鲫鱼源维氏气单胞菌噬菌体的分离及生物学特性
2022-01-17戴瑜来戴杨鑫许宝青林启存马恒甲蔡丽娟
戴瑜来,戴杨鑫,许宝青,林启存,马恒甲,蔡丽娟
(杭州市农业科学研究院,浙江 杭州 310024)
维氏气单胞菌(Aeromonasveronii)是一种人—兽—鱼共患的条件致病菌[1],该菌可感染草鱼[2]、鲫[3]、泥鳅[4]、青虾[5]、中华绒螯蟹[6]、中华条颈龟[7]、达氏鲟[8]、虹鳟[9]、大黄鱼[10]等多种水生动物,引发的疾病发病快、死亡率高,对水产养殖业造成了巨大经济损失。研究人员发现,维氏气单胞菌对β-内酰胺类(氨苄西林钠、苯唑西林)、大环内酯类(硫氰酸红霉素、乙酰螺旋霉素)、喹诺酮类(恩诺沙星)、氨基糖苷类(硫酸新霉素、卡那霉素)、四环素类(四环素、强力霉素)、磺胺类(磺胺嘧啶、磺胺间甲氧嘧啶、磺胺甲噁唑)、氯霉素类(甲砜霉素、氟苯尼考)等抗菌药均产生了一定的耐药性[11-15],且极可能会随着抗菌药的继续滥用而在水生动物临床上更加快速且广泛地传播。同时,抗菌药的广谱性会破坏水体环境及养殖动物肠道中正常微生物群系的平衡,影响养殖鱼类的生长发育,进而造成水产养殖效率低下。因此,寻找安全、高效、环保的生物抑菌剂具有重要意义。噬菌体(Baceriophage)是一类能够感染细菌、真菌、放线菌或螺旋菌等微生物的病毒总称,它具有裂解宿主专一性强、研发时间短,不影响正常菌群,对人和动物安全,耐药风险低且无环境污染等优点,其在细菌病防控领域的应用研究呈逐年上升趋势[16]。目前,已报道用于水生动物细菌病防控的噬菌体研究数量有限,为了丰富水产致病菌的噬菌体库,筛选具有潜在应用价值的菌株,本试验以维氏气单胞菌为宿主菌,分离筛选烈性噬菌体,研究其生物学特性,旨在为进一步探索噬菌体防控技术提供科学依据。
1 材料与方法
1.1 菌株 维氏气单胞菌(XJJYCA01)分离自发病鲫鱼,LB斜面保存于4 ℃。
1.2 试剂与仪器 试剂耗材:LB培养基、SM液、接种环、5 mL注射器、0.22 μm过滤头、一次性培养皿等,均购自生工生物工程(上海)股份有限公司。仪器:UV-2600紫外分光光度计、离心机(Eppendorf Centrifuge 5427R和Hermle Z323K)、HS-2恒温水浴锅、Hitachi H-7650型电子显微镜、JJ-Y系列电子天平、生化培养箱等。
1.3 噬菌体分离与纯化 参照王志丽[17]的双层平板法进行维氏气单胞菌的富集、筛选、纯化,略作修改。步骤如下:(1)无菌操作解剖,取发病鲫鱼肠道置于灭菌培养皿中,用生理盐水洗脱肠道内容物,4 000 r/min离心10 min后过0.22 μm滤头备用;(2)挑取维氏气单胞菌(XJJYCA01)单菌落接种到含有5 mL LB液体培养基的试管中,28 ℃ 120 r/min摇床上过夜培养;翌日,将其转到含25~30 mL液体培养基的锥形瓶中,28 ℃ 120 r/min震荡培养3 h,使其达到对数期(OD值0.4~0.6),然后向菌液中加入5 mL病鱼肠道洗脱液,再28 ℃振荡培养48 h。之后将混合液以12 000 r/min离心10 min,小心吸取上清转移至另一无菌螺旋离心管后备用;(3)取100 μL分离液与200 μL对数期的宿主菌混合,放置5~15 min后加到融化并冷却至45 ℃的半固体培养基中,混匀立即倒入底层琼脂平板上,铺匀,28 ℃ 倒置培养24 h。翌日观察噬菌斑的生成情况;(4)用灭菌牙签挑取单个噬菌斑,接到5 mL液体培养基中富集10 min,然后向其中加入200 μL宿主菌菌液,28 ℃培养2~3 h至液体澄清后,12 000 r/min离心取上清,即得到高效价的噬菌体。
1.4 噬菌体效价的测定 采用噬斑法测定噬菌体效价。吸取100 μL纯化的噬菌体液进行10倍稀释,将稀释的噬菌体液分别与200 μL的XJJYCA01混合,利用双层平板法观察噬菌斑的个数。效价(PFU/mL)=噬菌斑数×10×稀释倍数。
1.5 噬菌体的电镜观察 参照Ramíre-orozco等[18]的方法,采用磷钨酸负染法,用Hitachi H-7650型电子显微镜进行电镜观察。
1.6 噬菌体最适感染复数(MOI)测定 培养XJJYCA01至对数生长期(OD600≈0.5,约为1×108CFU/mL),按照不同的感染复数(1 000,100,10,1,0.1,0.01,0.001)各取0.2 mL混合,并加入1.6 mL的LB液体培养基,28 ℃培养4 h。经培养后,9 000 r/min离心10 min,0.22 μm过滤。收集上清,测定噬菌体效价,以产生最高噬菌体效价的MOI为最佳感染复数。
1.7 一步生长曲线 参照柏琴琴[19]并略作修改。具体操作如下:将XJJYCA01培养至对数生长期。将噬菌体SJTJYCA01效价浓度稀释到1×108PFU/mL。1 mL噬菌体稀释液和10 mL宿主菌细菌悬液混合于锥形瓶P1中,混合10 min后,取0.1 mL加入到含有10 mL LB培养液的锥形瓶P2中,混合均匀,再从锥形瓶B2中取0.2 mL至含有20 mL液体培养基的P3中,混合均匀后在28 ℃及180 r/min条件下振荡培养。从0开始每隔一段时间取样。所取样品经9 000 r/min离心2 min,取上清液用双层平板法测定过滤液中噬菌体的效价。以取样时间为横坐标,噬菌体效价的对数值为纵坐标,绘制一步生长曲线。裂解量=裂解期平均噬菌体数/潜伏期平均噬菌体数。
1.8 热稳定性及pH稳定性 将噬菌体SJTJYCA01裂解液稀释至1×1010PFU/mL,分装到3个5 mL 离心管中,每管各装3 mL。将离心管分别放置于40 ℃、60 ℃、80 ℃的恒温水浴锅中,每隔20 min测定各管中噬菌体的效价,直至测到60 min。以SM液为介质,用NaOH和HCl调节pH(3~11)。将噬菌体SJTJYCA01裂解液稀释至1×1010PFU/mL,取10 μL加入到990 μL不同pH的SM液中,28 ℃水浴2 h后测定各离心管中噬菌体的滴度。每个pH试验重复3次。
2 结果
2.1 噬菌体分离与纯化 从发病鲫鱼肠道中获得的分离液经双平板法后在平板上可见有大量噬菌斑产生,图1A可见噬菌斑呈圆形,中间透明,边缘清晰整齐,无晕环,但大小不一,分布不均匀,直径在0.2~1.1 mm,噬菌斑的存在说明肠道分离液中有宿主菌的噬菌体存在。噬菌体经纯化增殖后,梯度稀释,用双层平板法计数,该噬菌体的效价可达3.0×1012PFU/mL,噬菌体命名为SJTJYCA01。

图1 噬菌体的噬菌斑(A)和超微形态图(B)Fig.1 Plaque morphology (A) and TEM image (B) of phage
2.2 噬菌体的超显微形态 通过电子显微镜观察发现,本试验分离的噬菌体为有尾噬菌体,属于尾病毒目短尾噬菌体科,头部直径在45~60 nm,尾部长在16~26 nm,见图1B。
2.3 最适感染复数 噬菌体SJTJYCA01和宿主菌XJJYCA01的MOI为0.1时的效价最高(效价值达2.55×109),说明噬菌体SJTJYCA01和宿主菌XJJYCA01的最适感染复数为0.1。见表1。

表1 噬菌体SJTJYCA01的最适感染复数Table 1 Determination of optimal MOI of SJTJYCA01
2.4 一步生长曲线 一步生长曲线的结果见图2,噬菌体SJTJYCA01的潜伏期为20 min,裂解期达40 min,裂解量为裂解期平均噬菌体数(14 458 PFU/mL)/ 潜伏期平均噬菌体数(255 PFU/mL)=57。
2.5 热稳定性 在40 ℃条件下,噬菌体SJTJYCA01的存活率不随时间的增加而减少;在60 ℃条件下,噬菌体SJTJYCA01的存活率大幅下降,作用20 min后存活率仅为原噬菌体的3%;在80 ℃条件下,各个时间段均未见有噬菌体存活。见图3。

图2 噬菌体的一步生长曲线Fig.2 One-step growth curve of phage
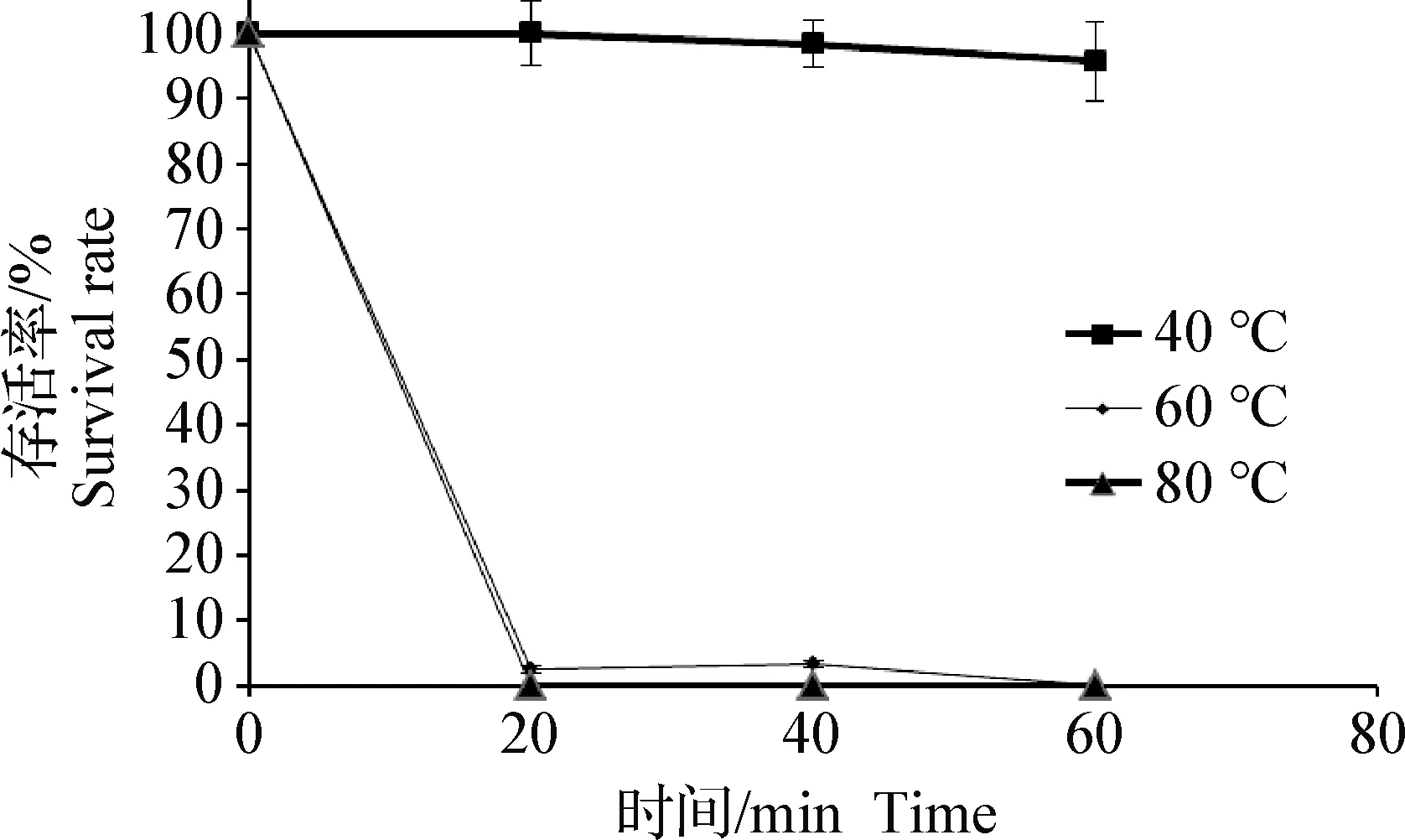
图3 噬菌体SJTJYCA01的热稳定性Fig.3 Thermal stability of SJTJYCA01
2.6 pH稳定性 噬菌体SJTJYCA01对强酸强碱均较为敏感,在pH 5~10时较为稳定;噬菌体SJTJYCA01的最适pH 7。见图4。

图4 噬菌体SJTJYCA01的pH稳定性Fig.4 pH stability of SJTJYCA01
3 讨论
近年来,国内外有关维氏气单胞菌病例的报道逐年增多,其流行呈明显上升趋势,并已成为一种危害严重的人—兽—鱼共患病原菌[20]。噬菌体作为一种新兴的生物抑菌剂日渐受到学者们的关注[21]。目前,国内外关于维氏气单胞菌噬菌体的研究鲜有报道,仅可见Taruna等[22]从池水中分离到维氏气单胞菌的噬菌体VTCCBPA5和吕孙建等[23-24]从中华鳖养殖塘中分离得到的非特异性宽谱噬菌体NTHP01(以嗜水气单胞菌为宿主分离获得)。
本试验以鲫鱼源维氏气单胞菌为宿主菌,从发病鲫鱼的肠道中分离获得噬菌体SJTJYCA01。该噬菌体在宿主菌上可形成圆形、透亮,具有典型烈性噬菌体特征的噬菌斑[25];参照《病毒分类——国际病毒分类委员会第九次报告》,对噬菌体SJTJYCA01的形态进行观察分析,结果可判定该噬菌体为尾病毒目短尾噬菌体科噬菌体(头径45~60 nm,尾长16~26 nm),此与Taruna等[22]分离获得的VTCCBPA5同属于短尾噬菌体科(头径56 nm,尾长5 nm),却有别于吕孙建等[24]分离获得的肌尾噬菌体科噬菌体NTHP01(头径60~145 nm,尾长80~455 nm)。因此,这是鲫鱼源维氏气单胞菌短尾科烈性噬菌体在中国的首次报道。
感染复数是研究噬菌体与宿主之间量化与产出的重要生物学指标,在不同感染复数条件下,噬菌体感染宿主菌后分裂得到的噬菌体子代的数量不同,因此,确定最适感染复数对于噬菌体进一步的应用研究具有重要作用。本试验获得噬菌体SJTJYCA01的最适感染复数为0.1,与王志丽[17]分离的鳖源气单胞菌噬菌体的最适感染复数相同,说明该噬菌体裂解效率较高,少量的噬菌体侵染宿主菌即可产生噬菌体子代;噬菌体感染宿主菌的潜伏期和裂解期的长短及裂解量的大小通常与噬菌体、宿主菌及生长条件有关,从潜伏期可得知噬菌体吸附于细胞并释放出子代噬菌体的最短时间,潜伏期和裂解量则是噬菌体增殖的2个特征性指标。噬菌体SJTJYCA01潜伏期和裂解期分别为20 min、40 min, 较噬菌体VTCCBPA5(潜伏期35 min)[22]、泥鳅源气单胞菌的噬菌体pAh1(潜伏期30 min)[26]和鳖源气单胞菌的噬菌体(潜伏期50 min)[17]要短,远小于噬菌体NTHP01(潜伏期150 min)[23],且同样对宿主菌具有较高的裂解量。
目前,气单胞菌属细菌(包括维氏气单胞菌)能造成多种水生动物发生运动性气单胞菌败血症[27],该病在浙江、湖南、湖北、江西等地的发病高峰期为每年的6~9月(养殖水温在20~30 ℃,pH一般呈弱碱或弱酸性),常常出现急性暴发病,严重时的死亡率可达90%以上。本试验分离获得的噬菌体SJTJYCA01在温度低于40 ℃时能保持较好的稳定性和活性,并在pH 7附近最为稳定。因此,噬菌体SJTJYCA01能很好的适应水产养殖环境,在水生动物细菌性疾病防控中应用具有极大的可能性。本试验筛选获得的维氏气单胞菌烈性噬菌体,不仅丰富了水产致病菌噬菌体的菌种资源,并将为噬菌体作为生物抑菌剂的开发应用提供科学依据。
