马法兰治疗视网膜母细胞瘤作用机制初步研究
2022-03-23陈星宇文旭洋贾仁兵葛盛芳
陈星宇 文旭洋 贾仁兵 葛盛芳
视网膜母细胞瘤是儿童最常见的眼恶性肿瘤,占儿童恶性肿瘤的3%[1,2]。随疾病进展,视网膜母细胞瘤可出现局部侵袭和远处转移,其中颅内转移是造成患儿死亡的主要原因[3]。通过早期筛查、诊断及治疗,可有效提高视网膜母细胞瘤患者保眼率[4]。视网膜母细胞瘤有多种保眼治疗手段,包括全身化疗、动脉介入化疗、敷贴放射治疗和局部治疗(激光、冷冻、玻璃体腔注射等)[2]。近来发现,动脉介入化疗可在不影响患儿生存率的前提下,显著提高保眼率[5]。此外,玻璃体腔内注射化疗药物对视网膜母细胞瘤玻璃体播散疗效明显[6]。马法兰是动脉介入化疗和玻璃体腔注射治疗最常用的药物,既往应用于乳腺癌、卵巢癌、多发性骨髓瘤等多种恶性肿瘤的全身化疗[7]。马法兰是一种烷化剂,可烷化DNA中的鸟嘌呤,与DNA发生交叉连接抑制蛋白合成[8]。但马法兰在视网膜母细胞瘤中的作用机制尚不明确,明确马法兰在视网膜母细胞瘤中的作用机制具有重要意义。
随着二代测序技术的发展,RNA-测序(RNA-sequencing, RNA-seq)已成为分析基因表达水平的重要手段[9]。在肿瘤中,该技术常被应用于检测基因表达水平及鉴别新基因和转录因子。在本研究中,我们将应用该技术,检测mRNA表达谱,并对其进行生物信息学分析,探索马法兰在治疗视网膜母细胞瘤中的作用机制。
资料与方法
一、视网膜母细胞瘤细胞系来源及培养方法
实验研究。视网膜母细胞瘤细胞SO-RB50来源于广州中山眼科中心,细胞在含10%胎牛血清(Thermo Fisher scientific)的RPMI1640(Gibco)培养液中培养,培养箱的培养条件为温度37 ℃,CO2浓度为5%。
二、马法兰与SO-RB50细胞系联合培养及取样分析
实验组为SO-RB50细胞与1.107 nM的马法兰联合培养,对照组SO-RB50细胞接受相对应浓度DMSO处理。分别在4 h,8 h,12 h以及24 h进行取样分析,通过Nanodrop2000检测所提RNA的浓度和纯度,通过琼脂糖凝胶电泳检测RNA完整性,通过Agilent2100 测定RIN值。质检合格后进行mRNA测序(mRNA sequencing, mRNA-seq)。
三、生物信息学技术分析测序
对测序结果进行图像识别,去污染,去接头。通过RNA测序最大希望算法(RNA-Seq by Expectation-Maximization,RSEM)将转录组数据比对至人类参考基因组上,对比对结果进行统计量化,得到每个样品比对到每个转录本上的reads数目。基于原始的reads数据,进行标准化(normalization),进而得到每个基因的表达值。然后,对不同组间差异表达基因计算及多重假设检验校正,得到FDR值(错误发现率)和log2|FC|(log2|Fold Change|)。筛选标准为FDR<0.05,log2FC>1(上调基因)或log2FC<-1(下调基因)。获取马法兰处理后不同时间点的差异表达基因,对其进行GO富集分析(DAVID: https://david.ncifcrf.gov/tools.jsp),通过R语言对时间序列上相同变化趋势的基因进行聚类,同时进行KEGG通路富集分析。
结 果
一、时序差异表达基因
经过标准化预处理后,所有的样品测序深度一致,以FDR<0.05和log2|FC|>1为标准,获得各时序的差异表达基因。火山图显示各时序的上调基因和下调基因(图1A)。火山图和韦恩图显示,马法兰在前4 h引起1 723个基因持续发生差异表达,前8 h引起213个基因持续发生差异表达,前12 h引起344个基因持续发生差异表达,24 h仅360个基因发生表达量的变化(图1B)。可见,马法兰对基因表达影响最显著的作用时间为前4 h。
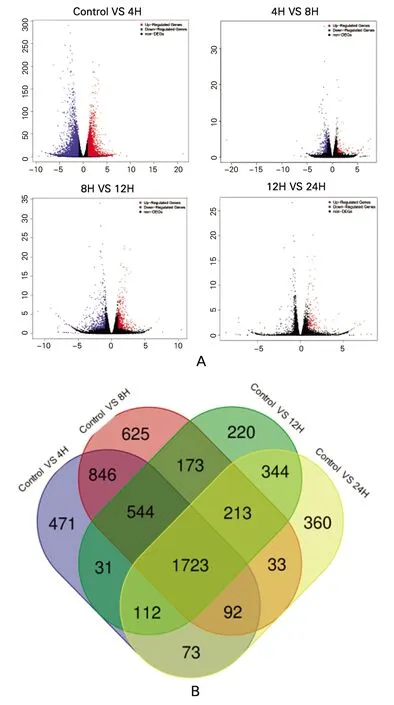
图1 时序的差异表达基因A示马法兰处理后各时间点样本进一步差异表达变化的基因。(红色圆点为表达上调差异表达基因FDR<0.05和log2FC>1,蓝色圆点为表达下调的差异表达基因FDR<0.05和log2FC<-1,黑色小点为表达未发生差异改变的基因) B示马法兰处理后各时间点样本进一步差异表达变化的基因数量
二、差异表达基因功能注释及富集分析
近一步通过GO富集分析马法兰处理4 h后引起的差异表达基因(DGEs),发现主要在以下几个方面发生改变:在生物学过程方面,差异表达基因主要涉及细胞进程、代谢过程、生物调节等;在细胞组分方面,差异表达基因主要影响细胞、细胞器和细胞膜等成分;在分子功能方面,差异表达基因主要富集在导致结合和催化活性等生物过程(图2A)。其中蛋白结合和细胞进程调控的改变最为显著(图2B)。对比不同时间点发现,马法兰处理8 h对比处理4 h,差异表达基因涉及突触相关的改变(图2C),马法兰处理12 h相比处理8 h,差异表达基因参与解剖结构相关改变(图2D),马法兰24 h相比处理12 h,差异表达基因影响生长和极性相关改变(图2E)。
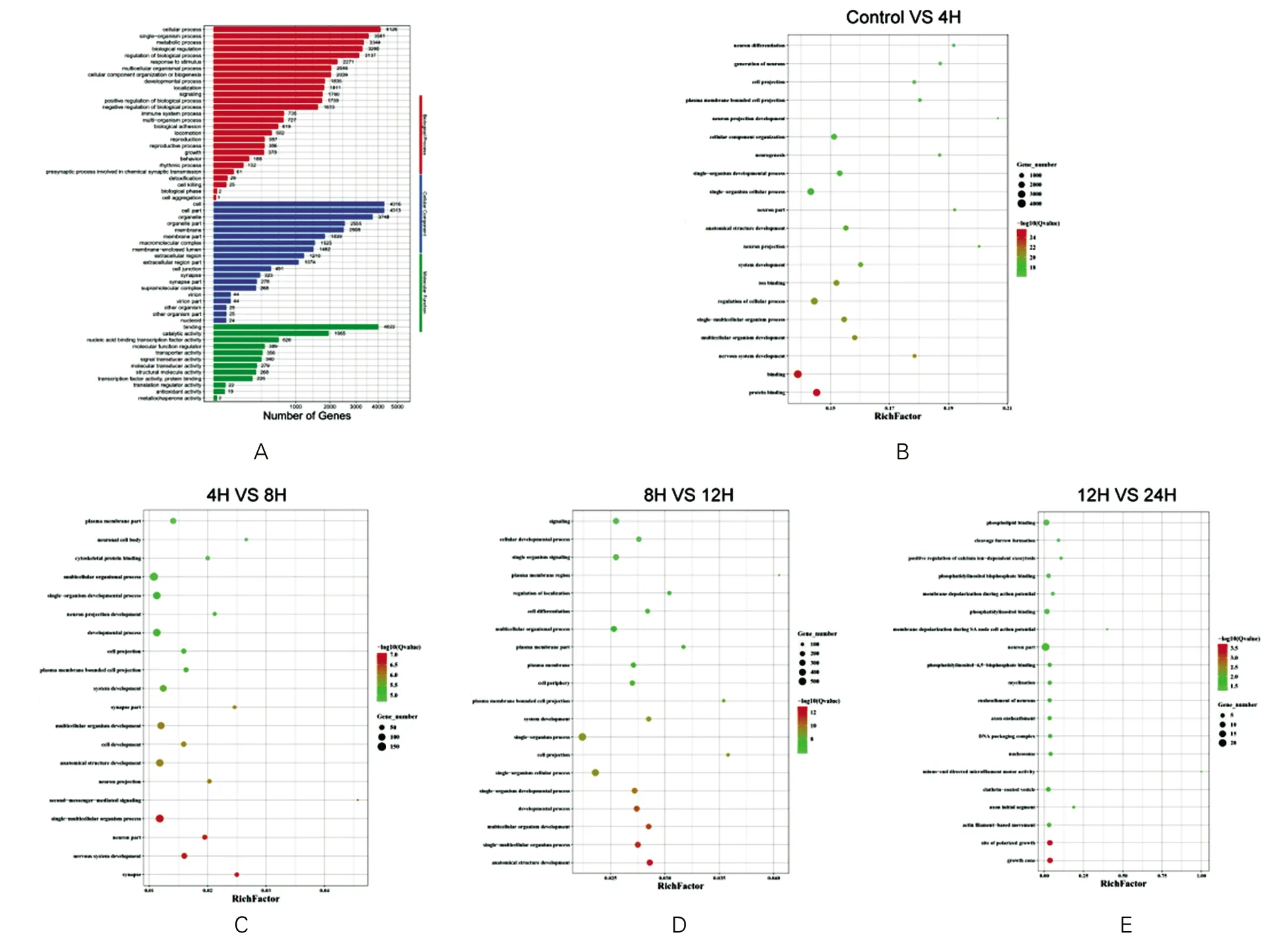
图2 差异表达基因的富集分析及功能注释A示马法兰4 h处理后差异基因GO富集情况 B-E示马法兰处理后各时间点样本差异基因的GO富集情况横坐标:富集指数,点的大小和颜色代表富集的显著性,点越大,颜色越深,表示富集程度越高;纵坐标:马法兰处理后各时间点样本差异基因的GO富集情况
三、相同变化趋势基因的聚类分析
通过R语言对在时间序列上相同变化趋势的基因进行聚类分析发现:DEGs一共可分成8类不同变化趋势的基因簇(图3A)。进一步分析发现:在这八类变化趋势的基因簇中,第2簇和第6簇的基因在马法兰作用后表达量分别迅速下调和上调,并于4 h后表达量维持不变;第7簇和第8簇的基因随马法兰作用时间延长,表达量不断持续升高或降低;第3簇的基因则在马法兰作用早期(4 h)表达量不变,在8 h后表达量显著升高(图3B)。

图3 差异表达基因聚类分析A示热图上相同变化趋势基因聚集分析,分成8个基因簇 B示8个基因簇在马法兰用后不同时间表达水平连续变化情况
四、不同簇基因KEGG功能富集及网络搭建
对不同簇的基因分别进行KEGG通路富集分析发现,马法兰作用后表达量迅速降低的第2簇基因主要富集于剪接体通路和核糖体生物发生通路(图4A),而表达量逐渐降低的第8簇基因主要富集于同源重组通路和DNA复制通路(图4E);马法兰作用后表达量迅速升高的第6簇基因主要富集于磷酸肌醇代谢通路和蛋白聚糖通路(图4C),表达量逐渐升高的第7簇基因主要富集于蛋白酶体通路(图4D);第3簇基因于马法兰作用8 h后显著升高,主要富集于类固醇生物合成通路和萜类化合物生物合成通路(图4B)。
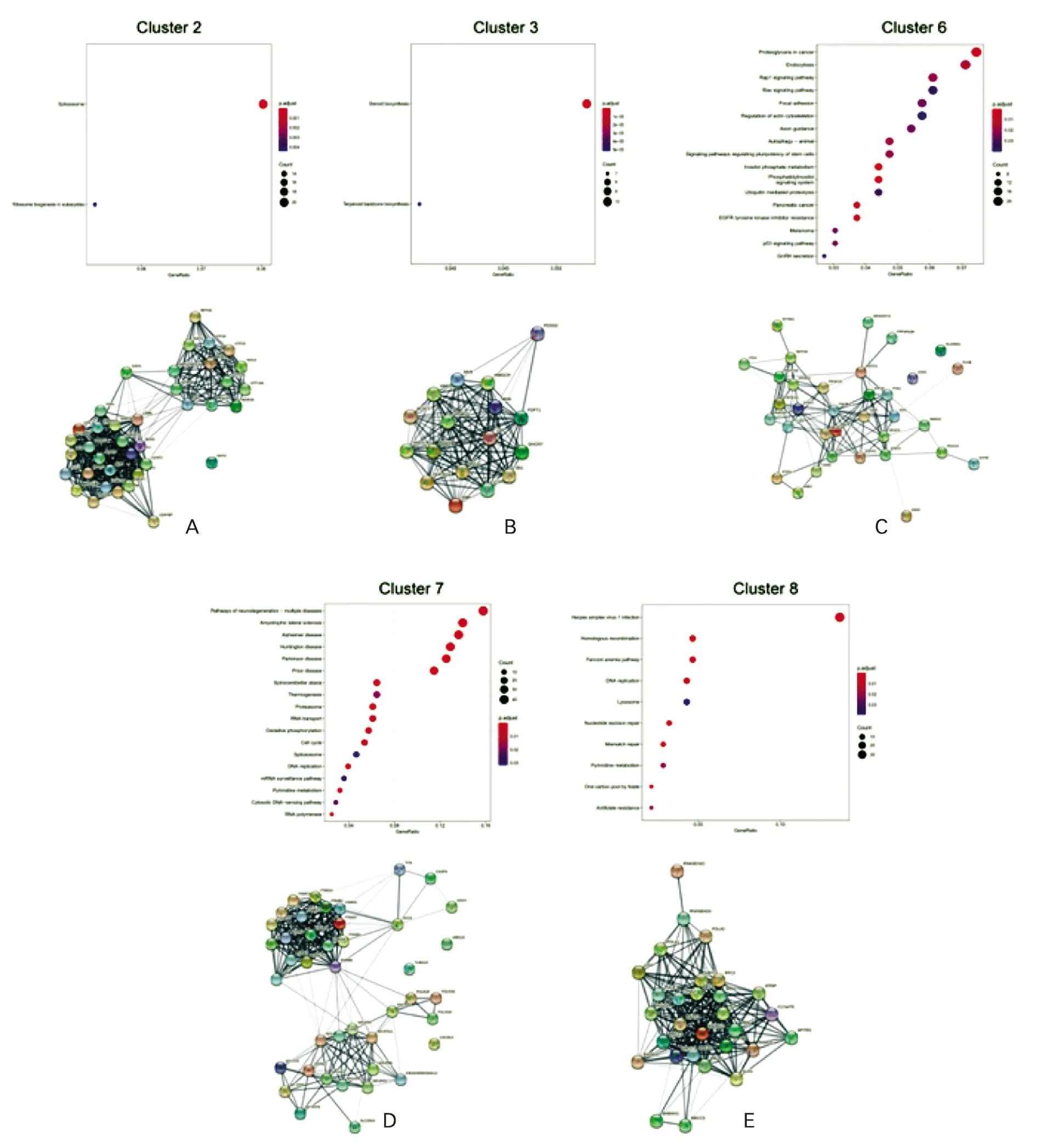
图4 各簇基因功能富集及互作网络A-E示差异基因簇基因KEGG通路富集情况横坐标:基因比率,点的大小和颜色代表富集的显著性,点越大,颜色越深,表示富集程度越高;纵坐标:马法兰处理后各时间点样本差异基因的KEGG富集情况
讨 论
视网膜母细胞瘤是对儿童危害最大的恶性肿瘤之一,多发生于3岁以下婴幼儿,早期干预往往有着较好的治疗效果和预后[10]。目前认为,视网膜母细胞瘤的发生主要与13号常染色体q14 基因组区相关,其中位于该区域的RB1双等位基因失活,被认为是其发生的最主要因素[11,12]。此外,多种基因和通路异常被发现与视网膜母细胞瘤发生相关,包括CDK、E2F1-CIP2A反馈通路[13]、MDM2[14]、MYCN[15]、SKY[16]、PLAC2[17]、PTEN-PI3K/AKT通路等[18]。
化疗是视网膜母细胞瘤的主要保眼治疗方法,其中动脉介入化疗和玻璃体腔化疗是目前提高患儿保眼率的主要治疗方法[19]。马法兰是视网膜母细胞瘤动脉介入化疗和玻璃体腔化疗中的主要化疗药物,也是一种广泛应用临床肿瘤治疗的药物[20],但其具体抗肿瘤作用机制尚不清楚。本研究通过RNA-seq数据结合生物信息学方法,分析了马法兰处理视网膜母细胞瘤细胞不同时长后基因表达的变化情况,初步揭示了马法兰在视网膜母细胞瘤中的治疗机制。
生物信息学分析表明,马法兰影响视网膜母细胞瘤基因表达谱,对各时间段差异表达基因进行聚类分析得到八类基因簇。第二簇基因在马法兰作用初期(4 h)表达量显著降低而后保持持续低表达,其中WDR3为甲状腺癌易感性相关基因[21],UTP14C可以介导破坏TP53引起的凋亡信号,从而引起卵巢癌[22]。在K562白血病和PC3前列腺癌细胞株中,SRPK1敲除或抑制可以降低细胞的增殖、侵袭和迁移[23]。SR因子及SRSF家族可作为乳腺癌以及一些未发现的肿瘤治疗靶点,例如SRSF6多可引起腺泡形态破坏,引起转移增殖[24]。第六簇基因在马法兰作用初期(4 h)表达量显著升高而后保持高表达,其中PSMD2在胃癌中通过抑制DUSP7、WIP1和PTEN,进而诱导ERK、P38和AKT的磷酸化,从而调控细胞增殖[25]。PSMD4在内质网应激诱导的凋亡肝癌细胞。PSMC3在非小细胞肺癌中通过miR-182-5p/NME2信号通路抑制细胞的侵袭和迁移[26]。MD-ND6突变可通过增加ROS促进宫颈癌变[27]。第七簇基因在马法兰作用视网膜母细胞瘤后表达量不断升高,其中PSMA4的变异与肺癌的发病率相关[28]。NDUFA11的调节与肺鳞状细胞癌的发生相关[29]。第八簇基因在马法兰作用视网膜母细胞瘤后表达量不断降低,例如BRCA1和RAD51C是遗传性卵巢癌和乳腺癌的主要基因[30,31]。RAD51在同源重组中起重要作用[32]。RFC2作为RFC家族的一员,已经被报道与结肠癌及各种恶性肿瘤相关,并在增殖、侵袭以及远处转移中起重要作用[33]。第三簇基因于马法兰作用8 h后显著升高,其中NSDHL在胰导管腺癌中的缺失可进一步激活转化生长因子β,从而导致上皮间充质的转化[34]。该簇基因多影响脂类、类固醇类合成。
综上所述,马法兰可能通过抑制DNA复制和同源重组,并促进磷酸肌醇代谢和萜类化合物生物合成等多个方面,达到抑制视网膜母细胞瘤生长并杀伤肿瘤细胞的作用,为临床更加合理选择应用马法兰提供了依据。
