chinensis under different phenological growth stages mowing(mean±SE,n=3)
2.2 羊草根系及植物性状
羊草根直径从0 增加到1 mm 时,不同处理间的根长、根表面积和根体积差异显著(P<0.05),当根直径超过1 mm 时,各处理间无显著差异(表1)。与CON相比,ES 处理显著(P<0.05)降低了羊草根直径在0~0.2 mm 和0.2~0.5 mm 的根长和根体积。HS 处理与CON、FS 处理在上述指标上无显著差异,但是HS和FS 处理在根直径为0.5~1.0 mm 时,羊草的根长、根表面积和根体积均显著高于CON 处理(P<0.05)。
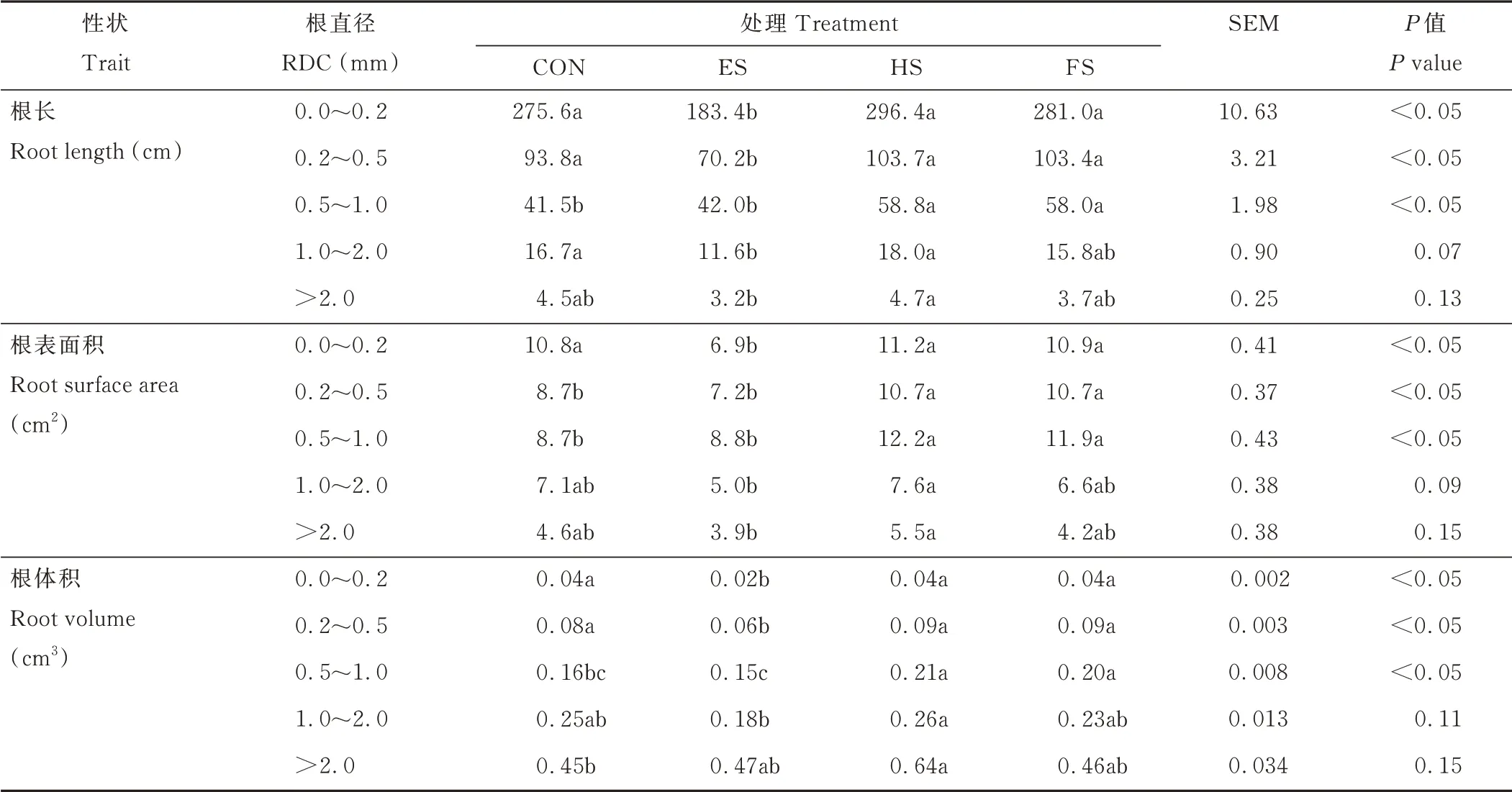
表1 不同物候期刈割羊草根长、根表面积和根体积的变化Table 1 Changes of root length,surface area and volume of L.chinensis under different phenological growth stages mowing(n=3)
刈割组与对照组相比,羊草茎、叶和鞘生物量呈降低的趋势,但是在羊草生物量分配格局中,其叶片比例平均增加了14%,茎比例平均减少了10%左右(P<0.05,图4)。
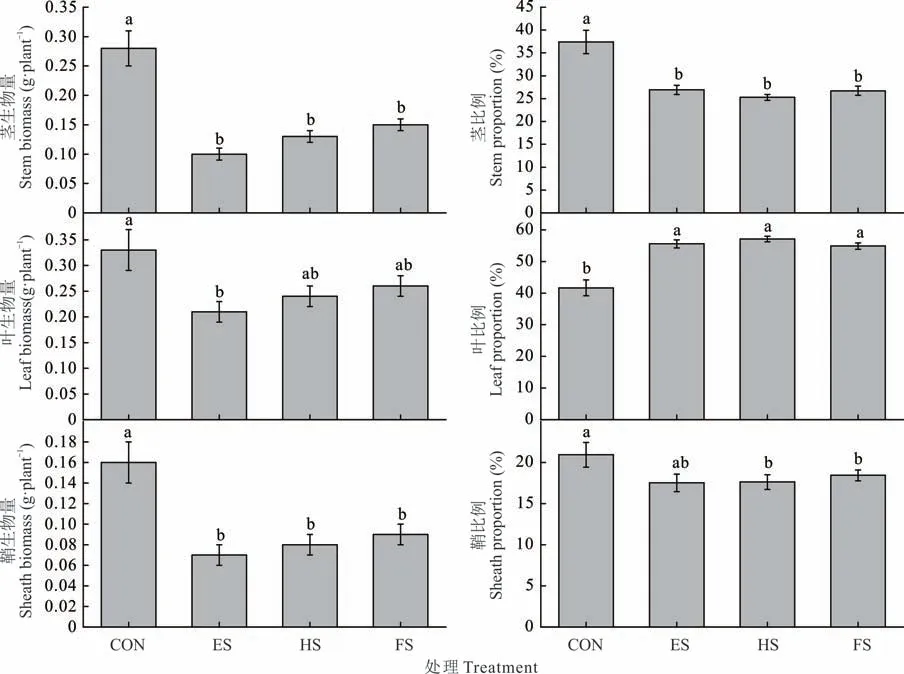
图4 不同物候期刈割羊草个体茎、叶和鞘生物量及其比例的变化Fig. 4 Changes of stem,leaf and sheath biomass of individual L. chinensis and their proportions under different phenological growth stages mowing(mean±SE,n=3)
2.3 光合作用
羊草净光合速率、气孔导度和胞间二氧化碳浓度在FS 处理最高(P<0.05,图5)。但在净光合速率、气孔导度和蒸腾速率方面,ES 和HS 处理与CON 无显著差异。
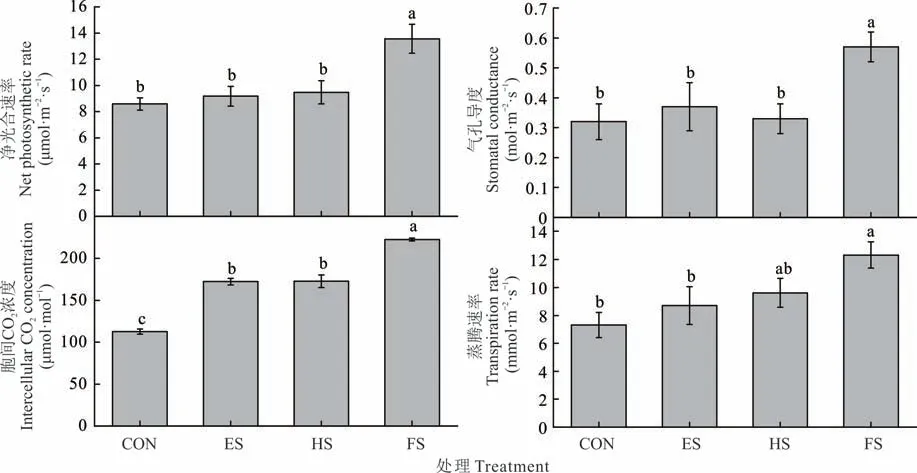
图5 不同物候期刈割羊草净光合速率、气孔导度、胞间CO2浓度和蒸腾速率的变化Fig. 5 Changes of net photosynthetic rate,stomatal conductance,intercellular CO2 concentration and transpiration rate of L.chinensis under different phenological growth stages mowing(mean±SE,n=3)
2.4 WSC 含量变化
随着初始刈割期从ES 期延迟到FS 期,羊草叶片WSC 含量呈增加趋势(图6),而FS 处理根WSC 含量则降到最低(P<0.05)。茎和鞘在各处理间无显著差异。
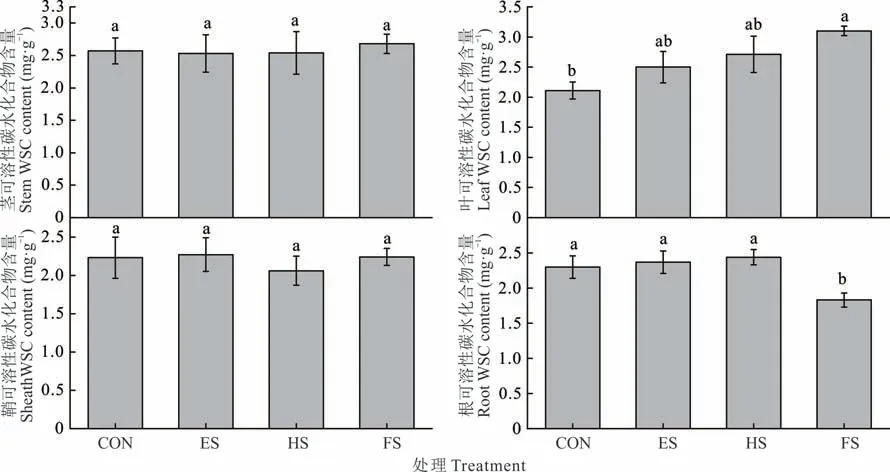
图6 不同物候期刈割羊草茎、叶、鞘和根WSC 含量的变化Fig. 6 Changes of stem,leaf,sheath and root WSC contents of individual L. chinensis under different phenological growth stages mowing(mean±SE,n=3)
3 讨论
羊草是欧亚东部温带草原的优势种,对环境反应敏感[21]。本研究结果表明,不同物候期刈割羊草的再生响应既有相似性,也有差异性。刈割后植物通常表现出较高的地上生长率,这种现象已经被广泛观察到[24-25]。植物在整个再生过程中已经进化出一套形态和生理机制来应对刈割造成的伤害[26-27]。ES 处理羊草再生DM 产量增长缓慢,根据之前的研究,这可能是由于羊草在拔节期养分贮藏量低所致[28-29]。更重要的是,根系对养分的吸收和同化能力是羊草再生的限制因素[30-31],ES 处理下羊草根的长度、表面积和体积在根直径0.0~0.2 mm 处最低。这种根被认为是细根[32],在资源获取方面具有重要作用[33]。刈割组羊草DM 均迅速增加,ES 处理羊草DM 再生产量在8 月1 日达到峰值后开始下降,但是FS 处理DM 再生产量仍保持增长,这和其净光合速率高于其他处理的结果保持一致,但FS 处理羊草根WSC 含量最低。这可能因为羊草根中WSC 的储量与植物再生进程有关[34],随着再生期的延长,羊草地上生长速度开始下降,此时羊草进行了权衡以保证更多的WSC 被分配到根部,以确保它有足够的养分储存,而不是优先增加生长[35-36]。这一结果表明,与其他处理相比,FS 处理不利于羊草WSC 的积累,可能是没有足够的再生时间。
尽管羊草的再生过程和形态不同,但不同物候期刈割时其再生DM 产量是接近的,特别是ES 和HS 处理,这是因为较长的再生周期被较低的相对再生率所抵消。然而,不同物候期刈割羊草产量差异显著,因此,总DM 累积量主要由刈割前的DM 来决定。总CP 累积产量在4 个处理中不成比例的增加,其中在HS 和FS 处理之间仅增加了1.54 g·m-2。这一结果表明,总CP 累积产量是有限的,并没有与总DM 累积量同步。在之前的研究中也观察到了类似的结果[37]。本研究结果表明,早刈割不利于羊草DM 累积和根系生长发育,晚刈割则不利于羊草根茎WSC 的积累,HS 刈割处理能够获得较高的DM 累积量,且对羊草根系的损害较小,保证羊草的可持续性生长。此外,从粗蛋白产量的角度来看,HS 刈割处理粗蛋白产量显著高于ES 刈割处理,而FS 处理粗蛋白产量相较于HS 刈割处理增加不显著,因此,HS 处理能够获得较高的粗蛋白产量。
在本研究中,与对照组相比刈割组导致羊草个体茎、叶和鞘生物量显著减少。前人的研究也表明,这一现象与刈割导致的矮化形态特征相一致[12]。研究结果还表明,与CON 相比,刈割后分配到叶片上的生物量要多于茎上的生物量。这种植株大小介导了N、P 和C 优先分配到叶片而不是茎[38-40]。这种现象被称为“功能平衡”,植物通过刈割增加对叶片的养分分配来获得更多的光照资源[41]。
4 结论
本研究结果表明,随着刈割时间的推迟,DM 和CP 累积量显著增加。刈割组各处理羊草再生DM 产量接近,但其形态特征和再生后的生理变化不同。在此过程中,ES 处理限制了根直径在0.0~0.2 mm 和0.2~0.5 mm 下的根长、根表面积和根体积的增加,FS 处理则不利于WSC 积累。与CON 处理相比,HS 刈割处理可以获得较高的DM 产量,且对羊草的损害较小。
