蓝莓花青素生物合成相关基因表达与分子进化分析
2022-02-16郑嘉伟王兰娇李大婧张晓晓黄午阳
郑嘉伟,王兰娇,李大婧,柴 智,张晓晓,黄午阳,*
(1.江苏大学食品与生物工程学院,江苏 镇江 212013;2.江苏省农业科学院农产品加工研究所,江苏 南京 210014)
花青素是自然界一类广泛存在于植物中的水溶性天然色素,存在于很多食品中。花青素对人类健康有积极的影响,具有抗氧化、抗癌、保护视网膜、降低血脂、抗衰老和改善肠道健康等重要功效[1]。蓝莓含有丰富的花青素。因此有关蓝莓花青素的研究也不断深入,其中包括一些有关基因及其调控方面的研究。植物花青素生物合成途径中的结构基因可以达到协同调控花青素生成的作用,在红葡萄中与花青素合成有关的查耳酮合酶(chalcone synthase,CHS)、尿苷二磷酸-葡萄糖-类黄酮-3-葡萄糖基转移酶(uridine diphosphate-glucoseflavonoid-3-glucosyl transferase,UFGT)等结构基因表达量均高于白葡萄[2]。CHS是花青素合成途径中的第一位关键酶,能够催化生成柚皮素查耳酮,并在查耳酮异构酶(chalcone isomerase,CHI)、黄烷酮-3-羟化酶(flavanone 3-hydroxylase,F3H)、二氢黄酮醇-4-还原酶(dihydroflavonol-4-reductase,DFR)和花青素合成酶(anthocyanidin synthase,ANS)的连续作用下形成花青素[3-5],而花青素还原酶(anthocyanidin reductase,ANR)是合成原花青素的关键酶,在形成花青素后经ANR的催化可生成原花青素,而不通过UFGT转化较为稳定花色苷[6-7],通过不同途径分支消耗相同底物也是花青素合成的一种调控机制,可以说ANR基因在花青素的生物合成过程中起负调控作用,有研究表明,拟南芥在缺少ANR基因的情况下能够促使花青素合成,使拟南芥的花青素含量提高[8]。当花青素合成遭到阻碍时,可以发现CHS、ANS和UFGT基因的表达水平大幅度降低[9]。植物中花青素含量受多方面因素影响。一方面,紫外线等外部环境会影响花青素生物合成基因的表达水平,紫外线可以诱导花青素积累在植物组织通过花青素生物合成基因激活,显著提高花青素化合物的水平[10]。另一方面花青素合成需要两类基因,一类是编码直接参与花青素和其他类黄酮形成的酶的结构基因,另一类是控制结构基因转录的调控基因[11]。花青素的合成途径属次生代谢产物黄酮类的分支,尽管不同物种在花青素的构成成分和积累模式上有较大区别,但其合成时经历的主要反应基本一致[12]。其合成路径主要分为3个阶段,第1、2阶段为类黄酮代谢的反应,第3阶段为各个花青素的形成阶段。花青素的形成主要依靠CHS、CHI、F3H、DFR、ANS、ANR等关键酶(图1)。目前相关学者已从金鱼草(Antirrhinum majus)、玉米(Zea mays)、杨梅(Myrica rubra)等众多植物中得到了花青素合成所需酶类的基因[13-15]。
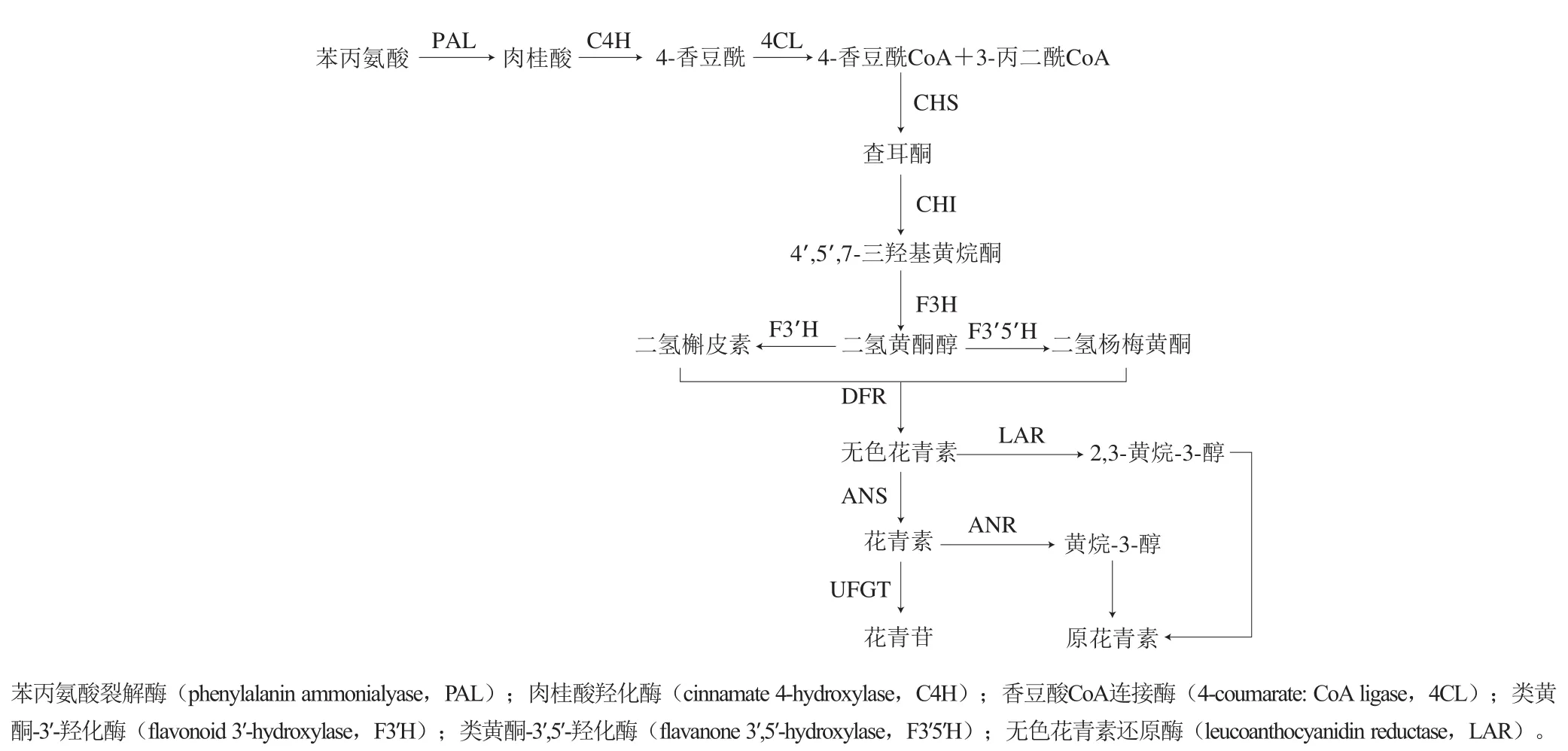
图1 花青素的生物合成途径Fig. 1 Anthocyanin biosynthesis pathway
转录因子又被称为反式作用因子,它是真核生物当中所含的一种DNA结合蛋白,作用于启动子区并与顺式元件结合,对基因的表达具有调节作用[16]。转录因子在高等植物各组织器官中普遍存在,广泛参与调控植物的生长发育,其作用方式多种多样,但大多是与特定靶基因上游的特异性核苷酸序列相结合,调控下游的基因转录的一种蛋白。转录因子特异性地结合靶基因的启动子、增强子等调控转录的区域,抑制或者促进转录,进而实现对具体代谢途径或生命活动的调控[17]。在植物花青素合成中可能由一个或多个转录因子基因调控结构基因协同表达,研究表明花青素在合成过程中,主要由髓细胞组织增生病毒癌基因同源物(v-myb avian myeloblastosis viral oncogene homolog,MYB)、bHLH、WD40转录因子及其MBW复合物分别或共同参与调控代谢途径[18],MYB基因已被证实是调控花青素生物合成的重要转录因子之一[19]。MYB基因家族很大,在植物的不同代谢途径中能发挥不同的用途,它能够参与植物表皮细胞的分化、辅助气孔细胞运动、调控黄酮类化合物的生物合成以及抵抗真菌感染等作用。在参与花青素生物代谢过程时,MYB转录因子能够通过调控花青素合成相关酶基因促进花青素的生成,如葡萄中VvMYBPA1能够调控CHI基因的表达,VvMYBBA2能够促进UFGT的表达[20]。同时在一些植物研究中发现其具有负调控作用[21],有研究证实,在拟南芥中存在的AtMYB4转录因子能够通过抑制C4H基因的合成达到负向调控植物对UV-B耐受性的作用[22]。Yao Gaifang等[23]发现该基因可调控花青素合成,且抑制其表达可促使红皮梨中花青素生成。R2R3MYB调节剂通常是决定花青素积累的关键因素[24-25]。此外,R2R3MYB蛋白的c端序列很长,它专门控制花青素生物合成、偶联和转运花青素到液泡中[26]。花青素生物合成是一个复杂的体系,具体调控方式及作用机理还有待进一步研究。本研究通过分析不同品种蓝莓花青素生物合成结构基因和转录因子基因表达量,进而探究其与花青素含量的相关性及系统发育,探索花青素生物合成的调控机制。
1 材料与方法
1.1 材料与试剂
8个品种蓝莓样品由江苏省南京市中山植物园中国科学院植物研究所提供,产自浙江诸暨。
蓝莓花青素标准品Mv-3-O-gal、Mv-3-O-glc 美国Sigma-Aldrich公司;Dp-3-O-glc、Cy-3-O-gal、Cy-3-O-ara、Pt-3-O-gal、Pt-3-O-glc 北京索莱宝科技有限公司;Dp-3-O-ara、Pt-3-O-ara 上海吉至生化有限公司;Dp-3-O-gal、Pn-3-O-gal、Cy-3-O-glc 法国Extrasynthese公司;Mv-3-O-ara 上海甄准生物科技有限公司;乙腈(色谱纯)美国Tedia试剂有限公司;磷酸(色谱纯) 阿拉丁试剂(上海)有限公司;RNAprep Pure多糖多酚植物总RNA提取试剂盒(DP441) 天根生化科技(北京)有限公司;cDNA合成试剂盒TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix、TransStart Top Green qPCR SuperMix荧光定量聚合酶链式反应(polymerase chain reaction,PCR)试剂盒 北京全式金生物公司。
1.2 仪器与设备
AG梯度PCR仪 德国艾本德股份公司;Applied Biosystems Q3实时定量PCR仪 美国赛默飞世尔科技公司;NanoDrop One超微量紫外分光光度计 基因有限公司;Tg16-WS台式高速离心机 长沙湘仪离心机仪器有限公司;Tanon 2500全自动数码凝胶成像系统 上海天能科技有限公司;DYY-2稳压稳流电泳仪 上海越磁电子科技有限公司;Thermo Forma 900超低温冰箱 上海甄明科学仪器有限公司。
1.3 方法
1.3.1 花青素含量的测定
蓝莓花青素按前期报道的方法[27]提取后,提取物用0.22 μm聚偏二氟乙烯膜过滤,采用高效液相色谱法测定各品种蓝莓果花青素提取物中各花青素含量,总花青素含量为各花青素含量的总和,包括酰基化的花青素。1200高效液相色谱仪配置G1311A二元泵和G1315D二极管阵列检测器,并采用C18色谱柱进行分离。以体积分数1.0%的磷酸缓冲液为流动相A,以100%乙腈为流动相B。流速0.6 mL/min,运行柱温25 ℃,波长520 nm。
1.3.2 蓝莓总RNA提取
将存于-80 ℃的样品放入研钵中,与液氮一起迅速研磨成粉末;向475 μL裂解液中加入25 μLβ-巯基乙醇,随后加入100 mg样品粉末并立即旋涡剧烈振荡混匀,之后参照RNAprep Pure多糖多酚植物总RNA提取试剂盒说明书进行蓝莓总RNA提取,以OD260nm/OD280nm比值检验RNA纯度,采用琼脂糖凝胶电泳检测RNA完整性。
1.3.3 cDNA合成
将提取出的完整RNA稀释后,用cDNA试剂盒合成反转录第一链cDNA,加入1 000 ng总RNA。
cDNA合成反应体系为2 μL的5×PrimeScriptRTMaster Mix(Perfect Real Time)、2 μL总RNA、6 μL RNase-Free ddH2O。反转录条件为:反转录(37 ℃反应15 min),反转录酶失活(85 ℃反应5 s),4 ℃冷却。于冰上冷却后,分装并置-20 ℃冰箱存储备用。
1.3.4 实时PCR(real-time PCR)测定
根据花青素生物合成通路选择主要结构基因VcCHS、VcCHI、VcF3H、VcDFR、VcANS、VcANR和转录因子基因VcMYBPA1进行测定。内参基因准确平稳表达是real-time PCR结果的重要决定因素,一般选择表达水平相对稳定的参考基因,如Actin、NCED1、GAPDH、EF-1α、UBQ、SAND等,本研究经real-time PCR实验比对后,选择已知的UBQ作为内参基因,所用引物见表1[28-29]。

表1 花青素合成相关基因引物序列Table 1 Primer sequences used for amplification of target genes involved in anthocyanin synthesis
以cDNA为模板,根据试剂盒说明书进行操作,其中cDNA质量浓度为50 ng/μL,引物浓度为10 μmol/L。PCR程序为:95 ℃预变性2 min,95 ℃变性30 s,52 ℃退火30 s,72 ℃延伸30 s,循环次数为40。反应结束后得到荧光值变化曲线和溶解曲线,以目的基因与内参基因Ct值之差计算基因的相对表达差异,参考2-ΔΔCt分析法[30]。
1.3.5 RNA浓度的计算
超微量紫外分光光度计用RNase-Free ddH2O调零后,取2 μL用RNase-Free ddH2O稀释的蓝莓RNA提取物测定其OD260nm值,并按照下式计算RNA质量浓度:
质量浓度/(ng/μL)=OD260nm×稀释倍数×40
1.3.6 分子系统进化分析与数据处理
2 结果与分析
2.1 不同品种蓝莓花青素生物合成相关基因的表达
2.1.1 相关结构基因相对表达量分析
选取8 种蓝莓果实进行总花青素含量测定,如图2所示。可以看出园蓝、蓝美1号、绿宝石总花青素含量处于较高水平,而海岸、灿烂以及奥尼尔处于较低水平,莱格西和比洛克西处于居中水平。
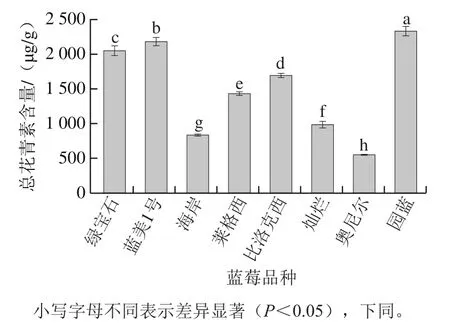
图2 8个品种蓝莓果实总花青素含量Fig. 2 Total anthocyanin contents of eight blueberry varieties
由表2可以看出,总花青素含量最高的为兔眼蓝莓园蓝,其总花青素含量高达(2 331.48±67.33)μg/g;其次为南高丛蓝莓蓝美1号和北高丛蓝莓绿宝石,含量分别为(2 184.07±59.36) μg/g和(2 055.56±61.61) μg/g;而南高丛蓝莓奥尼尔的总花青素含量最低,仅为(554.56±2.83)μg/g,与园蓝相差4 倍多。

表2 8 种蓝莓果实信息及总花青素含量Table 2 Fruit traits and total anthocyanin contents of eight blueberry varieties
对6个蓝莓花青素合成途径中重要结构基因进行real-time PCR检测后,使用相对定量的方式分析其表达量,如图3所示,每个基因均能够在蓝莓中正常表达,但相同基因在不同品种蓝莓的表达量上存在明显差异。VcCHS、VcCHI、VcF3H、VcDFR以及VcANS的表达量在不同品种间的差距较为接近,例如这5个结构基因均在奥尼尔中呈现出最高表达量,灿烂的表达量较高,而蓝美1号的表达量均呈现最低状态,其余品种之间的5个结构基因表达量存在较小浮动,但整体趋势相似;除奥尼尔外,其余品种的VcCHS表达量均较为接近;比洛克西、灿烂和海岸的VcCHI表达量相对较高;莱格西、园蓝和灿烂的VcF3H表达量相对较高;园蓝和灿烂的VcDFR表达量相对较高;比洛克西、灿烂和海岸的VcANS表达量相对较高。而VcANR在不同品种蓝莓间的表达量与其他5 种基因差异较大,其表达量最低的品种为绿宝石,表达量最高的品种为灿烂,莱格西、比洛克西、奥尼尔、蓝美1号的表达量也相对偏高。
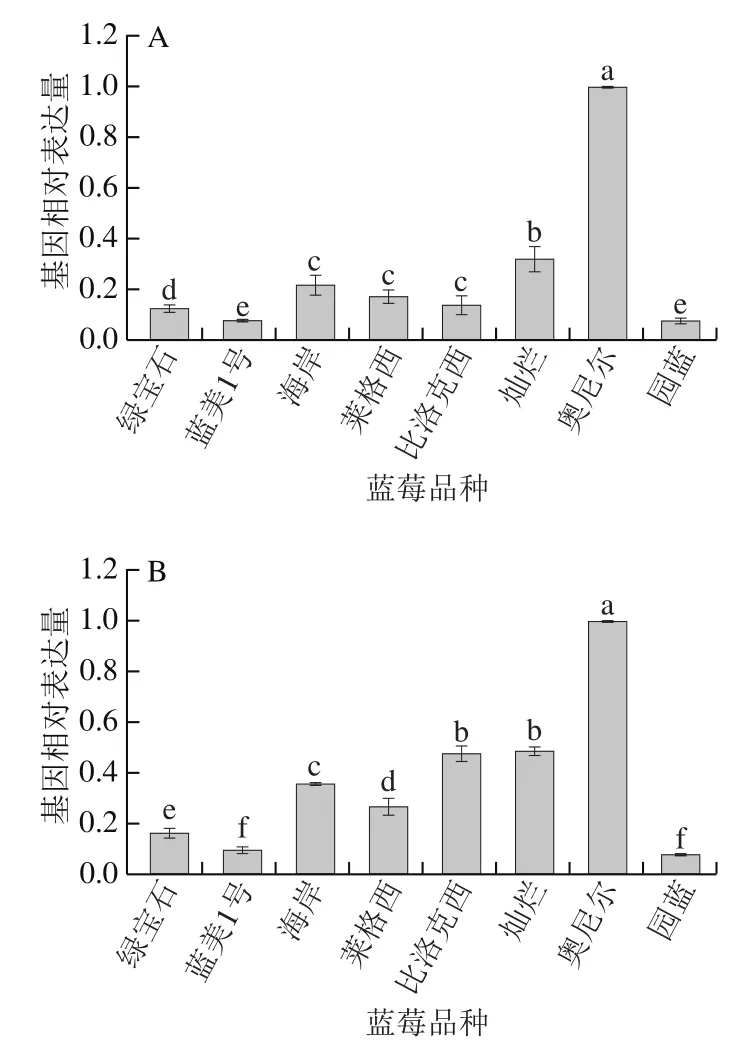
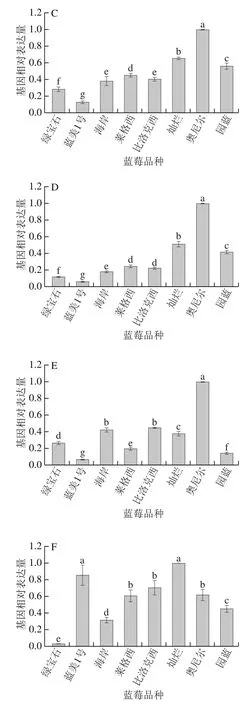
图3 不同品种蓝莓花青素合成各相关结构基因VcCHS(A)、VcCHI(B)、VcF3H(C)、VcDFR(D)、VcANS(E)、VcANR(F)相对表达量Fig. 3 Relative expression levels of VcCHS (A), VcCHI (B), VcF3H (C),VcDFR (D), VcANS (E), and VcANR (F) involved in the biosynthesis of anthocyanins in different blueberry varieties
绿宝石的总花青素含量高,虽然其VcCHS、VcCHI、VcF3H、VcDFR和VcANS基因的表达量均偏低,但VcANR在不同品种蓝莓间的表达量最低,推测与其他品种的蓝莓相比,这5 种基因在绿宝石中主要贡献产生花青素,而其他的类黄酮物质如查耳酮、黄酮醇、黄烷醇较少,且由于负调控基因VcANR极低,ANR催化花青素生成原花青素分支途径被抑制,花青素总含量依旧较高;灿烂的VcCHS、VcCHI、VcF3H、VcDFR和VcANS基因表达量均较为领先,基本处于所有品种蓝莓中的第2位,但其总花青素含量却仅为一般水平,推测是由于其VcANR基因表达量最高,而ANR基因对花青素合成具有负调控作用,其表达量越高,对花青素合成的影响越大,导致花青素向原花青素的方向转化,致使灿烂的总花青素含量减少;奥尼尔的VcCHS、VcCHI、VcF3H、VcDFR和VcANS基因表达量均呈现最高状态,其VcANR基因表达量居于中等位置,但其总花青素含量在8 种蓝莓中最低,这可能是VcCHS、VcCHI、VcF3H、VcDFR主要贡献生成其他类黄酮物质,而且由于其含有一些VcANR基因使其花青素含量降低,但其VcANR基因表达量不足以使其花青素含量降低至此,可能还存在其他基因调控或影响因素导致其花青素含量大幅度减小。同样,蓝美1号VcCHS、VcCHI、VcF3H、VcDFR和VcANS基因表达量均呈现最低状态,而VcANR基因表达量最高,但总花青素含量在8 种蓝莓中偏高,也可能是其他基因(如UFGT等)调控。
整体来看,花青素合成途径的关键酶基因都对最终花青素的生成量起到一定作用,其中VcCHS、VcCHI、VcF3H、VcDFR和VcANS基因负责催化生成花青素,对花青素合成起到明显的正调控作用;而VcANR基因对花青素的合成有负调控作用,能够将产生的花青素催化形成原花青素,最终使蓝莓花青素含量降低。然而单凭这些基因的作用并不能解释所有蓝莓花青素含量与相关基因表达量的关系,仍需从多种角度做进一步研究。
从不同品种蓝莓中各基因表达量对比可以看出,莱格西、比洛克西、蓝美1号和灿烂中VcANR基因表达量最高,证明该基因在这些品种蓝莓花青素合成中积极参与,而在奥尼尔、绿宝石中VcANR基因表达量最低,说明该基因在这两种蓝莓中的作用不十分明显;园蓝和绿宝石中VcF3H基因以及海岸中VcANS基因在其对应品种蓝莓花青素的合成过程中参与性最强;莱格西、比洛克西、园蓝和灿烂的VcCHS基因以及蓝美1号和海岸的VcDFR基因在其对应品种蓝莓花青素合成中的作用较弱。
整体来看,VcANR和VcF3H基因在蓝莓花青素合成过程中基本能够呈现出表达量较高的态势,而VcCHS、VcANS和VcDFR基因通常表达量较低。可见不同品种的蓝莓中各基因的表达情况及参与程度存在差异,使得不同品种的蓝莓最终表现出不同的花青素含量。
2.1.2 相关调控基因相对表达量分析
本实验部分品种蓝莓的MYB表达量极低,也可能是由于不同蓝莓植株间存在遗传差异性或果实空间分布对环境信号的敏感度不同,一般来说,果实表面的光照分布也可能是影响MYB表达水平不明显的因素,已有研究人员在苹果中分离出调控果皮花青素合成的MdMYB1转录因子,并发现光照诱导对该基因表达量的影响很大[31]。
图4为不同品种蓝莓VcMYBPA1转录因子基因的相对表达量,其在所有品种中均存在表达,但品种间存在较大差异。可以看出奥尼尔的VcMYBPA1基因表达量最高,其次为灿烂,约为奥尼尔的1/2;莱格西、比洛克西的表达量为中等水平;蓝美1号、园蓝和海岸的基因表达量较低,而绿宝石的VcMYBPA1基因表达量最低。蓝莓VcMYBPA1基因表达量的整体趋势与VcCHS基因表达量最为相似,可能在更大程度上通过调控VcCHS基因影响花青素的合成。
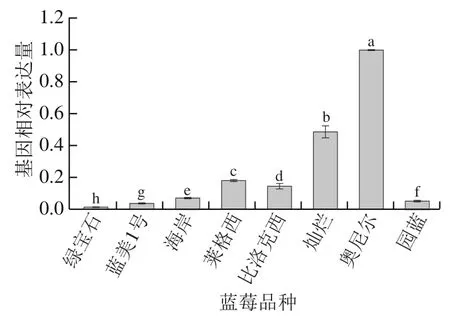
图4 不同品种蓝莓MYB调控基因相对表达量Fig. 4 Relative expression levels of MYB-regulated genes in different blueberry varieties
将图4与图2对比来看,VcMYBPA1基因表达量最高的奥尼尔,其花青素含量最低,VcMYBPA1基因表达量较高的灿烂,其花青素含量也仅为中等水平,造成这种现象可能是VcMYBPA1转录因子反馈抑制,对其产生负调控作用,使得花青素含量降低;VcMYBPA1基因表达量处于中间位置的莱格西、比洛克西,其花青素含量无明显变化趋势;而VcMYBPA1基因表达量较低的蓝美1号、园蓝、绿宝石和海岸的花青素含量却相对偏高,尤其是园蓝、蓝美1号和绿宝石的花青素含量明显高于其他品种,居前3位,可能是少量的VcMYBPA1对其产生正调控作用,促进花青素的生成。以此推测MYB转录因子能够对花青素的生物合成产生促进作用还是抑制或限制花青素的合成,与MYB基因表达量有关,当MYB基因表达量过高时,其可能发挥负调控作用影响花青素的合成,使蓝莓花青素含量降低;当MYB基因表达量处于中等水平时,其对花青素合成的调控作用不十分明显;当MYB基因表达量较低时,其能够发挥正调控作用促进花青素的合成,从而生成更多的蓝莓花青素;这之间可能存在一定的平衡点,当MYB基因表达量超过平衡点时则发挥负调控作用,低于平衡点则有益于花青素的合成。
2.2 不同品种蓝莓花青素生物合成相关基因分子进化与系统发育分析
对上述real-time PCR产物进行测序分析后,得到其反向互补序列,上传至GenBank,并利用BLAST工具将其与数据库中的同种基因进行比对,选择一致性较高的基因采用NJ法构建分子进化树,上传基因和比对基因及其序列号如表3、4所示,根据其置信度辨别样品蓝莓基因与数据库中基因的亲缘关系。
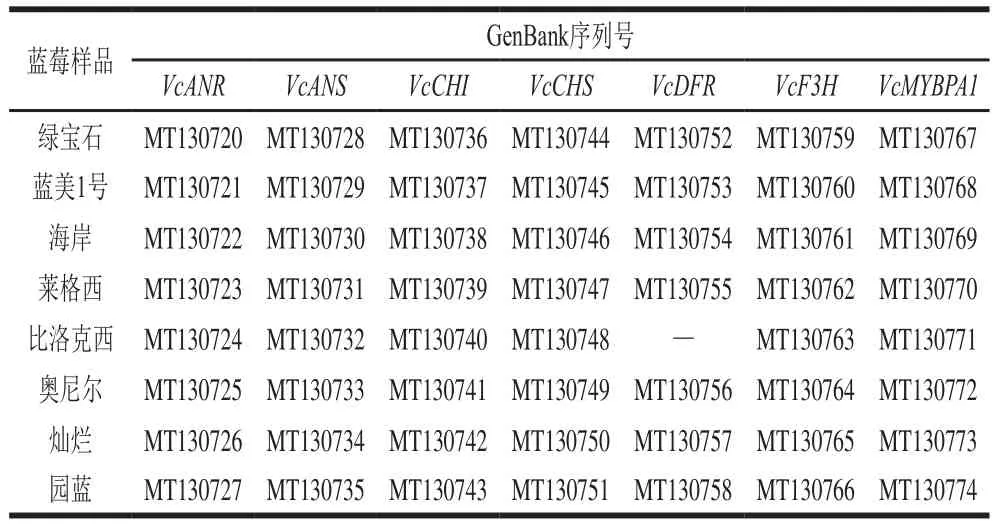
表3 检测蓝莓样品的基因序列号Table 3 GeneBank accession numbers of anthocyanin biosynthesisrelated genes in blueberry samples

表4 参比基因及其序列号Table 4 GeneBank accession numbers of reference genes
样品蓝莓为越橘属植物,GenBank样品名称包括莱格西、比洛克西、奥尼尔、蓝美1号、园蓝、灿烂、绿宝石和海岸。如图5系统发育树和序列比对结果显示,越橘属(Vacciniumspp.)和油茶属(Camellia)的ANR基因功能相近,而与其他物种亲缘关系较远。根据图6也可知二者的蛋白结构域也较为相似,因此越橘属的ANR基因可能是由油茶属进化而来。ANR基因在越橘属中保守性较高(93.38%),在相同谱系的不同种中均存在较为一致的保守序列。

图5 ANR基因的系统发育树Fig. 5 Phylogenetic tree of ANR gene
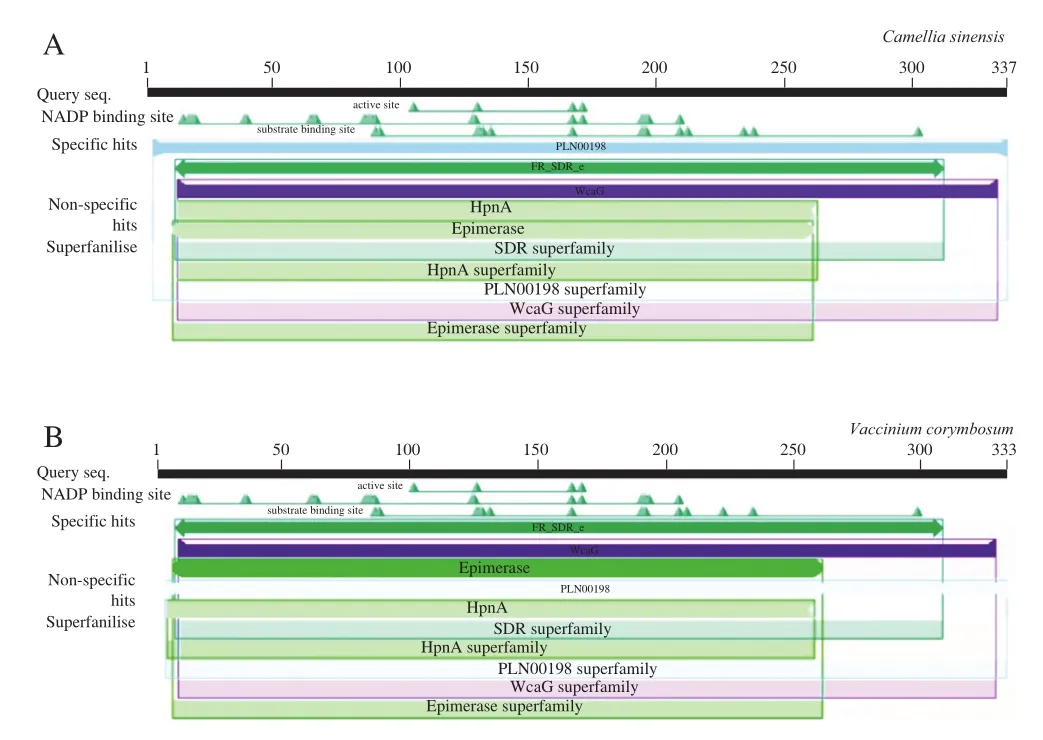
图6 越橘属(A)和油茶属(B)ANR蛋白结构域Fig. 6 ANR protein domains of Vaccinium (A) and Camellia (B)
ANS(97.4%)、CHI(94.83%)、CHS(98.72%)、DFR(94.48%)、F3H(99.45%)在越橘属中保守性均很高,但与其他物种之间存在较大差异,表明这些基因在物种分化后可能独立进化,并在不同物种中趋于保守。植物转录因子MYB是近年来发现的一类与调控植物生长发育、生理代谢、细胞的形态和模式建成等生理过程有关的一类转录因子。结果显示,本研究所测序的MYB蛋白序列属于I类MYB基因家族成员1R-MYB/MYB-related[32],越橘属与油茶属的MYB基因具有很近的亲缘关系,该基因在越橘属、油茶属及栎属(Quercus)中均属于同一个基因家族成员,可能由共同的祖先分化而来,并发挥相似功能。
3 结 论
本研究对蓝莓花青素合成途径中的关键结构基因VcCHS、VcCHI、VcF3H、VcDFR、VcANS、VcANR和转录因子基因VcMYBPA1进行real-time PCR测定得到其相对表达量。研究发现,以上所有基因在蓝莓中均有表达,但不同品种的蓝莓间存在较大差异,其中VcCHS、VcCHI、VcF3H、VcDFR和VcANS基因对花青素的合成有调控作用,能够促进花青素的生成;但VcANR基因对花青素的合成有负调控作用,可使蓝莓花青素含量降低;VcANR和VcF3H基因在蓝莓花青素合成过程中普遍较为活跃,而VcCHS、VcANS和VcDFR基因则相对保守;MYB转录因子对花青素合成的调控作用相对复杂,与MYB基因的表达量有关,当MYB基因表达量过高时,可能发挥负调控作用降低蓝莓花青素含量;当MYB基因表达量处于中等水平时,其对花青素合成的调控作用不显著;当MYB基因表达量较低时,能够促进花青素的合成。花青素的合成还与其他调控因子及影响因素有关,具体调控方式及作用机理还有待进一步研究。
此外,本研究系统测定不同蓝莓品种花青素合成相关基因的序列并进行分子进化和系统发育分析。在蓝莓花青素合成相关基因的进化过程中,其ANR基因与油茶属ANR基因功能相似,并且二者的蛋白结构域也较相似,推测越橘属植物与油茶属是近源关系;蓝莓MYB基因与油茶属MYB基因的亲缘关系也很近,且该基因在越橘属、油茶属及栎属中为同一基因家族成员,可能由共同的祖先分化而来;ANS、CHI、CHS、DFR和F3H基因在越橘属中的保守性很高,与其他物种间存在较大差异,可见这些基因在物种分化后可能会发生独立进化。蓝莓花青素合成相关基因的系统发育分析,有助于了解基因的进化以及生物系统发生的内在规律,指导优质蓝莓品种的选育。总之,蓝莓花青素的生物合成受类别、品种、种植地区、基因调控等多方面因素的影响,本研究探索不同品种蓝莓花青素生物合成相关基因表达量和分子进化,为开发优质蓝莓品种花青素类功能性食品提供理论依据。
