干扰lncRNA SNHG22抑制EPHA3基因对结直肠癌LoVo细胞周期与凋亡的影响
2022-01-04姜进平朱钟钟黄耿程凯罗海平
姜进平 朱钟钟 黄耿 程凯 罗海平
1鄂东医疗集团黄石市中心医院(湖北理工学院附属医院)胃肠肛肠外科 435000;2鄂东医疗集团黄石市中心医院(湖北理工学院附属医院)泌尿外科 435000
结直肠癌在大多数国家和地区的病例数呈现逐年增加的趋势,是肿瘤相关死亡的主要原因之一[1]。对于中晚期结直肠癌患者,其预后较差,5 年生存率非常低[2]。探究结直肠癌发生的调控机制,有助于开发新的诊断标志物和治疗靶标。长链非编码RNA(lncRNA)是一类新型小分子非编码RNA,因为不具有编码蛋白潜力,曾被认定为转录的“噪音”[3]。最近研究发现,lncRNA在染色质重塑、染色体失活、转录激活等生物过程中发挥重要功能[4]。lncRNA 已被证实作为抑癌基因或癌基因影响包括结直肠癌在内的很多肿瘤的发生、发展[5]。据报道,SNHG22 在甲状腺癌、卵巢癌、胃癌等肿瘤中高表达,是恶性肿瘤进展的重要因素[6-8]。SNHG22 在结直肠癌中的生物学功能和潜在分子机制尚不清楚。本研究探讨了干扰SNHG22表达对结直肠癌LoVo细胞周期和凋亡的影响,探讨其在结直肠癌细胞中可能的作用机制。
1 材料与方法
1.1 细胞与主要试剂 结直肠癌LoVo 细胞购于美国典型培养物保藏中心;针对SNHG22 的小干扰RNA(si-SNHG22)、小干扰RNA 阴性对照(si-NC)购于上海GenePharma 公司;DMEM 培养基购于美国Gibco 公司;细胞周期试剂盒购于美国Sigma公司;Lipofectamine 3000购自美国Invitrogen 公司;细胞凋亡试剂盒购于北京索莱宝生物科技有限公司;实时定量聚合酶链反应(qRT-PCR)试剂盒购于日本TaKaRa公司;抗体GAPDH、Cyclin H、Caspase6、CDK3、促红细胞生成素产生肝细胞A 型受体3(erythropoietin-producing hepatocellular receptor A3,EPHA3)购于美国CST公司。
1.2 细胞培养和感染 将LoVo 细胞在含10%胎牛血清的DMEM 培养基培养,保持在37 ℃、5%CO2环境下生长。转染前一天,接种LoVo 细胞在12 孔板,使用Lipofectamine 3000 将si-SNHG22 或si-NC 转染到LoVo 细胞,即si-SNHG22 组和si-NC 组。转染5 h 后,替换为完全DMEM培养基。
1.3 qRT-PCR 检测SNHG22 和EPHA3 信 使RNA(mRNA)的表达 采用TRIzol试剂从转染后的LoVo 细胞中提取总RNA,采用PrimeScript RT 试剂盒逆转录为cDNA。qRT-PCR 扩增以GAPDH 为内源性对照。引物序列如下:SNHG22 正向引物5’-AGGAGAGCTGCTCTTCACAGG-3’,反向引物5’-TCCTAGGCTGAGTGTGTCTCC-3’;EPHA3 正向引物5’-ACTCTACGAGACTGCAATAGCA-3’,反向引物5’-TCCCCAAGATCCATTTGAGTGA-3’。采用2-ΔΔCt方法进行SNHG22和EPHA3 mRNA表达的计算。
1.4 流式细胞术评估LoVo 细胞周期 收集转染后的LoVo细胞,70%体积分数的乙醇重悬固定5 h。采用磷酸盐缓冲液洗涤3次,加入PI试剂和RNA酶试剂,在培养箱内避光染色35 min。通过流式细胞仪分析LoVo细胞周期分布。
1.5 流式细胞术评估LoVo 细胞的凋亡 收集转染后的LoVo 细胞,采用磷酸盐缓冲液洗涤3 次,Annexin V-FITC试剂重悬细胞,加入PI 染液,4 ℃下避光染色35 min,补充Annexin V-FITC 试剂,使用流式细胞仪检测LoVo 细胞早期凋亡和晚期凋亡,两者之和即为细胞凋亡率。
1.6 Western blot 检测蛋白表达 通过RIPA 缓冲液提取转染后的LoVo 细胞总蛋白,十二烷基硫酸钠-聚丙烯酰胺凝胶电泳分离40 μg 蛋白裂解物,转移至聚偏二氟乙烯膜。10%脱脂牛奶将膜进行封闭。与一抗在4 ℃下孵育过夜(GAPDH、EPHA3、Cyclin H、Caspase6、CDK3 抗体均按1∶1 000 稀释),然后与辣根过氧化物酶标记的二抗孵育4 h。采用ECL试剂显影目标蛋白条带。
1.7 统计学分析 采用SPSS 22.0软件分析实验数据,计量资料符合正态分布,均以均数±标准差(±s)表示,每个独立实验均重复3 次,组间数据之间统计分析应用独立样本t检验,以P<0.05为差异有统计学意义。
2 结 果
2.1 SNHG22 在LoVo 细胞中的干扰情况 si-NC组 和si-SNHG22 组SNHG22 的表达分别为(1.07±0.15)和(0.22±0.04),si-NC 组SNHG22 表达是si-SNHG22 组的4.86 倍(P<0.01),提示干扰SNHG22 明显降低LoVo 细胞中SNHG22的表达水平。
2.2 干扰SNHG22 阻滞LoVo 细胞周期 细胞周期结果(图1)显示,与si-NC 组相比,干扰SNHG22 明显增加LoVo 细胞的G0~G1 期细胞比例(P<0.01),明显减少S 期细胞比例(P<0.05)。
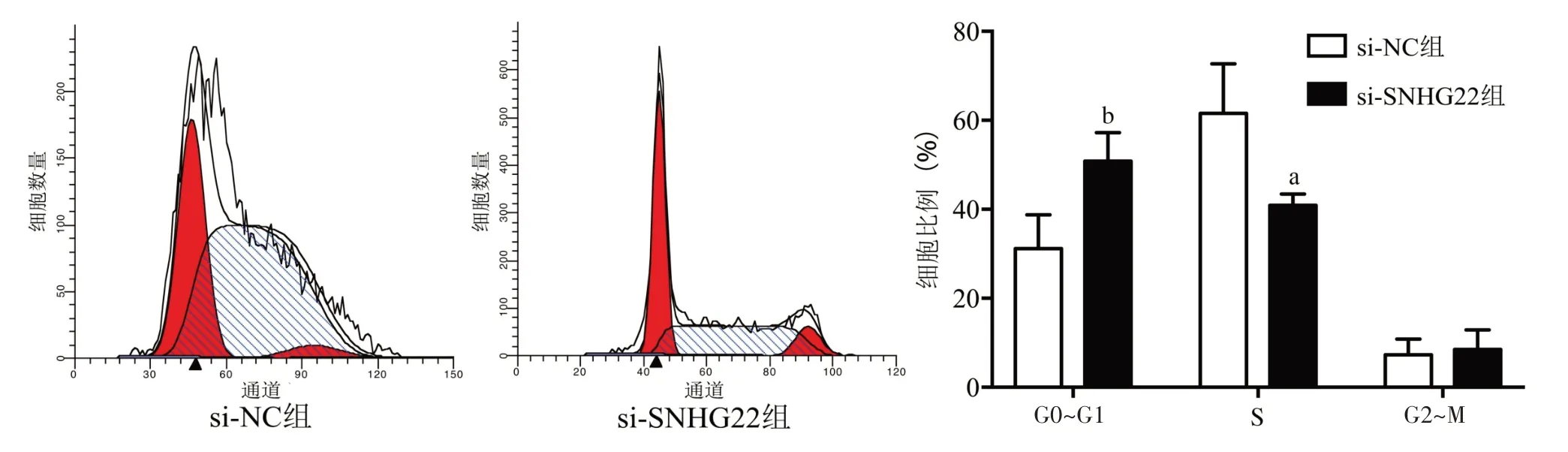
图1 干扰SNHG22对LoVo细胞周期的影响
2.3 干扰SNHG22 促进LoVo 细胞的凋亡 细胞凋亡结果(图2)显示,si-NC组和si-SNHG22组LoVo细胞凋亡率分别为(10.84±3.22)%和(40.61±5.96)%。与si-NC 组相比,干扰SNHG22后,LoVo细胞的凋亡率明显增加(P<0.01)。

图2 干扰SNHG22对LoVo细胞凋亡的影响
2.4 干扰SNHG22 抑制LoVo 细胞中EPHA3 mRNA 表达 si-NC 组和si-SNHG22 组LoVo 细胞中EPHA3 mRNA的表达分别为(1.31±0.12)和(0.31±0.06),与si-NC 组相比,干扰SNHG22 显著降低LoVo 细胞EPHA3 mRNA 表达(P<0.01)。
2.5 EPHA3 蛋白和凋亡及周期相关蛋白的表达Western blot 结果(图3)表明,与si-NC 组相比,干扰SNHG22 明显降低LoVo 细胞EPHA3 蛋白水平,显著提高细胞凋亡蛋白Caspase6 表达水平,显著降低细胞周期蛋白Cyclin H、CDK3表达水平。

图3 干扰SNHG22对LoVo细胞EPHA3蛋白和凋亡、周期相关蛋白表达的影响
3 讨 论
迄今为止,lncRNA 如SNHG6[9]、miR4435-2HG[10]、NEAT1[11]、CCAL[12]等被发现在结直肠癌组织或细胞株重表达失调,参与调节细胞周期、细胞凋亡、细胞转移等各种生物过程。作为一种新型肿瘤促进因子,SNHG22在卵巢癌组织中表达显著增加,并且与卵巢癌的低分化水平显著相关,SNHG22 可以促进卵巢癌细胞的化疗耐药性,而敲低SNHG22 表达可以增强卵巢癌细胞对顺铂和紫杉醇的敏感性[6]。在甲状腺乳头状癌组织和细胞株中观察到高表达的SNHG22,其高表达水平与甲状腺乳头状癌患者不利的临床病理特征和较差的总生存率密切相关,敲低SNHG22 通过调节miR-429表达在体外有效抑制甲状腺乳头状癌细胞增殖、迁移和侵袭,加速细胞凋亡[8]。SNHG22 在胃癌组织中高表达,SNHG22通过结合miR-361-3p促进胃癌细胞增殖、迁移、侵袭、细胞周期,抑制胃癌细胞凋亡[7]。以上的研究表明,SNHG22 是一个致癌lncRNA。SNHG22 对结直肠癌进展的影响尚需进一步研究。
为了确定SNHG22 在结直肠癌细胞中的详细功能,本研究利用RNA 干扰技术下调LoVo 细胞中SNHG22 的表达水平,随后在对LoVo 细胞周期和凋亡检测中发现,干扰SNHG22表达显著提高了LoVo细胞G0~G1期的细胞比例,减少了S 期的细胞比例,并且增加了LoVo 细胞的凋亡率。同时,Western blot 结果表明干扰SNHG22 可提高LoVo 细胞凋亡蛋白Caspase6 表达水平,降低细胞周期蛋白Cyclin H、CDK3 表达水平。这意味着干扰SNHG22 表达能够显著抑制结直肠癌的发生和发展。EPHA3基因定位于人类染色体3p11.2,EPHA3 蛋白首次被发现在B 淋巴白血病细胞表面,其结构高度保守。正常组织中EPHA3 蛋白呈微量表达,但其在多种实体瘤如胃癌、乳腺癌、胶质母细胞瘤等过度表达,其阳性表达率与肿瘤的血管形成、大小、分级、侵袭以及患者的生存率显著相关[13-14]。EPHA3高表达水平与结直肠癌的分化程度、淋巴结转移密切相关,过表达EPHA3 可促进结直肠上皮细胞的恶性转化,在结直肠上皮细胞中发挥促癌功能[15]。本研究在qRT-PCR和Western blot测定中,检测到干扰SNHG22 表达后,LoVo 细胞中EPHA3 基因表达水平显著降低,提示干扰SNHG22 抗结直肠癌的功能可能是通过下调EPHA3基因的表达实现。
综上所述,干扰SNHG22 表达通过下调EPHA3 基因表达,抑制结直肠癌LoVo 细胞周期进展,诱导LoVo 细胞的凋亡,SNHG22 可能成为结直肠癌的诊断标志物和分子治疗靶点。
利益冲突:所有作者声明无利益冲突。
