模拟干旱胁迫下褪黑素和表油菜素内酯对沟叶结缕草长期继代培养愈伤组织再生的影响
2022-03-12周认蔡宇林恬逸柴明良
周认,蔡宇,林恬逸,柴明良
(浙江大学农业与生物技术学院,杭州 310058)
干旱是影响植被生长最严重的环境问题之一,全球干旱半干旱地区占总陆地面积的41%,我国243 万km2面积的土地为干旱和半干旱地区。近年来,全球气候的变化,人口、农业用水需求的增长以及淡水储量的降低,加剧了干旱对草业生产的影响[1]。因此,探究植物干旱胁迫的生理生化响应机制、培育耐旱节水型新品种是草坪草性状改良的重要途径。沟叶结缕草[Zoysia matrella(L.)Merr.]是我国南方最重要的草坪草之一,属于耐旱性较强的暖季型草坪草,是应对淡水资源短缺和全球变暖导致夏季干旱频发问题的潜在优质草种,因此,进一步选育耐旱性强的新品种迫在眉睫。但沟叶结缕草几乎无可育的种子,难以进行杂交育种,故主要使用体细胞无性系变异技术对其性状改良,该技术依赖于高效的离体再生体系[2]。沟叶结缕草普遍存在胚性愈伤组织诱导率低、继代生长缓慢以及胚性维持困难等问题[3]。为此,本实验室建立了长期培养的沟叶结缕草愈伤组织体系,通过逐代积累变异增加了获得变异株的可能性,已借助生物技术手段筛选出多种性状优良的植株[4−8]。由此可见,选择继代多年的沟叶结缕草愈伤组织进行抗旱植株选育,具有培养条件可控、变异率高等优势,获得的变异植株是筛选、评价耐旱沟叶结缕草的理想材料。
目前,关于褪黑素(melatonin, MEL)和表油菜素内酯(epibrassinolide,EBL)对植物离体再生培养影响的相关研究结果表明:MEL 能提高滇黄芩(Scutellaria amoena)愈伤组织的生长与再生水平[9],也能促进葡萄(Vitis vinifera)体细胞胚的诱导和生长[10];EBL 显著提升了长期继代的沟叶结缕草愈伤组织的增殖、再生能力[11]。故探究MEL和EBL恢复愈伤组织再生能力的最适浓度,能解决干旱选择压力下长期继代的愈伤组织再生率下降的问题,有利于获得抗性变异株系。
干旱时植物体产生活性氧(reactive oxygen species, ROS),从而引发氧化胁迫,该现象对植物细胞干旱胁迫下的再生过程具有双重作用:一方面抗氧化酶活性提高能促进再生;另一方面发生氧化胁迫的细胞膜通透性发生改变,可导致细胞死亡[12]。聚乙二醇(polyethylene glycol, PEG)作为模拟干旱剂,通过调节基质的渗透压来抑制植物体的水分吸收,创造类似于在实际干旱时完整植物细胞中观察到的水分亏缺条件,且PEG 无毒性,是诱导植物干旱状态理想的渗透调节剂之一[13]。可见,选择适宜浓度的PEG 是筛选获得耐旱植株至关重要的前提。
本研究以MEL和EBL作为外源激素,探究2种激素对干旱胁迫下长期继代培养的沟叶结缕草愈伤组织再生水平的影响,分析再生过程中愈伤组织抗氧化酶的活性变化,为外源MEL 和EBL 对继代多年的沟叶结缕草在逆境胁迫下的生理生化调控机制研究、再生能力的恢复以及耐旱体细胞无性系变异筛选提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为浙江大学观赏植物组织培养实验室离体继代培养17 年的沟叶结缕草胚性愈伤组织体系,继代培养基为Murashige & Skoog(MS)培养基+2.00 mg/L 2,4−二氯苯氧乙酸+1.15 g/L 脯氨酸+40.00 g/L 蔗糖+3.00 g/L 植物凝胶,添加0.10 mg/L苄氨基嘌呤、0.20 mg/L糠基腺嘌呤、0.15 mg/L玉米素和0.10 mg/L噻苯隆,每4周继代培养1次[14]。
1.2 试验方法
1.2.1 模拟干旱胁迫下愈伤组织的再生
在1/2 MS 基本培养基中加入30.00 g/L 蔗糖、3.00 g/L植物凝胶,并分别添加0、5%、10%、15%、20%的PEG−6000(上海阿拉丁生物试剂有限公司),再加入2−吗啉乙磺酸(2−morpholinoethanesulfonic acid,MES)缓冲液,稳定培养基的pH[15]。在预培养3周的愈伤组织中,选取直径约为3 mm,大小相似、长势旺盛的愈伤组织块接种至培养瓶中,每瓶接种9(3×3)块,各浓度梯度设置5个生物学重复。在光照16 h/d,光照强度约为40.00 μmol/(m2·s),温度为(25±2)℃的组织培养室中进行培养。再生6周后,观察并计算愈伤组织的生芽数、再生率及褐化率[11]等。

1.2.2 模拟干旱胁迫下褪黑素、表油菜素内酯对愈伤组织再生的影响
在0~20%PEG−6000的再生培养基中,分别添加不同浓度的MEL 或EBL(表1),MEL、EBL 均购于上海阿拉丁生物试剂有限公司。愈伤组织接种方式、再生培养条件、周期以及再生指标观察记录方法同1.2.1节。
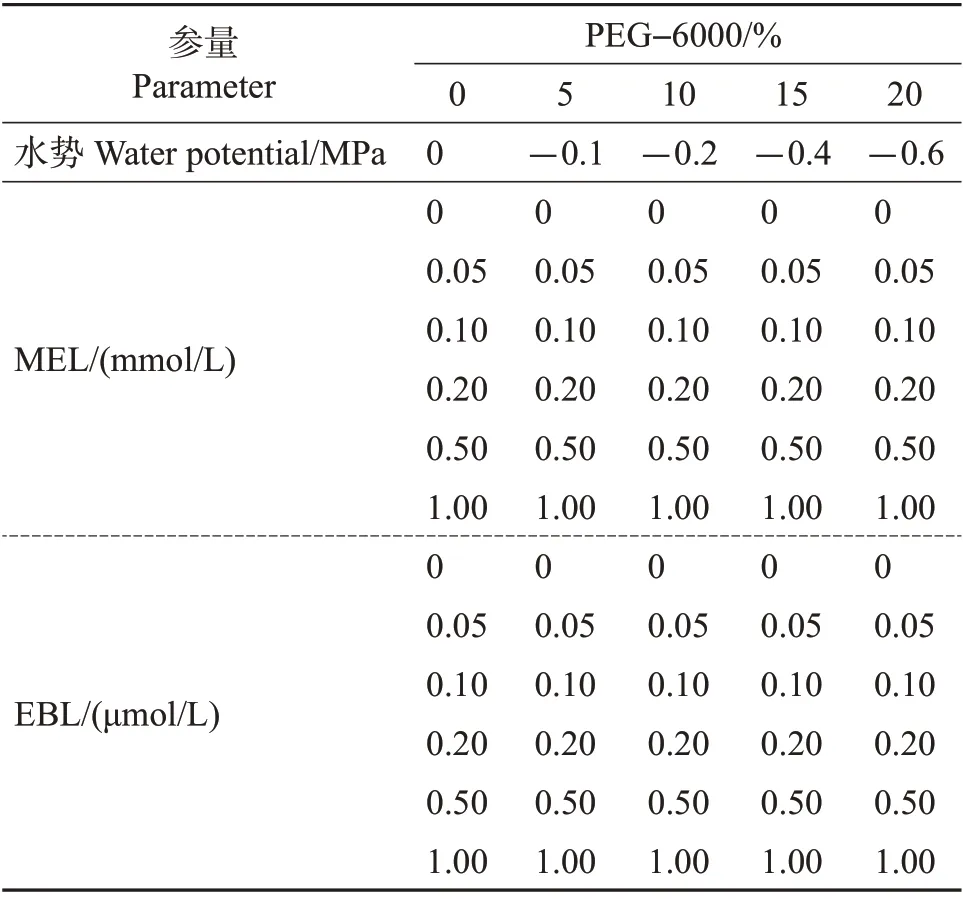
表1 模拟干旱胁迫下沟叶结缕草愈伤组织再生过程中添加的MEL、EBL浓度Table 1 Concentrations of MEL and EBL added during the callus regeneration of Z. matrella under simulated drought stress
1.2.3 模拟干旱胁迫下褪黑素、表油菜素内酯对愈伤组织再生生理的影响
对不同处理的沟叶结缕草愈伤组织再生6周后的植株进行抗氧化酶活性测定,包括过氧化氢酶(catalase, CAT)、超 氧 化 物 歧 化 酶(superoxide dismutase, SOD)、过氧化物酶(peroxidase, POD)的活性和丙二醛(malondialdehyde,MDA)含量。随机称取每个处理下再生培养6周后的植株0.10 g,加入液氮,每个处理取3 个重复,随后加入3 mL 50 mmol/L 磷酸缓冲盐溶液(phosphate buffered saline,PBS),将混合物置于研磨仪中,研磨充分后,在4 ℃条件下以1.2×104r/min 离心20 min,保留上清液。参考LIN 等[16]、LIU 等[17]的方法,用UV−2550 型分光光度计分别测定抗氧化酶CAT、POD、SOD 活性及MDA含量。
1.3 数据统计分析
试验结果均取3 次生物学重复的平均值,使用SPSS 21.0 软件进行方差分析(analysis of variance,ANOVA)以及邓肯多重比较,以P<0.05 表示差异有统计学意义。
2 结果与分析
2.1 模拟干旱胁迫下褪黑素、表油菜素内酯对沟叶结缕草愈伤组织再生的影响分析
由观察得知,再生过程中沟叶结缕草胚性愈伤组织的颜色由浅黄转为深绿,14 d 后表面突起分化形成再生芽,继而长出再生苗,愈伤组织部分则变为黑褐色。在干旱胁迫下,其再生率受到影响,且随着干旱胁迫程度加剧,再生率从17.4%降至0(表2)。当PEG−6000 质量分数达到15%时,大部分愈伤组织褐化死亡,分化芽平均数降至0.6个;当使用20%PEG−6000胁迫时,愈伤组织失去分化能力,再生率为0;而在5%或10% PEG−6000 胁迫下,部分愈伤组织能分化成苗并生根。
愈伤组织的再生水平在不同浓度MEL 或EBL处理下存在差异。在0.20 mmol/L MEL 处理下,愈伤组织的再生率最高,为61.1%,是对照组的3.5倍。当MEL 浓度大于0.50 mmol/L 或EBL 浓度大于0.50 μmol/L 时,愈伤组织丧失再生能力,褐化率显著上升,说明高浓度激素抑制了再生,甚至导致愈伤组织死亡。
干旱胁迫时添加不同浓度MEL或EBL,沟叶结缕草愈伤组织再生率表现出明显差异(表2)。培养6周后,添加适宜浓度的MEL或EBL对愈伤组织再生率具有促进作用,2种激素均缓解了干旱胁迫对愈伤组织再生造成的负面影响。MEL 或EBL 对愈伤组织再生水平的促进效果随着浓度的增加先增大后减小。无干旱胁迫时,0.20 mmol/L MEL使愈伤组织再生率达到最高,褐化率随着MEL 浓度升高而增加。在5% PEG−6000 胁迫下,0.10 mmol/L MEL 和0.10 μmol/L EBL对愈伤组织再生率促进最大,能恢复并提高其再生率分别至37.0%、20.3%,分别为对照的2.3倍和1.2倍,其次是0.05 mmol/L MEL显著提高了愈伤组织的再生水平。在10%PEG−6000 胁迫下,0.20 mmol/L MEL和0.50 μmol/L EBL使愈伤组织再生率高达55.6%和20.8%,除了0.10 mmol/L EBL 处理,其余各浓度的MEL、EBL处理组都在一定程度上恢复了愈伤组织的再生能力。当PEG−6000质量分数 达 到15%时,0.10 mmol/L MEL 和0.05 μmol/L EBL 促进再生的作用最强,再生率分别为对照的10.4倍、6.7倍,0.20 mmol/L MEL和0.10 μmol/L EBL的作用次之。高浓度PEG破坏了愈伤组织胚性细胞的结构,造成了无法逆转的伤害,外源施加MEL 和EBL 不能恢复其再生能力。值得注意的是,0.05 mmol/L MEL对高浓度干旱胁迫下愈伤组织的再生有一定的恢复作用,再生率为44.4%,该部分再生植株可作为耐旱株系进一步进行田间筛选。

表2 模拟干旱胁迫下MEL、EBL对沟叶结缕草愈伤组织再生的影响Table 2 Effects of MEL and EBL on callus regeneration of Z.matrella under simulated drought stress
在5%~15% PEG−6000 胁 迫 下,0.05~0.20 mmol/L MEL或0.05~0.50 μmol/L EBL处理对长期继代的愈伤组织分化水平具有显著的恢复作用,其中,在10%PEG−6000胁迫时加入0.20 mmol/L MEL的条件下,培养基水势较低,愈伤组织细胞渗透吸水能力变差,但仍然保持一定的吸水量,故能保持高再生率。MEL和EBL浓度过高则抑制再生,并导致褐化率显著升高(图1)。

图1 再生6周后模拟干旱胁迫下MEL处理的愈伤组织再生情况Fig.1 Regeneration status of the callus treated with MEL under simulated drought stress after six weeks
2.2 MEL、EBL 对模拟干旱胁迫下沟叶结缕草再生生理指标的影响分析
模拟干旱胁迫下,MEL或EBL对沟叶结缕草愈伤组织再生过程中的保护酶活性和MDA含量有明显影响(P<0.05)(图2),不同浓度MEL 或EBL 对CAT、SOD、POD 活性以及MDA 的作用效果不同。在未获得再生植株的处理组中,对其愈伤组织进行生理测定。无干旱胁迫时,0.20 mmol/L MEL 处理的愈伤组织分化芽总数最多,此时CAT活性显著升高,POD、SOD活性显著降低。当用5%、10%PEG−6000 胁迫时,0.05~0.20 mmol/L MEL 处理的愈伤组织酶活性变化遵循以上规律。在10%PEG−6000胁迫下,当MEL浓度为0.10 mmol/L时,CAT活性最高,为相应对照组的4.8倍;POD活性比相应对照减少70.3%;SOD 活性降低至对照的61.0%。随着MEL 浓度的增加,CAT 活性先升高后降低再升高。由此可知,MEL对沟叶结缕草愈伤组织的再生形态及生理生化机制具有重要的调节作用。

图2 模拟干旱胁迫下MEL、EBL处理的愈伤组织再生过程中生理指标的变化Fig.2 Physiological index differences of the regenerated callus treated with MEL and EBL under simulated drought stress
在不同程度干旱胁迫时,EBL处理的愈伤组织CAT活性先降低再升高,POD、SOD活性则先升高后降低,其中,在10% PEG−6000 胁迫时,0.10 μmol/L EBL处理的CAT活性最低,比同组对照减少31.8%,POD 活性比对照增加39.5%,SOD 活性比对照增加34.0%。在5%、10% PEG−6000 处理时,0.05~0.10 μmol/L EBL 使CAT 活性显著降低,POD、SOD 活性升高,MDA 含量变化无显著差异。MEL 或EBL 的浓度与其对愈伤组织抗氧化酶系统的调节程度呈负相关,抗氧化酶活性呈现的变化趋势与再生形态指标的变化趋势基本一致。综上所述,在5%、10%PEG−6000 胁迫下加入0.05~0.20 mmol/L MEL 或0.05~0.10 μmol/L EBL,可通过激活不同的抗氧化反应途径以减轻干旱胁迫对植物体细胞的伤害,并促进胚性细胞的发生和发育。
3 讨论
采用长期保存的沟叶结缕草愈伤组织进行再生培养,能提高变异植株的获得率,但随着继代时间和次数的增加,愈伤组织的再生率由继代6 年的80%降至继代17 年的17.4%[2],且高浓度PEG 作用下愈伤组织的再生能力显著减弱,以致无法获得足够的理想再生植株。因此,在再生培养基中添加适宜浓度的MEL 或EBL 对耐旱植株的选育十分关键。相关研究表明,培养基中添加低浓度(0.10 μmol/L或1.00 μmol/L)MEL有助于滇黄芩愈伤组织再生,高浓度(100.00 μmol/L)MEL 则抑制再生[9]。本研究中,0.20 mmol/L MEL 显著增强了继代多年的沟叶结缕草愈伤组织的再生水平,但当MEL浓度大于0.50 mmol/L 时则抑制再生,故MEL 能促进长期继代培养的愈伤组织胚性保持和增殖、不定芽分化及生根,提高干旱胁迫下愈伤组织的再生率[18−19],进而为田间耐旱植株的筛选提供丰富材料。
继代多年的沟叶结缕草胚性愈伤组织开始分化时,细胞中出现折叠盾片状胚胎,CAT 和POD 活性降低,SOD活性升高,MDA含量下降,说明植物愈伤组织的再生水平与抗氧化酶活性密切相关[16,20]。测定干旱胁迫时愈伤细胞抗氧化酶活性及MDA含量,发现再生能力强的愈伤组织中至少有一种抗氧化酶活性较高,MDA含量较低。有报道表明,2种生态型的芦苇(Phragmites communisTrin.)在20% PEG−8000 胁迫下胚性悬浮再生培养时,沙丘芦苇的SOD、CAT 活性升高,比沼泽芦苇显示出更强的ROS 清除能力[21]。因此,抗氧化酶活性及MDA 含量指标能在一定程度上反映沟叶结缕草愈伤组织在干旱胁迫下的再生能力。BAYOUMI等使用20%的PEG−6000处理幼苗1周,证实PEG−6000在小麦(Triticum aestivumL.)中产生了化学干旱的作用[22],随后PEG−6000 被用于水稻(Oryza sativaL.)[23]、甘蔗(Saccharum officinarumL.)[24]、番茄(Lycopersicon escolentumMill.)[25]、百里香(Thymus vulgarisL.)[26]等植物的耐旱性筛选研究[27]中,但在草坪草中鲜有报道。本研究中沟叶结缕草愈伤组织经5%、10%PEG−6000处理能产生再生植株,说明沟叶结缕草愈伤组织对干旱胁迫具有适应性。
MEL具有优越的抗氧化性,通过在酶水平或转录水平上调节活性氮、氧类代谢酶来实现植物的硝基氧化稳态,从而有效缓解干旱胁迫的伤害[28−29]。本研究中,适宜浓度的MEL提高了沟叶结缕草愈伤组织离体分化过程中的CAT活性,有利于早期胚胎发育和胚性细胞的分化,这与BIDABADI等的研究结果[30]基本一致;同时降低了POD 活性,而低水平POD活性对细胞分裂、增殖具有促进作用[20]。因此,MEL主要通过提高CAT活性、降低POD活性来减轻干旱胁迫对愈伤组织分化的负面影响。0.02 μmol/L EBL能通过油菜素内酯(brassinolide,BR)途径激活长期继代的沟叶结缕草愈伤组织抗氧化酶的响应,促进其生长和提高其再生能力,在培养基中可作为细胞分裂素的替代物,用于胚性愈伤组织的诱导培养和再生,而浓度大于0.10 μmol/L时则抑制再生[11]。本研究表明,干旱胁迫时较高浓度EBL仍能提高愈伤组织的再生率,促使愈伤组织再生过程中POD、SOD活性显著升高,但其作用效果不如MEL。
试验中,MDA 含量与再生能力呈负相关,作为判断细胞过氧化反应强弱的一个指标,未加MEL、EBL时,细胞内MDA含量升高,当添加适宜浓度的MEL、EBL 后,MDA 含量降低。说明MEL、EBL 通过激活抗氧化酶系统来降低膜脂质过氧化作用,提高抗氧化酶的活性,从而抑制MDA的产生和积累,增强愈伤组织的再生能力。
4 结论
本研究结果表明,0.20 mmol/L MEL 能促进继代培养17年的沟叶结缕草愈伤组织再生,显著提高其CAT 活性。模拟干旱胁迫导致愈伤组织的再生能力下降,在5%或10%PEG−6000 胁迫下,愈伤组织仍能再生;而在高浓度PEG−6000 模拟干旱胁迫下,愈伤组织褐化死亡,丧失分化能力。外源MEL和EBL能显著缓解干旱胁迫对愈伤组织的损伤,恢复其生长和再生的能力,从而获得足够的再生植株。0.20 mmol/L MEL 或0.50 μmol/L EBL 对10%PEG−6000 模拟干旱胁迫下的愈伤组织再生生长具有显著的促进作用,其中0.20 mmol/L MEL的效果最佳。0.05~0.20 mmol/L MEL 或0.05~0.10 μmol/L EBL 可调控愈伤组织再生的生理生化过程并提高胚性细胞分化水平,进而提高植株再生率。总之,本研究探索了逆境胁迫下提高长期继代培养的愈伤组织再生能力的方法,为沟叶结缕草耐旱体细胞无性系变异育种提供了新思路。
