lncRNA-HCG11通过调控miR-144-3p/PRP11分子轴参与结直肠癌细胞增殖及转移的分子机制研究
2021-12-08于德水张晓艳
于德水 张晓艳
结直肠癌(CRC)发病率在所有恶性肿瘤中位列第三[1]。超过50%的结直肠癌患者最终会发展成远端转移。肿瘤细胞的侵袭和转移是导致结直肠癌患者治疗失败以及死亡的主要原因[2]。大量研究表明,lncRNAs参与调节肿瘤的增殖、侵袭与转移等生物学行为[3]。lnc-RNA 可作为内源竞争性 RNA(ceRNAs)或发挥miRNA海绵作用,在转录后水平调节miRNA下游靶蛋白的表达,进而促进或抑制肿瘤细胞的增殖、侵袭和迁移。其中,lncRNA-HCG11在肝癌和乳腺癌细胞中表达上调,与肿瘤细胞的增殖、侵袭和迁移相关[4]。研究发现,miR-144-3p在胶质母细胞瘤、骨髓瘤、肺腺癌、食管癌、前列腺癌等中发挥抑癌基因的作用,抑制肿瘤细胞增殖、侵袭和迁移[5]。富含脯氨酸蛋白(proline-rich protein 11,PRR11)是近年发现的一个与肿瘤发生发展均密切相关的新基因,在非小细胞肺癌、乳腺癌、胰腺癌以及卵巢癌等多种恶性肿瘤组织中高表达,促进肿瘤的增殖、侵袭、迁移以及上皮-间质转化等恶性生物学行为[6]。并且,在胰腺癌中,miR-144-3p靶向下调PRR11表达,抑制胰腺癌细胞PANC-1的增殖。本研究旨在探讨lncRNA-HCG11通过调控miR-144-3p/PRP11分子轴参与结直肠癌细胞增殖及转移的分子机制,寻找结直肠癌中调控癌细胞增殖迁移的新靶点和标志分子,为诊断和预后提供依据。
1 材料与方法
1.1 细胞、试剂与耗材
人结直肠癌细胞株(HCT116、SW480、Lovo)和人正常结直肠上皮细胞NCM460均购于中国科学院上海生命科学研究所细胞库。PMI-DMEM 培养基、胎牛血清、双抗(链霉素,青霉素)及胰酶均购自Gibco公司;Transwell小室购自Corning公司;Trizol试剂、逆转录试剂盒、LipofectamineTM2000转染试剂盒、BCA蛋白定量试剂盒均购自Invitrogen 公司;双荧光素报告基因试剂盒购自Omega 公司;miR-144-3p mimic/inhibitor/scramble及实验用到的siRNAs和引物序列由TransGen生物科技公司合成;SYBR Green q-PCR Master Mix试剂盒购自Sigma公司;HCG11、PRP11的过表达质粒购自上海吉凯生物公司;质粒提取试剂盒、RIPA裂解液、CCK-8试剂盒、ECL 发光试剂盒均购自碧云天生物科技公司;Western blot一抗(antibody-GADPH,antibody-PRP11)和二抗(羊抗兔)购自CST公司;自动酶标仪购自BIO-BRAD公司;电泳仪和凝胶成像系统购自Thermo Fisher Scientific公司。
1.2 细胞培养和分组
人结直肠癌细胞株(HT29、LOVO、Lovo)用10%FBS、100 U/ml青霉素和100 μg/ml链霉素的DMEM细胞培养液,在37 ℃、5% CO2培养箱中培养。细胞分为si-HCG11组;pcDNA-HCG11组;miR-144-3p mimic组;pcDNA-PRP11组;pcDNA-lnc-HCG11+miR-148a-3p mimic组;miR-148a-3p mimic+pcDNA-PRP11组;si-lnc-HCG11+pcDNA-PRP11组。每组转染实验组设置NC对照。
1.3 细胞转染
选取对数生长期的结直肠癌细胞,胰酶消化,用DMEM培养基调整密度至1×105个/ml。接种到6孔板,每孔加2 ml细胞悬液,培养24 h,按照1.2中的分组设置,采用LipofectamineTM2000转染试剂盒转染,48 h后,荧光显微镜下观察转染效果。
1.4 qPCR检测结直肠癌细胞中HCG11和miR-144-3p的表达
Trizol 试剂提取人结直肠癌细胞株的总RNA,DNA酶处理后用逆转录试剂盒进行逆转录。取2 μl cDNA产物放入EP 管,采用SYBR Green Master Mix反应体系进行扩增。扩增体系总体积为 20 μl,反应条件为:95 °C 预变性 30 s,95 °C 变性 5 s、60 °C 退火、72 ℃ 延伸30 s,共循环 40 次。采用U6作内参,检测各组细胞HCG11和miR-144-3p相对表达。测定模板的Ct值,循环数法(2-ΔΔCt)定量相对表达量。见表1。

表1 PCR引物序列
1.5 CCK-8法检测细胞的增殖活力
胰酶消化对数生长期的各组细胞,96孔板接种细胞,接种量1×104个/孔,每组 设置3个复孔,每孔加100 μl培养基,分别在24 h、48 h、72 h、96 h后,更换新鲜培养基。检测前每孔加入20 μl CCK-8试剂,孵育1 h,用酶标仪测定450 nm波长处的吸光值(D450),绘制细胞增殖曲线图。相同条件下实验重复3次。
1.6 Western blotting检测细胞相关蛋白表达
收集细胞,用含有蛋白酶抑制剂的裂解缓冲液裂解细胞,用BCA蛋白浓度检测试剂盒测定蛋白浓度。50 μg/孔上样,10% SDS-PAGE凝胶分离1 h,转移至PVDF膜上,用含5%脱脂奶粉的PBS缓冲液封闭1 h,加入一抗,4 ℃过夜,PBST缓冲液漂洗3次,5 min/次,加入二抗,37 ℃ 孵育4 h。PBST缓冲液漂洗3次,加入ECL显影,再洗膜3次。使用 ECL 蛋白质印迹试剂盒显色。GAPDH用作内参,蛋白凝胶成像系统分析样品中目标蛋白的相对含量。实验重复3次,取平均值。
1.7 Transwell实验检测细胞的侵袭和迁移能力
侵袭实验中将 Matrigel 加入 Transwell 小室上室的聚碳酸酯滤膜上,37 ℃孵育15 min,使Matrigel胶凝固;迁移实验则无需 Matrigel 基质胶。将细胞用无血清培养基培养24 h后,用无血清培养基稀释细胞密度为1×105个/ml。在Transwell小室24孔板中,每孔上室加入200 μl细胞悬液,下室加入600 μl完全培养基,37 ℃、5% CO2培养48 h,用棉签拭去小室上方未穿膜的细胞,PBS上下冲洗2遍,膜下细胞用5%的多聚甲醛固定,0.5% 结晶紫溶液染色 10 min,显微镜下随机选取10个视野检查(200×),对细胞进行记数。相同实验条件下重复3次。
1.8 双荧光素酶报告基因检测lncRNA-HCG11与miR-144-3p、miR-144-3p与PRP11的相互作用
将lnc-HCG11与miR-144-3p、miR-144-3p与PRP11结合部位的序列及其突变体序列插入Firefly荧光素酶报告基因下游,构建表达载体。将miR-144-3p mimics与pGL4-HCG11-WT/MUT或pGL4-PRP11-WT/MUT共转染到HEK293T细胞。对照组用miR-144-3p scramble与pGL4-HCG11-WT/MUT或pGL4-PRP11-WT/MUT共转染。转染实验完成后,细胞继续培养48 h。裂解细胞,测荧光强度。Renilla荧光值作为内参。RUL值=Firefly luciferase RLU值/Renilla luciferase RUL值。
1.9 统计学处理
实验结果用统计软件SPSS 20.0分析,多组比较和两组间比较分别采用单因素方差分析和t检验,用GraphPad Prism 7对实验数据进行图片绘制。P<0.05或P<0.01表示差异具有统计学意义。
2 结果
2.1 结直肠癌和正常结直肠上皮细胞中lncRNA-HCG11的表达水平
qRT-PCR的检测结果表明,与正常结直肠上皮细胞NCM460相比,lncRNA-HCG11在3种结直肠癌细胞株(HCT116、SW480、Lovo)中的表达水平显著上调(P<0.01或P<0.001,图1)。3种结直肠癌细胞中,lncRNA-HCG11在Lovo中的表达水平最高(P<0.05或P<0.01,图1)。后续实验选取lncRNA-HCG11高表达的Lovo细胞。

注:与NCM460比较,**P<0.01,***P<0.001,与Lovo相比,#P<0.05,##P<0.01。
2.2 lncRNA-HCG11对结直肠癌细胞Lovo生物学行为的影响
qRT-PCR检测结果表明,与对照组相比,敲降lncRNA-HCG11,Lovo细胞中lncRNA-HCG11的表达水平显著降低(P<0.01,图2A);过表达lncRNA-HCG11,Lovo细胞中lncRNA-HCG11的表达水平明显著升高(P<0.001,图2A);CCK-8实验结果表明,敲降lncRNA-HCG11显著抑制Lovo细胞增殖活力(P<0.05或P<0.01,图2B)。Transwell实验结果表明,敲降lncRNA-HCG11,Lovo细胞的侵袭细胞数和迁移细胞数显著低于对照组(P<0.01,图2C-F)。而过表达lncRNA-HCG11,在CCK-8实验和Transwell实验中都得到相反的结果(P<0.05或P<0.01,图2B-F)。因此,敲降lncRNA-HCG11可抑制Lovo细胞的增殖侵袭和迁移;过表达lncRNA-HCG11则促进Lovo细胞的增殖、侵袭和转移。

注:与NC组比较,*P<0.05,**P<0.01,***P<0.001。
2.3 lncRNA-HCG11调控miR-144-3p的表达
starBase数据库的预测结果表明,miR-144-3p可能是lncRNA-HCG11的靶基因(图3A)。双荧光素酶报告基因实验结果表明,转染miR-144-3p mimics,lnc-HCG11野生型载体组HEK293T细胞的荧光强度与对照组相比显著降低(P<0.01,图3B),而lncRNA-HCG11突变型载体组细胞的荧光强度与对照组无显著差异(图3B)。这表明lncRNA-HCG11与miR-144-3p 靶向结合。同时,qRT-PCR定量结果表明,与对照组相比,敲降lncRNA-HCG11,Lovo细胞中miR-128-3p的表达水平明显升高(P<0.001,图3C)。由此可知,lncRNA-HCG11对miR-144-3p具有负调控作用。
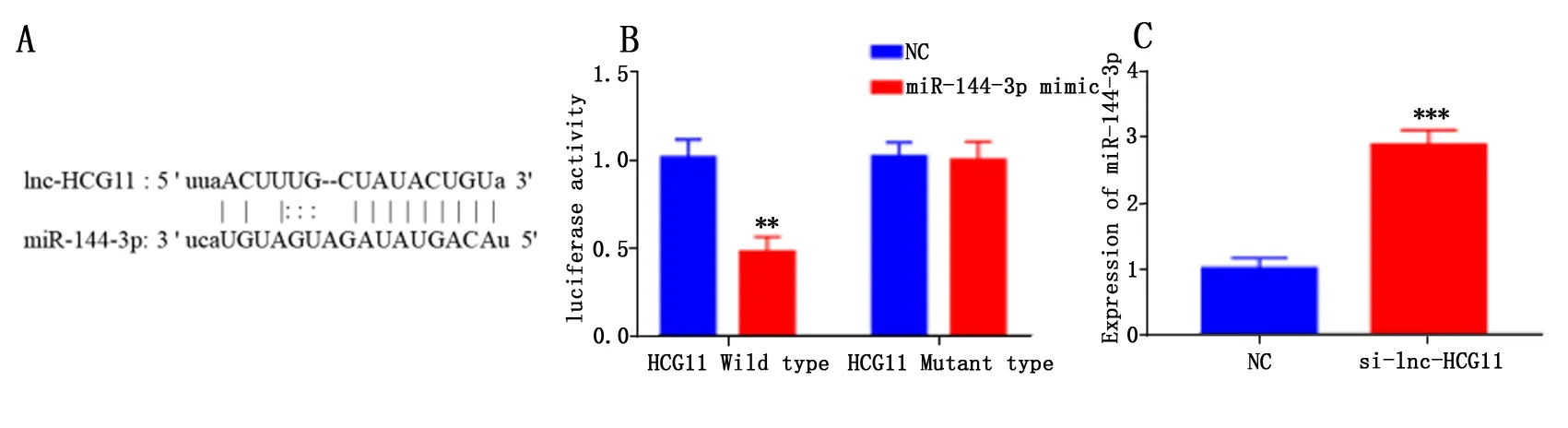
注:与 NC 组比较,**P<0.01,***P<0.001。
2.4 miR-144-3p对结直肠癌细胞Lovo生物学行为的影响
qRT-PCR检测结果表明,与对照组相比,过表达miR-144-3p,Lovo细胞中miR-144-3p的表达水平显著升高(P<0.001,图4A);CCK-8实验结果表明,过表达miR-144-3p显著抑制Lovo细胞增殖活力(P<0.05或P<0.01,图4B);Transwell实验结果表明,过表达miR-144-3p,Lovo细胞的侵袭和迁移能力与对照组相比显著降低(P<0.01,图C-F)。而转染miR-144-3p inhibitor显著降低miR-144-3p表达(P<0.01,图4A);在CCK-8实验和Transwell实验中都得到相反的结果(P<0.05或P<0.01,图B-F)。因此,过表达miR-144-3p显著抑制Lovo细胞的增殖、侵袭和迁移;抑制miR-144-3p表达能显著促进Lovo细胞的增殖、侵袭和迁移。
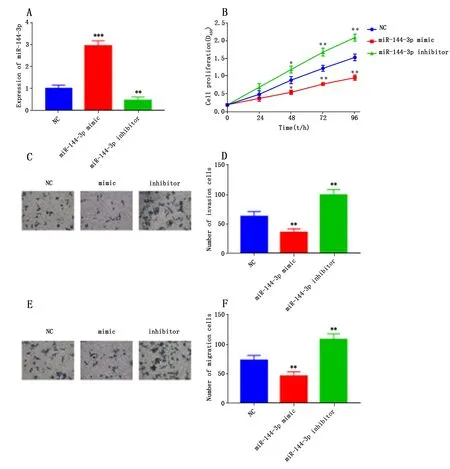
注:与 NC组比较,*P<0.05,**P<0.01,***P<0.001。
2.5 miR-144-3p对PRP11表达的调控作用
starBase数据库的预测结果表明,PRP11可能是miR-144-3p的靶基因(图5A)。双荧光素酶报告基因实验结果表明,转染miR-144-3p mimics,PRP11野生型载体组HEK293T细胞的荧光素酶活性显著下降(P<0.01,图5B),而miR-144-3p mimics与PRP11突变型载体共转染,HEK293T细胞的荧光素酶活性与对照组相比无显著差异(图5B)。同时,Western blotting检测结果表明,过表达miR-144-3p,Lovo细胞中PRP11的表达水平显著下降(P<0.01,图5C)。这表明,miR-144-3p负调控PRP11的表达。

注:与正常对照组比较,** P<0.01。
2.6 lncRNA-HCG11通过miR-144-3p/PRP11分子轴对结直肠癌细胞生物学行为的影响
Western blot检测结果表明,过表达PRP11,Lovo细胞中PRP11表达水平明显高于对照组(P<0.001,图6A);CCK-8检测结果表明,与对照组相比,过表达PRP11显著促进 Lovo细胞的增殖活力(P<0.05或P<0.01,图6B)。Transwell实验表明,与对照组相比,过表达PRP11组Lovo细胞的侵袭细胞数和迁移细胞数显著升高(P<0.01,图6C-D)。回复实验中,同时转染pcDNA-lnc-HCG11+miR-148a-3p mimic,miR-148a-3p mimic+pcDNA-PRP11,si-lnc-HCG11+pcDNA-PRP11,显著恢复了Lovo细胞中PRP11的表达水平(P<0.01,图6A)。CCK-8和Transwell实验检测结果分别表明,lnc-HCG11通过下调miR-144-3p对PRP11的抑制作用,促进Lovo细胞增殖、侵袭和迁移(P<0.05或P<0.01,图6B-D)。由此可知,lnc-HCG11与PRP11竞争结合miR-144-3p,促进Lovo细胞增殖、侵袭和迁移。
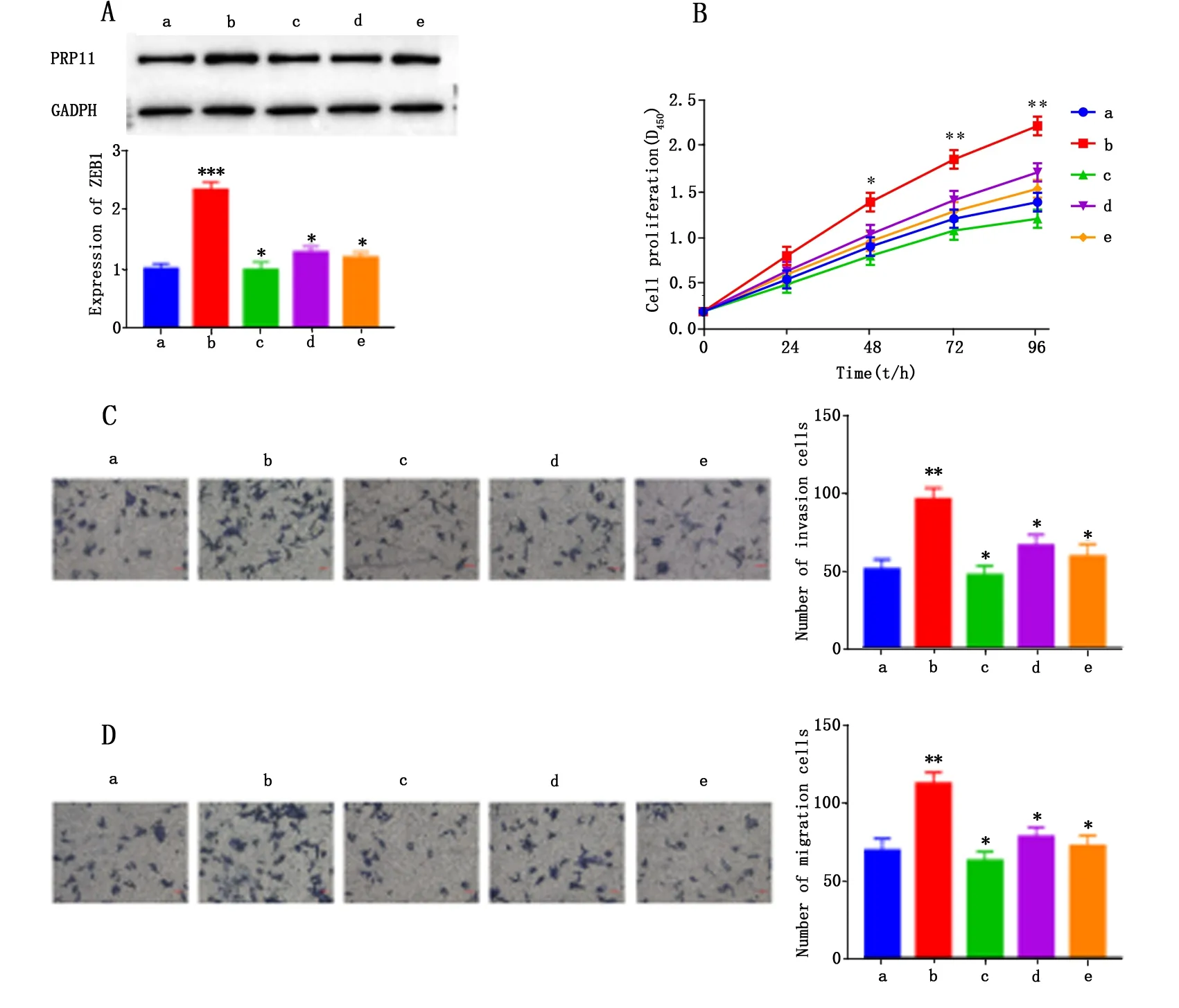
a:NC组;b:pcDNA-PRP11组;c:miR-144-3p模拟+pcDNA-PRP11组;d:si-lnc-HCG11+pcDNA-PRP11组;e:miR-144-3p模拟+pcDNA-PRP11组。与a组比较,*P<0.05,**P<0.01,***P<0.001,与b组比较,#P<0.01。
3 讨论
结直肠癌(colorectal cancer,CRC)是常见的消化道恶性肿瘤之一,在我国的发病率呈上升趋势。结直肠癌患者的5年存活率与肿瘤细胞的浸润、转移程度负相关。近年来结肠癌的诊断和治疗虽然在一定程度上降低了患者死亡率,但侵袭、转移仍是导致结肠癌患者预后差和治疗失败的主要原因[7]。因此,深入研究结直肠癌侵袭和转移相关的分子机制,对患者的诊断和预后有重要意义。大量研究证实,lncRNA在肿瘤的侵袭转移中发挥了重要调节作用,已发现多个lncRNA与结直肠癌的侵袭和转移相关[8]。例如,He等发现,lncRNA-CCAT1在结直肠癌组织中的表达水平显著高于癌旁组织,在结肠癌细胞SW620中过表达CCAT1可显著促进细胞增殖和侵袭[9]。Lu等发现,lncRNA-PANDAR在结直肠癌细胞中的表达水平显著高于正常结肠上皮细胞,在体外实验中,敲降lncRNA-PANDAR显著抑制结直肠癌细胞生长、迁移和侵袭。Xu等发现,lncRNA-MALAT1在结肠癌细胞HT29中,表达上调,沉默lncRNA-MALAT1显著抑制结直肠癌细胞的增殖、侵袭和迁移,促进细胞凋亡[10]。本研究发现,lncRNA-HCG11在3株结直肠癌细胞中的表达明显高于正常结肠上皮细胞;敲降lncRNA-HCG11显著抑制肠癌细胞Lovo的增殖、侵袭和迁移能力;双荧光基因报告实验表明,lncRNA-HCG11负调控miR-148a-3p。
研究证实,miRNAs可与靶基因mRNA 3'端非翻译区结合,促进靶基因mRNA降解或者抑制其翻译,在转录后水平调节其下游靶基因的表达,从而发挥促癌或抑癌基因的作用。有研究报道,miR-144-3p在多种肿瘤中异常表达,并参与调控肿瘤的增殖和转移。其中,HCMV阳性胶质母细胞瘤细胞中,miR-144-3p 通过靶向下调TOP2A的表达,抑制HCMV阳性胶质瘤的体外增殖、克隆形成和侵袭[11];前列腺癌细胞中,miR-144-3p 通过靶向下调CEP55的表达,抑制前列腺癌细胞的增殖、诱导细胞凋亡。本研究发现,过表达miR-144-3p显著抑制结直肠癌细胞Lovo的增殖、侵袭和迁移;进一步实验表明miR-128-3p靶向负调控PRP11的表达。
PRP11是近年发现的与肿瘤发生发展密切相关的调节分子,PRP11参与细胞周期的调控,可能通过推动细胞的S期向G2/M期转换,进而促进肿瘤细胞增殖。研究表明,PRP11在胃癌、卵巢癌、乳腺癌、胰腺癌等多种肿瘤组织中高表达,与患者的不良预后相关[12]。虽然已证实PRP11参与细胞周期调控,但其涉及的分子机制仍然未知。目前为止,PRR11 在结直肠癌中的相关研究尚未报道。本研究通过生物信息学预测发现PRP11可能是 miR-144-3p的靶基因,并且,miR-144-3p与lnc-HCG11的位点也可以结合PRP11。基于以上,我们以内源竞争内源性 RNA理论为基础,探讨了lnc-HCG11通过miR-144-3p/PRP11分子轴对结直肠癌细胞Lovo增殖侵袭和迁移的调控作用。
本研究结果表明,HCG11在结直肠癌细胞系Lovo中表达水平显著高于正常结直肠上皮细胞,LncRNA-HCG11通过与PRP11竞争结合miR-144-3p,从而促进结直肠癌细胞Lovo的增殖和迁移。
