同伴互动的认知神经机制研究分析与未来展望
2022-02-11王宇陈晨杨韵莹郑娅峰董艳
王宇 陈晨 杨韵莹 郑娅峰 董艳
[摘 要] 同伴互动是人类获取知识的重要途径,揭示其内在学习机制与规律对优化学习策略、提升学习效果具有积极意义。认知神经科学为大脑交互研究提供了基础,打破了以往无法解释互动学习模式的认知黑箱,为多维视角探究同伴互动的内在机制提供了跨学科研究的新路径。研究通过探寻同伴互动的认知科学神经基础与技术应用,揭示出现有研究聚焦于发现或验证单一互动行为的神经指标、复杂互动行为背后人际脑同步指标及其影响因素。已有成果为聚焦以学生为中心的教学实践提供融合生理、心理和行为三者模态数据的有效根据,为教师优化学生协作学习的策略提供科学、精准的依据。最后,作者从数据融合、情境聚焦、设备研发、方法整合等方面对未来研究进行了展望。
[关键词] 同伴互动; 认知神经机制; 神经影像技术; 人际脑交互; 协作学习
[中图分类号] G434 [文献标志码] A
[作者简介] 王宇(1992—),女,新疆乌鲁木齐人。博士研究生,主要从事STEM教育、项目式学习研究。E-mail:fergie_w@163.com。董艳为通讯作者,E-mail:98086@bnu.edu.cn。
一、引 言
同伴互动是人类获取知识的重要途径之一。同伴互动促进合作学习或协作学习,成为认知发展和知识习得的重要策略[1]。积极有效的同伴互动可促进学生优势互补、提升合作学习效果,消极、低效的同伴互动会产生低效合作,甚至产生小组矛盾[2]。同伴互动发生机制,有助于设计有效的同伴写作教学策略。传统教育学领域的互动研究主要采用行为分析、问卷调查、访谈观察等,其结果难以反映复杂环境下学习者认知过程和学习状态,亟须发掘、整合新的研究视角和技术方法对其进行表征。作为将神经科学与认知心理学相结合的跨学科研究领域,认知神经科学利用神经影像技术,将互动行为、大脑神经活动机制与影响因素三者联系起来,较为完整地描述互动过程,为教育实践提供充实的理论基础和科学依据。
本研究通过梳理同伴互动的认知神经机制相关研究,从互动中的个体和群体层面阐述主要研究观点,并从单一互动行为和复杂互动行为两个视角总结当下研究的焦点及其主要发现,并结合当前我国教育发展的实际情况,对教育认知神经科学视角下的同伴互动研究的发展前景进行展望,以期促进当前跨学科教育研究的发展。
二、同伴互动神经机制研究的发展进程
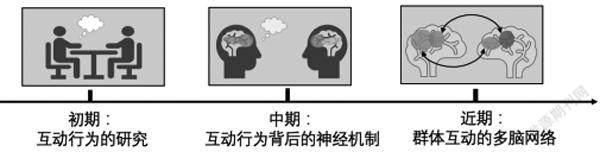
本研究基于同伴互动研究从孕育、萌芽再到迅速发展的过程梳理,发现社会学及教育学领域的互動研究大体经历了“互动行为的研究—互动行为背后的神经机制—群体互动的多脑网络”三大发展阶段,如图1所示。初期互动行为的研究属于认知心理学范畴。随后,神经科学进一步探索了互动行为背后的神经机制,为认知过程增加了生理学证据。近年来,神经影像科学技术的发展促使社会神经科学领域的研究不仅实现了同时获取多人互动的“行为—大脑”数据,还能通过构建多脑网络进一步揭示同伴互动背后的大脑神经活动机制及其影响因素。
(一)同伴互动行为研究
早期研究主要聚焦行为层面,是对互动质量、影响因素、结构关系最为直观的评价和解释方法。行为层面的研究主要基于简单的互动任务,如言语交流、目光交流和合作游戏等。协作学习的核心是互动行为的发生及其产生的学习效果,因此,这类研究主要关注互动行为和互动效果。互动行为以学习者个体的口头语言、肢体动作或面部表情等为基础,从而体现同伴互动过程中外显交互行为的相互关联,其指标主要包括次数、时间和频率等;互动效果则以任务完成的时长、正确率、作品质量或能力高低为表现载体,其指标主要包括测试分数、能力高低、产品质量等。在同伴互动过程中,学习者为从那里获得技能和知识,就要协调彼此的行为——促进彼此行为上的人际行为同步。这种行为层面上的人际同步是一种沟通和相互理解的手段,不仅能够促进群体认知过程的协调,还有助于增强团队凝聚力。多学科研究范式的整合为教育情境中的相关研究提供实验设计、数据采集和分析等支持。
(二)个体互动行为背后的神经机制
行为层面的研究能发现互动特征和影响因素,但无法归因。近十多年来,人们开始好奇互动过程中行为发生和观点产生是从大脑哪个区域生成的?人际行为同步背后的神经活动是怎样的,又如何控制的?为打开这一认知黑箱,研究者开始运用神经科学的手段探索个体与外界发生互动时大脑内部神经活动的特性。由于初探时期的技术水平有限,加之社会互动的复杂性,研究者大多孤立考察单个被试执行互动行为时的神经活动。
该层面的研究是为了揭示大脑内的神经活动与行为之间的关联性,即从个体水平上探究有哪些功能脑区参与了互动过程。首先,要确定神经影像实验中的自变量和因变量,自变量是个体所发出的不同行为或接收到的刺激调控(语言、动作、创造、搭建等),因变量为个体大脑神经响应。其次,针对研究问题,根据先前的理论选择兴趣脑区、设计互动实验任务,并确定数据收集、分析方法。在获得行为和大脑神经活动的数据后,可计算得出兴趣脑区的激活状态以及功能连接(两个脑区信号在时间上存在相关性),进而建立互动行为与神经活动之间的关系,也能够为探索同伴互动相关认知功能的神经基础提供证据。
(三)群体互动的多脑网络
个体水平的经典实验研究范式帮助研究者发掘社交互动学习相关的神经标记,但对于多个个体而言,有关大脑活动间动态联系的宝贵信息总是会被忽略。认知科学家通过神经影像实验研究探索不同社会情境下的多个个体间大脑活动的动态联系特征,并发现互动行为背后的神经表征“人际脑同步”——当个体间发生社会互动时,其大脑活动之间会表现出一种脑同步提升的现象[3]。
近年来,多人同步交互扫描(超扫描)应运而生,即同时记录两人或多人互动过程。超扫描技术由Montague在2002年首次创用,是一种适用于自然教育情境的、高生态效度的研究范式[3]。不仅可以反映互动双方的心理、认知活动和学习策略,还可以探索互动过程中神经层面信息的传输和交换[4]。
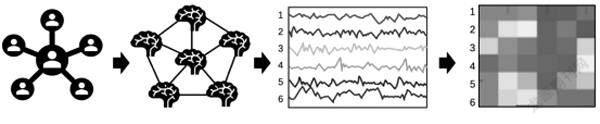
构建多脑网络主要关注的指标包括相关性、相干性、空间模式相似性,大致流程如图2所示。相关性是指用相关性分析两名被试的fNIRS脑活动数据,表示两人在互动过程中大脑活动在时域上的关联程度。相干性有两种:第一种是双脑活动的相干性,主要是指互动过程中互动双方的脑活动在频域上的关联性;第二种是双脑活动的小波相干性,即人际脑同步或者人际神经同步,是指互动双方的脑活动在特定时间、频率范围内的动态联系。空间模式相似性是指互动过程中不同个体目标脑区所构成的脑网络结构的相似性。
三、同伴互动神经机制的研究层次
从上述历程来看,个体层面研究不仅是神经科学探索大脑内部结构、内部功能与外部行为之间联系的基本内容,也是后续群体层面及整个互动领域的基石。为深入揭示多人同伴互动过程中行为、神经活动以及环境因素之间的动态联系,群体层面研究利用超扫描、小波变换相干等新颖实验和数据分析方法发掘群体互动行为与脑网络活动之间的动态联系。
(一)同伴互动的个体层面
个体层面的研究主要是在同伴互动的过程中,利用设备分别采集不同个体大脑神经活动数据,然后将其与行为数据进行协同分析,探究个体与他人发生互动行为时的认知过程、神经活动特征的差异以及影响因素,其数据收集和分析内容主要包括兴趣脑区的激活状态和程度、行为表现和任务绩效等。
基于不同实验方式和互动任务,先前研究已探索出不同行为受控于不同的大脑结构。例如:个体合作互动时,大脑的奖赏系统和颞顶联合区被激活,当进一步对互动形式进行细分时,言语互动和非言语互动行为所激活的脑区则又不尽相同,言语互动时初级听觉区、背外侧前额叶、眶额回和内侧前额叶会被激活[5],非言语互动又主要涉及镜像神经元系统(Mirror Neuron System,MNS)和心理化系统(Mentalizing System,MS) [6]。随着神经影像数据获取和分析技术的进步,越来越多的研究者进一步验证前额叶皮层、颞顶联合区、额极、顶叶、颞叶等区域在多人互动过程中的激活状态及人际脑同步特征[7-10]。
虽然近些年不少研究通过记录个体大脑活动的研究范式来探讨社会互动背后的神经机制,但通过文献梳理不难发现,不同任务下的互动行为所涉及的大脑结构及其活动状态也不尽相同。例如:双人面对面哼唱互动时激活的是右侧额下回[11];合作积木搭建时激活的是右侧前额叶和右侧颞上钩[12]。甚至稍稍改变同一任务特征,也会造成大脑活动差异。由此可见,不同互动任务涉及的大脑活动差异巨大,几乎难以找到一致性规律。研究需要探讨如何从群体层面展开,从分散个体数据提炼共性结论,发掘行为、神经活动特征和因素之间的动态联系。
(二)同伴互动的群体层面
神經影像技术发展使我们能从群体层面全面、深入地研究互动机制、影响因素及干预策略。认知神经机制研究与人际脑同步的测量有关。实证研究结果表明,人际脑同步是一个可以说明思维交互之间关系的可靠指标[13]。作为社交互动活动的神经标记,人际脑同步在社交及教育领域的许多研究中已被采用,如互动决策、沟通交流和互动教学等。
1. 人际脑同步的神经指标
早期超扫描研究包括发掘互动过程中人际脑同步的神经指标并创新多脑数据的分析方法。2011年,Funane等人首次利用fNIRS进行同伴互动超扫描研究,发现两人小组在互动完成合作按键任务过程中,彼此的前额叶皮层发生神经同步现象[14]。随后,Cui等人通过小波变换相干分析方法进一步发现,额上皮层的人际相干性能追踪两个同伴在0.08~0.3Hz(相当于周期3.2~12.8s)频率范围内的合作互动行为[15]。频率范围体现互动过程的时间结构,说明互动时的人际脑同步与互动学习任务本身的时间结构有关。从那时起,小波变换相干逐渐成为量化分析脑间同步性的流行方法。
随着技术和研究范式的发展,更多研究证实,当个体间发生互动行为时大脑之间会出现人际脑同步。众多研究提出两种具有代表性的假说:合作互动假说和相似任务假说。一方面,不少研究为合作互动假说提供支持,认为脑同步的出现可能意味着个体间发生了合作互动。大量研究发现合作性互动会发生脑同步[7],并且脑同步与教学效果或学习表现密切相关[16-18]。另一方面,也有研究为相似任务假说提供证据,表明当不同个体单独完成同样任务时,个体间脑同步仍有提升现象[19]。
2. 人际脑同步的影响因素
不少研究者探索了人际脑同步的影响因素。首先,小组内部的性别结构。例如:前额叶区域的人际脑同步很大程度上取决于同伴性别,异性同伴互动过程中会出现更高的人际脑同步,指出性别异质可能会促进双人同伴的小组合作[20]。其次,不同性别在互动中会影响互动行为和互动效果。例如:研究发现包含男性的小组互动绩效优于女性双人组[21]。最后,同伴关系的差异也会影响互动过程中的人际脑同步。在对恋人、朋友、陌生人三种关系的互动研究发现,恋人组的互动表现比朋友组和陌生人组更好,因为只有恋人组的上额皮层在互动过程中出现了人际脑同步,且其互动表现与人际脑同步的增强呈正相关[13]。这类研究为组内性别结构以及人际关系在同伴互动中的作用提供了重要的依据。
人际脑同步是探究同伴互动神经基础、互动质量及相关因素的重要指标,它与大脑神经响应的分析结果能够彼此补充,从而完善研究结论。然而,也有不少研究发现,在同伴互动过程中,个体与同伴发生人际脑同步的对应大脑区域不一定相同,这也暗示着未来教育研究中仅对学习者某一脑区的激活状态进行分析仍然无法确定该区域在整个互动过程中的作用。因此,想要验证“互动是否真实有效的发生”,要在此基础上进一步从群体层面分析。只有结合个体与群体层面的研究,才能构成描述同伴互动神经机制的完整画面。
四、同伴互动神经机制的研究焦点
本研究发现,同伴互动神经机制的研究焦点总体上可分为单一互动行为和复杂互动行为两方面。一方面,单一互动行为是同伴互动的基本元素,包括对话、手势、眼神等,其主要关注信息传递的认知过程及相对应的大脑结构及功能;另一方面,复杂互动行为针对社会和教育情境的复杂性,往往基于包含多个认知过程和互动行为的互动过程来探索其背后的神经基础和影响因素。两方面研究彼此补充,促进教育学与脑科学研究在不同情境场域下的深度融合,成为未来研究的热点和趋势。
(一)单一互动行为
1. 言语交流
言语交流是同伴互动最重要的形式。研究者大多基于不同的对话任务,探究不同互动方式、环境条件下发生同伴互动时个体大脑中的神经机制。
首先,基于不同互动形式,研究证明了面对面交流相比于远距离同伴互动的优势。研究者先通过在同样时间内获取两人交流时的脑活动数据来监控目标脑区的激活状态,再通过人际脑同步高低来评价交流的有效性。例如:有研究发现,只有在面对面对话的过程中,互动双方的左下额叶皮层的神经同步显著增加,并且这种神经同步主要来自伙伴之间的直接互动[22]。这些研究表明,面对面的对话交流具有其他类型的互动交流所没有的特殊神经特征,并且能够通过互动伙伴之间的人际脑同步反映出来,其研究结果不仅为项目式学习、跨学科STEM教育等协作学习新模式所具有的优势提供了脑科学证据,更为在线协作学习投入研究提供了新的研究思路。
其次,言语交流的参与人数也是研究焦点之一。最初的研究受设备和数据分析技术的限制,仅通过分析两人的脑活动数据来发现相关脑区激活及两人间的人际脑同步[22]。得益于超扫描技术的发展,研究者在实验中增加了研究人数。基于三人小组无领导讨论的任务,研究者针对左侧颞顶联合区(社会思考的重要领域),成功通过人际脑同步指标发现了小组讨论中的领导者[23]。四人同伴合作完成日语文字链游戏的研究证实了前额叶皮层在多人同伴互动中的作用[24]。不难发现,对互动团体规模的研究不仅验证了更多的神经指标,还能够帮助研究者识别个体在同伴互动中的不同角色,为课堂中的分组策略提供科学依据。
最后,基于不同言语交流任务的研究在验证相关脑区的同时,发现了更加有效的交流方式。例如:在协作完成创造性问题解决任务时,超扫描研究发现,轮流回答的方式比头脑风暴更能够激发学习者的互动兴趣、个体会表现出更多的换位思考行为,因而其团队的创造性表现也更好[10];另一项研究基于面对面的经济博弈任务指出,右侧颞定联合区的人际脑同步增强是同伴间积极互动的神经标志,也意味着面对面能够增加陌生同伴之间的共同意图,从而导致更加积极地相信合作决策并在游戏中获得更多实际收益[7]。
2. 注意与模仿
注意与模仿是简单互动行为研究的重要内容。注意是模仿行为发生的前提,是个体捕捉周围信息的重要行为,联合注意是指多人在第三个对象上的共同关注焦点,既可以由自我发起,也可以由同伴发起[25]。同伴发起的联合注意是个体的关注点能够跟随他人的手势和注视方向的能力,而自我发起的联合注意是指使用手势或眼神凝视来引导他人注意力的能力[26]。在同伴互动的过程中,主导者也可能会主动发起共同的注意力以引导学习者的注意力,而学习者在跟随主导者的指示时,也可能会接着发起其他注意请求。
对同伴互动中注意与模仿行为的研究大多是基于合作按键任务。实施时被试者双方面对面或并排坐在电脑前,根据屏幕中的闪光提示共同按键,或一人在尽可能短的时间内模仿同伴的按键行为。这一类研究主要探索了注意与模仿行为的神经指标,而基于不同模仿任务的研究发现,出现人际脑同步的脑区有所不同,有的研究通过对比被试在独自敲击键盘和双人互动模仿敲击键盘两种模式下的神经活动,确定了前运动皮层的人际脑同步可以作为模仿行为的神经指标[27],而其他基于按键任务的研究又将前额叶皮层的人际脑同步作为同伴互相模仿的神经指标,这意味着在教育情境下,学习者在不同的学习任务中与同伴进行互动时所利用的大脑区域和认知功能也有所不同。
(二)复杂互动行为
学习是一个复杂的认知活动,虽然基于单一互动行为的神经机制研究能够有效排除实验中的无关变量,清晰地揭示其背后的神经指标和相关因素,但也忽视了众多自然互动情境下无法避免的影响因素,限制了研究结论的现实意義。当前对复杂互动行为的研究主要集中在同伴交流的协调与交替、合作与竞争模式对互动效果的影响,下文对相关研究进行介绍,为未来丰富教育认知神经科学研究的研究情境、范式、理论基础提供参考。
1. 协调与交替
行为的协调与交替是同伴互动的重要特征,也是感知、理解和情感传播的重要媒介[28]。在同伴互动中,不同个体所扮演的角色有所不同,例如:在双人互动学习中,往往会存在一名“教师”和一名“学习者”。为了从“教师”那里获得技能和知识,“学习者”必须与“教师”协调自己的行为。根据合作互动假说,这种行为协调会潜在促进二人之间的人际脑同步,进而促进二者之间认知过程的协调[17]。此外,同伴互动还需要学习者之间的轮流交替行为,例如:在交谈的过程中轮流发表观点。
脑科学领域的研究多关注游戏、讨论、唱歌以及乐器合奏等互动过程中的协调合作,探索不同互动行为和干预因素下的神经活动特征。例如:有研究通过对比大学生在互动协调或独立两种模式下完成任务时左中额叶的人际脑同步,发现其能够作为同伴互动过程中亲社会效应的神经网络指标[29]。并且相对于独立模式,互动协调模式下的两人具有更高的共享意向,因而表现出更优的同步行为和更大的后续互助倾向[13]。还有研究初步探索了社会经济地位差异影响下亲社会交流行为的神经机制,发现差异较大的两人在互动时前额叶皮层的人际脑同步更高,并进一步解释亲社会倾向、社会经济地位与人际脑同步之间的相关性,从而提出了与共享认知相关的神经机制[29]。这类研究揭示了协调与交替在不同影响因素下的神经指标,为未来协作学习研究中的小组结构策略提供参考。
2. 合作与竞争
在同伴互动学习过程中,教师或指导者需要设计有效的干预策略以更高效地实现学习目标,其中,合作与竞争就是教育研究者和脑科学研究者常采用的干预策略。在该类实验中,被试往往会被告知将与同伴分别在合作或竞争情境下完成任务。
首先,探索合作与竞争模式下同伴互动行为的神经指标是该领域的研究热点。相互理解和心智理论是合作的重要前提,所涉及脑区有额叶、颞叶和杏仁核等,它们在处理社交信息方面的作用互为补充,以促使或阻碍个体间的合作[6]。例如:在合作互动时,两人的背膜前额叶皮层的人际脑同步增加,表明当同伴互动合作需要心智理论时,该区域可能会被激活[12]。此外,还有研究发现,合作过程中两人的右后颞上沟的人际脑同步增加,由于该区域涉及互动双方对彼此想法的理解,因此,判断其可以作为合作投入度增加的神经指标[7]。
其次,研究合作和竞争的任务形式非常多样,所验证的神经指标也各不相同。例如:基于按键任务的超扫描研究中,实验结果显示在合作模式下,两方的额上回皮层的人际脑同步增加[15];当两人小组合作完成基于计算机的建造游戏时,他们的协助行为会激活其伙伴的双侧顶叶下顶叶,而在竞争条件下,该脑区的激活程度则会受到对手的阻碍行为的影响而减弱[30]。还有研究基于搭积木的互动任务,发现双人同伴在合作和竞争模式下的互动过程中都会出现右侧额中回和额上回区域人际脑同步的增加,表明该区域参与了目标导向的同伴互动,例如:复杂的搭建动作和思考决策[12]。除了对脑区激活和人际脑同步的探讨,在任务绩效上,研究发现在完成创造性任务时,合作模式的互动效果要比竞争模式更好[31]。
最后,针对团体合作过程,不少研究探讨了团体成员构成对互动合作效果的影响,其研究结论却不尽相同。有的研究发现,在解决创造性问题时,创造力较低的两人的合作绩效更优,且对神经指标和行为指标的数据分析结果都表明,当两个较低创造力的人一起解决创造力问题时,他们倾向于相互合作[8]。与其相反,还有一些研究则发现,同伴的智商与创造性任务成果的创造性、流畅性和独创性正相关[9]。对于性别因素,研究则认为,性别构成不会对同伴完成创造性任务的效果产生影响[32]。
上述研究为探索不同情境、任务及干预下互动行为及效果背后的神经机制提供了一些指导和应用建议,但对教育研究的启示还存在局限。首先,由上文可知,人们完成任务时发生互动行为所利用的脑区及其神经活动状态有很大差异,即使是在完成同一任务时受不同因素影响也会造成神经活动的差异,很难找到其共有的特征和规律,教育研究应当采取哪些任务来探索其普遍机制呢?其次,无论基于什么设备,数据分析时采用的方法都是将多个被试、多次实验所获得的数据进行平均后进行“配对成像相减”的方法得出差异值,这种结果往往只能说明特定脑区在特定实验任务和条件下的激活状态,不能说明该指标能够反映所有相似任务中的互动有效性[33]。最后,大多相关研究面临生态效度问题,fMRI研究仅限于被试两人平躺在设备中进行话语或小动作的互动交流,而多通道的EEG和fNIRS虽然不会限制被试的手部活动,但多数需要被试静坐在椅子上完成实验任务。真实教育场景下的同伴互动是复杂的、持续的且受诸多环境因素影响的,这对神经影像技术的应用提出了新挑战。
五、未来研究展望
随着研究方法的更新进步,神经影像技术在教育神经领域的应用有巨大潜力。本研究结合上述分析,提出如下展望:
(一)实现多维数据融合,构建同伴互动过程的分析模型
除大脑内部的神经活动以外,同伴互动过程中还会产生文本、对话、表情、手势等多样态的交互数据,多源数据有效融合才能精准反映学习者与同伴互动的学习效果。因此,将神经活动数据、行为数据以及调查数据进行协同分析,并科学解释其中蕴含的实际教学意义是研究面临的重要任务之一。未来研究需要借助神经影像技术、学习分析技术、观察访谈、问卷调查等多种途径来获取同伴互动过程中的生理、神经、认知活动以及社会交互等数据,将来自不同时间和空间维度的多模态数据彼此补充、有效融合,构建多维分析模型,为后续研究建立基础的理论框架,这对于教育认知神经学视角下的同伴互动分析研究具有重要的基础性意义。
(二)聚焦具体教育场景,促进认知神经技术在教育中的应用
本文对研究焦点进行总结后发现利用神经影像技术探索同伴互动学习的认知过程的研究依然有限,缺少真实的教学场景和具有教育意义的实验任务,且任务时长也较短,因而其研究结论难以被教育研究者借鉴。新冠肺炎疫情时期,在线学习的价值迅速凸显,且STEM教育、创客教育和项目式学習等新型学习模式也在教育实践中得到越来越多的应用,但大量教师却因缺乏科学指导而无法选择有效的协作模式和互动策略。未来研究需要细化不同教育情境、教学方式、学科任务等场域下的同伴互动,加强认知神经科学与教育教学研究者之间的跨领域对话与合作,通过应用跨学科的知识、研究方法和技术促进认知神经技术在教育领域的应用,丰富教育认知神经科学的理论和知识体系。
(三)关注个体差异,为个性化互动策略提供依据
认知神经机制不仅是揭示认知过程的有效手段,也可有效证明其对个体学习效果和能力提升的作用,可否通过认知神经技术对互动过程不同阶段中的影响因素进行识别,并对表现欠佳的学习者进行诊断呢?倘若能够找到大脑结构发育或认知活动障碍的缘由,教师就能有效选择干预或治疗措施来提升互动水平,从而影响学习效果。目前已有研究探讨了学习者同伴互动心理等因素对个体在团队中互动行为及情感投入的影响[34]。接着,进一步通过脑扫描实验对比探究同伴互动中大脑活动的个体差异,推断不同学习者的认知能力发展需求,进而采取更有针对性的个性化学习策略。
(四)积极研发便携式神经影像设备,拓展研究情境
虽然当前神经影像设备的性能比起若干年前已有很大提升,但其便携性和生态效度仍有限。情境认知和具身认知的研究表明,身体的放松姿态、自由流畅的手部活动、外部空间的自由等,均对创造力有着显著影响[33]。实际上,成本低、易操作、通道数量较少的便携式神经影像设备亦可满足对互动过程中神经活动的初步探索。例如:双通道fNIRS设备和依托脑机接口技术开发的穿戴式脑电设备就能够探索课堂情境下的数学学习活动的神经机制[35-36]。因此,未来鼓励教育技术、认知神经科学和计算机工程领域学者的跨学科合作,研发能够即时、持续地采集多模态行为或生理数据的设备和技术平台。
(五)以教育理论为主要指导,整合多元化研究方法
教育神经科学视角下的同伴互动研究可获得关于该过程的三维数据:神经活动数据、外显行为数据以及内隐的情感态度数据。受限于单一学科研究方法,大多数研究未能将三类科学证据很好关联,因而难以用于建构内涵丰富、科学的教育知识和理论,也难以指导教育实践。未来研究需要依托相关理论开展,利用各种神经影像技术、情感分析、行为监测以及基于人工智能的数据分析等技术手段,整合神经影像实验法、准实验法、行动研究法等多种研究范式,为教育认知神经科学领域的研究提供解决问题、知识创新的新路径。
[參考文献]
[1] 徐锦芬,舒静.基于语料库的二语学习者互动口语研究新视野——《运用语料库探索英语学习者口语:学习者会话》述评[J].外语电化教学,2018(5):91-94.
[2] ANDREWS J J, RAPP D N. Benefits, costs, and challenges of collaboration for learning and memory[J]. Translational issues in psychological science, 2015, 1(2): 182-191.
[3] MONTAGUE P R, BERNS G S, COHEN J D, et al. Hyperscanning: simultaneous fmri during linked social interactions[J]. Neuroimage, 2002, 16(4): 1159-1164.
[4] 朱朝喆.近红外光谱脑功能成像[M].北京:科学出版社,2020:125-127.
[5] BABILONI F, ASTOLFI L. Social neuroscience and hyperscanning techniques: past, present and future[J]. Neuroscience & biobehavioral reviews, 2014, 44: 76-93.
[6] WANG M Y, LUAN P, ZHANG J, et al. Concurrent mapping of brain activation from multiple subjects during social interaction by hyperscanning: a mini-review[J]. Quantitative imaging in medicine & surgery, 2018, 8(8): 819-837.
[7] TAO L, SAITO G, LIN C, et al. Inter-brain network underlying turn-based cooperation and competition: a hyperscanning study using near-infrared spectroscopy[J]. Scientific reports, 2017, 7(1): 8684.
[8] XUE H, LU K, HAO N. Cooperation makes two less-creative individuals turn into a highly-creative pair[J]. Neuroimage, 2018,172: 527-537.
[9] MAYSELESS N, HAWTHORNE G, REISS A L. Real-life creative problem solving in teams: fnirs based hyperscanning study[J]. Neuroimage, 2019, 203: 116161.
[10] LU K, YU T, HAO N. Creating while taking turns, the choice to unlocking group creative potential[J]. Neuroimage, 2020, 219: 117025.
[11] OSAKA N, MINAMOTO T, YAOI K, et al. How two brains make one synchronized mind in the inferior frontal cortex: fnirs-based hyperscanning during cooperative singing[J]. Frontiers in psychology, 2015, 6(127): 1811.
[12] NING L, CHARIS M, WITT EE, et al. Nirs-based hyperscanning reveals inter-brain neural synchronization during cooperative jenga game with face-to-face communication[J]. Frontiers in human neuroscience, 2016, 10: 00082.
[13] PAN Y, CHENG X, ZHANG Z, et al. Cooperation in lovers: an fnirs-based hyperscanning study[J]. Human brain mapping, 2017, 38(2): 831-841.
[14] FUNANE T, KIGUCHI M, ATSUMORI H, et al. Synchronous activity of two people's prefrontal cortices during a cooperative task measured by simultaneous near-infrared spectroscopy[J]. Journal of biomedical optics, 2011, 16(7): 77011.
[15] CUI X, BRYANT D M, REISS A L. Nirs-based hyperscanning reveals increased interpersonal coherence in superior frontal cortex during cooperation[J]. Neuroimage, 2012, 59(3): 2430-2437.
[16] DIKKER S, WAN L, DAVIDESCO I, et al. Brain-to-brain synchrony tracks real-world dynamic group interactions in the classroom[J]. Current biology, 2017, 27(9): 1375-1380.
[17] PAN Y, NOVEMBRE G, SONG B, et al. Interpersonal synchronization of inferior frontal cortices tracks social interactive learning of a song[J]. Neuroimage, 2018, 183: 280-290.
[18] ZHENG L, CHEN C, LIU W, et al. Enhancement of teaching outcome through neural prediction of the students' knowledge state[J]. Human brain mapping, 2018, 39(2): 3046-3057.
[19] NUMMENMAA L, GLEREAN E, VIINIKAINEN M, et al. Emotions promote social interaction by synchronizing brain activity across individuals[J]. Proceedings of the national academy of sciences, 2012, 109(24): 9599-9604.
[20] CHENG X, LI X, HU Y. Synchronous brain activity during cooperative exchange depends on gender of partner: a fnirs-based hyperscanning study[J]. Human brain mapping, 2015, 36(6): 2039-2048.
[21] BAKER J M, LIU N, CUI X, et al. Sex differences in neural and behavioral signatures of cooperation revealed by fnirs hyperscanning[J]. Scientific reports, 2016, 6(1): 26492.
[22] JIANG J, DAI B, PENG D, et al. Neural synchronization during face-to-face communication[J]. The journal of neuroscience, 2012, 32(45): 16064-16069.
[23] JIANG J, CHEN C, DAI B, et al. Leader emergence through interpersonal neural synchronization[J]. Proceedings of the national academy of sciences, 2015, 112(14): 4274-4279.
[24] NOZAWA T, SASAKI Y, SAKAKI K, et al. Interpersonal frontopolar neural synchronization in group communication: an exploration toward fnirs hyperscanning of natural interactions[J]. Neuroimage, 2016, 133: 484-497.
[25] OBERWELLAND E, SCHILBACH L, BARISIC I, et al. Look into my eyes: investigating joint attention using interactive eye-tracking and fmri in a developmental sample[J]. Neuroimage, 2016, 130: 248-260.
[26] HECKE A, MUNDY PC, ACRA CF, et al. Infant joint attention, temperament, and social competence in preschool children[J]. Child development, 2007, 78(1): 53-69.
[27] HOLPER L, SCHOLKMANN F, SHALOM DE, et al. Extension of mental preparation positively affects motor imagery as compared to motor execution: a functional near-infrared spectroscopy study[J]. Cortex, 2012, 48(5): 593-603.
[28] WILSON M, WILSON TP. An oscillator model of the timing of turn-taking[J]. Psychonomic bulletin & review, 2005,12(6):957-968.
[29] BALCONI M, FRONDA G, VANUTELLI ME. Donate or receive? social hyperscanning application with fnirs[J]. Current psychology, 2019, 38(4): 991-1002.
[30] LIU T, SAITO H, OI M. Role of the right inferior frontal gyrus in turn-based cooperation and competition: a near-infrared spectroscopy study[J]. Brain and cognition, 2015, 99: 17-23.
[31] LU K, XUE H, NOZAWA T, et al. Cooperation makes a group be more creative[J]. Cerebral cortex, 2019, 29(8): 3457-3470.
[32] LU K, TENG J, HAO N. Gender of partner affects the interaction pattern during group creative idea generation[J]. Experimental brain research, 2020, 238(5): 1157-1168.
[33] 郝寧.创造力的神经机制及其教育隐意[J].全球教育展望,2013,42(2):63-73.
[34] MCGILLICUDDY D. "They Would Make You Feel Stupid"—ability grouping, children's friendships and psychosocial wellbeing in Irish primary school[J]. Learning and instruction, 2021, 75: 101492.
[35] NOZAWA T, SAKAKI K, IKEDA S, et al. Prior physical synchrony enhances rapport and inter-brain synchronization during subsequent educational communication[J]. Scientific reports, 2019, 9(1): 12747.
[36] QU Z, CHEN J, LI B, et al. Measurement of high-school students' trait math anxiety using neurophysiological recordings during math exam[J]. IEEE access, 2020, 8: 57460-57471.
