基于MiSeq高通量测序技术不同地区奶豆腐细菌群落结构研究
2021-11-22张春林张苗苗杨甜甜郭壮肖石高侯强川
张春林,张苗苗,杨甜甜,郭壮,肖石高,侯强川
(1.内蒙古农业大学 生命科学学院,呼和浩特 010018;2.湖北文理学院食品科学技术学院,湖北 襄阳 441053;3.湖北澳利龙食品股份有限公司,湖北宜昌 444100)
0 引言
奶豆腐是一种以牛奶、马奶、羊奶为原料,经煮沸使乳清与乳酪分离,随后经过凝固、自然发酵等过程制作而成的传统发酵乳制品[1],因其形状与豆腐相似而得名。奶豆腐已有几千年的食用历史,深受我国北方蒙古、哈萨克等游牧民族的喜爱[2]。奶豆腐中富含蛋白质、氨基酸以及对人体有益的微量元素[3],具有提高免疫力、美容养颜等方面的作用。目前我国奶豆腐的消费较少,究其原因与当前奶豆腐主要以手工制作为主,尚未实现规模化生产,产品品质无法得到有效保障等有关。
作为一种代表性传统发酵乳制品,奶豆腐中有着丰富的微生物群系,其在产品发酵过程中起着至关重要的作用,其中细菌对于奶豆腐风味及感官品质的形成发挥重要作用。例如乳酸菌通过产生大量的酸性物质[4],有利于奶豆腐的凝固,同时提高了产品的营养和风味[5],而丙酸杆菌和哈夫尼菌等细菌的存在则有利于奶豆腐颜色的形成[6]。传统手工奶豆腐多为牧民自家制作,制作环境、生产工艺[7]、气候条件[8]等因素均会对奶豆腐微生物群落结构产生影响,进而导致不同地区手工制作的奶豆腐品质存在差异。因此,对奶豆腐细菌群落结构进行解析有助于揭示不同地区奶豆腐风味品质存在差异的潜在机制,促进奶豆腐后续的工业化生产。当前以Illumina MiSeq 高通量测序技术为代表的第二代测序技术具有通量大、准确度高等优点,能够全面解析微生物群落结构[9],目前已经应用于食品[10]、土壤[11]、水体[12]等领域的研究。
本研究从新疆塔城和内蒙古锡林郭勒两个代表性的奶豆腐制作地区各采集了10 份奶豆腐样品,采用Illumina MiSeq 高通量测序技术对其中的细菌群落结构进行解析,探究了两个地区奶豆腐中微生物群落结构的共性和差异,为奶豆腐的工业化生产及品质控制提供了基础数据。
1 材料与方法
1.1 材料与设备
1.1.1 材料
奶豆腐:采集自新疆塔城和内蒙古锡林郭勒地区各10 份,奶豆腐样品收集后置于自封袋中,依次编号为TC1~TC10和XM1~XM10,运回实验室中于-20 ℃暂存,用于后续分析。
DNeasy mericon Food Kit 提取试剂盒,德国QIA⁃GEN 公司;引物338F/806R,武汉天一辉远生物科技有限公司;Illumina MiSeq 测序配套试剂,美国Illumina公司。
1.1.2 设备
MiSeq PE250 高通量测序平台,美国Illumina 公司;R930 机架式服务器,美国DELL 公司;Veriti FAST梯度PCR 仪,美国ABI公司。
1.2 奶豆腐DNA 提取、PCR 扩增和高通量测序
将奶豆腐样品解冻完成后用无菌研钵将其磨碎置于无酶无菌离心管中,使用QIAGEN DNeasymeri⁃con Food Kit 试剂盒进行基因组DNA 提取。参照Wang 等人的研究方法[13],使用带不同barcode 的16S引物以16S rRNA V3~V4 区为靶点进行PCR 扩增,将经过纯度和完整性检测合格后的扩增产物进行Illu⁃mina MiSeq高通量测序。
1.3 生物信息学分析
根据引物中不同的barcode(核苷酸标签)将返回的序列分配给每个样品,使用QIIME 分析平台[14]将合格的序列依次 使用PyNAST[15]、UCLUST[16]、Chime⁃raSlayer 进行对齐、划分操作分类单元(operational tax⁃onomic units,OTU)以及去除含嵌合体OTU 的操作,然后使用Silva[17]、Greengene[18]和RDP[19]数据库进行序列比对以明确各OTU 的分类学地位,并对各样品菌群α多样性和不同地区样品菌群β多样性进行计算。
1.4 图形绘制与数据分析
使用R 软件(v3.6.2)对α多样性指数、相关性热图进行可视化,基于加权和非加权UniFrac 距离进行β多样性分析,使用Canoco(v4.5)软件进行冗余分析(Re⁃dundancy analysis,RDA),其他图均采用origin2017 进行绘制。
2 结果与分析
2.1 不同地区奶豆腐中细菌群落组成分析
本研究20 份奶豆腐样本中共测得序列647 963条,平均每份样本的测序量为32 398 条。依据97%相似度进行OTU划分后共得到4 760个非嵌合体OTU,平均每份样本OTU 数为238 个。基于OTU 的注释信息,本研究首先在门水平对不同地区奶豆腐中细菌群落结构进行了比较分析,结果如图1所示。
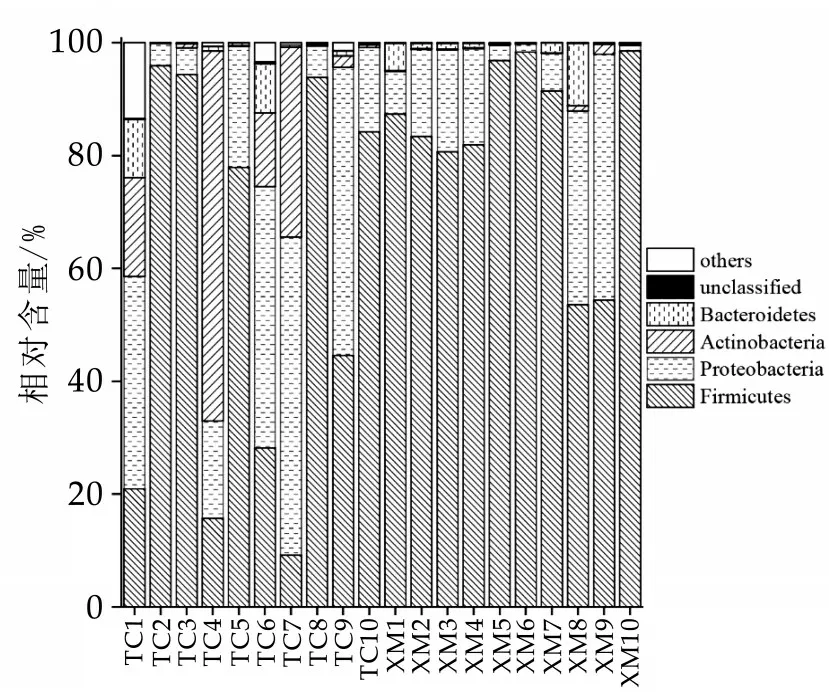
图1 不同地区奶豆腐中优势菌门的相对丰度
所有奶豆腐样本中共注释到32 个细菌门。由图1 可知,奶豆腐中的优势菌门(平均相对含量大于1.0%)共有4 个,分别为Firmicutes(硬壁菌门)、Proteo⁃bacteria(变形菌门)、Actinobacteria(放线菌门)和Bac⁃teroidetes(拟杆菌门),其平均相对含量分别为69.56%、20.35%、6.85%和2.19%,累计相对含量为98.95%。这与雷炎[20]等人之前的研究基本一致,不同之处在于上述研究发现奶豆腐中硬壁菌门和变形菌门的相对含量基本相同。显著性检验发现,塔城地区样品中放线菌门的相对含量显著高于锡林郭勒地区(P<0.05),显示出不同地区奶豆腐菌群群落结构存在一定差异。本研究进一步在属水平上对奶豆腐中细菌群落结构进行了比较分析,结果如图2所示。
由图2 可知,奶豆腐中含有12 个优势细菌属(平均相对含量大于1.0%),分别为Lactococcus(乳球菌属,28.52%)、Lactobacillus(乳杆菌属,26.77%)、Streptococcus(链球菌属,5.23%)、Acetobacter(醋杆菌属,4.98%)、An⁃oxybacillus(无氧芽胞杆菌属,3.57%)、Kocuria(考克氏菌属,2.87%)、Pseudochrobactrum(假苍白杆菌属,2.70%)、Arthrobacter(节杆菌属,1.93%)、Pseudomonas(假单胞菌属,1.76%)、Acinetobacter(不动杆菌属,1.52%)、Rouxiella(鲁希氏菌属,1.34%)和Ralstonia(罗尔斯顿菌属,1.18%)。可见奶豆腐中的优势菌属大多隶属于乳酸菌,且以乳球菌属和乳杆菌属为主。在优势菌属类群方面,本研究的结果与之前的报道基本一致[21],但不同菌属间相对含量存在明显差异。乳酸菌作为传统发酵乳制品中主要的微生物,不仅可以在奶豆腐发酵过程中分解乳糖产生乳酸,提升营养价值,还可以分解蛋白质形成具有特殊风味的物质,对于形成奶豆腐的特殊风味具有重要的作用[22]。例如,乳酸乳球菌可将牛奶中的乳糖发酵形成乳酸,进而产生风味物质双乙酰和乙醛[23],其胞内的肽酶和胞外蛋白酶在乳制品成熟过程中可促进蛋白质水解,对最终产品风味物质的形成具有重要作用[24]。此外,相关报道显示,乳杆菌属中的植物乳杆菌在发酵过程中可以产生一系列抑菌物质如抗菌肽、双乙酰、过氧化氢等[25],这对于提高机体免疫力和抑制病原菌具有积极的作用。
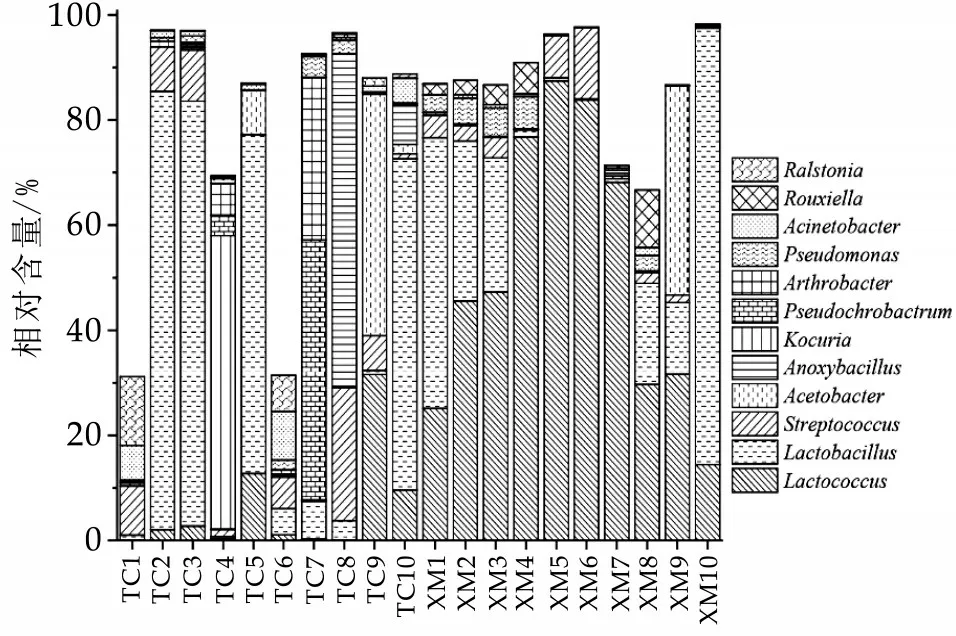
图2 不同地区奶豆腐中优势菌属的相对丰度
2.2 不同地区奶豆腐细菌群落结构α和β多样性分析
α多样性常用来评估样本中微生物的多样性情况,其常用的评估指标包括物种多样性指数(香农指数和辛普森指数)和物种丰富度指数(发现物种数和Chao1 指数)。本研究为探究不同地区奶豆腐微生物多样性的整体差异,对4 种不同的α多样性指数进行了比较分析,结果如图3所示。
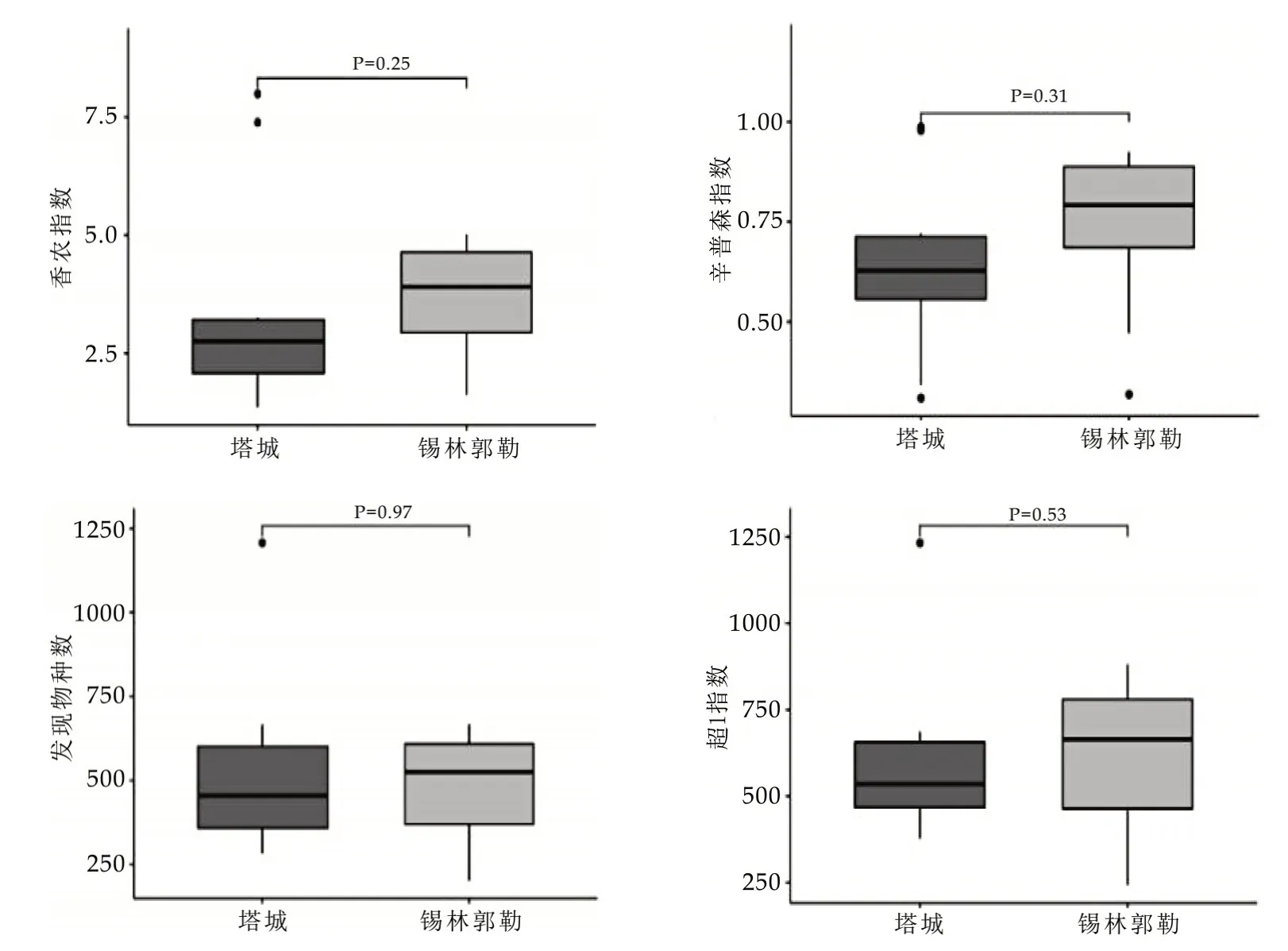
图3 不同地区奶豆腐中微生物α多样性的比较分析
由图3 可知,锡林郭勒地区奶豆腐中微生物在香农指数和辛普森指数上均略高于塔城地区奶豆腐,但所有α多样性指数差异均不显著(P>0.05)。可见,塔城和锡林郭勒地区奶豆腐中微生物的多样性和丰富度均无显著性差异。本研究在此基础上进一步对不同地区奶豆腐中微生物群落的整体结构进行了比较分析,结果如图4所示。
由图4(a)可知,在以前两个主成分绘图时,采集自锡林郭勒的奶豆腐样品分布较为集中,而塔城的奶豆腐样品在图中分布较为分散,同时两个地区的样品存在一定的交叠。由图4(b)可知,锡林郭勒和塔城奶豆腐沿第一主成分方向具有完全分离的趋势。上述结果表明,两个地区的奶豆腐样品在高丰度的菌群组成方面具有一定相似性,但是低丰度的菌群组成存在较大差异。同时,隶属于塔城的奶豆腐菌群群落结构差异大于锡林格勒地区奶豆腐,这为将来从中分离和收集乳酸菌菌株资源提供了宝贵的原材料。之前有研究发现奶豆腐中的微生物主要来源于环境[21],塔城位于欧亚大陆腹地,属于温带干旱和气候[26],而锡林郭勒为高原草场,属北温带大陆性气侯[27]。两者在气候上的差异会可能会导致环境中的微生物存在明显的不同[28],这可能是导致奶豆腐中微生物群落结构存在差异的最主要的原因之一。除此之外,不同牧民家庭在制作奶豆腐时挤压乳酪以分离乳清时所采用的力度可能不同,使得原始奶豆腐中的水分含量存在较大差异。由于塔城地区的气候更为干燥,乳酪中原始的水分含量对最终奶豆腐产品中菌群组成影响更为显著,导致该地区奶豆腐制品中菌群群落结构差异更大。

图4 基于加权(a)和非加权(b)UniFrac距离的主坐标分析
2.3 不同地区奶豆腐中差异菌属的甄别
为了进一步甄别导致两个地区奶豆腐中微生物群落结构差异的关键细菌属,本研究以不同采样地作为分组依据,以优势菌属为响应变量进行了RDA 分析,结果如图5所示。
由图5 可知,仅有乳球菌属和链球菌属共计2 个优势菌属与RDA 排序轴上的奶豆腐样本具有良好的赋值相关性。可见乳球菌属和链球菌属是造成两地奶豆腐微生物群落结构差异的关键细菌类群,其中乳球菌属在锡林郭勒奶豆腐中的相对含量较高,链球菌属在塔城奶豆腐中的相对含量较高。Li 等人使用乳酸乳球菌进行奶制品发酵时能提升酸奶中3-甲基丁醛和3-甲基-2-丁酮等挥发性香味物质的含量,对于提升酸奶风味品质具有重要的作用[29];而嗜热链球菌作为酸奶发酵常用的发酵剂之一,其在与保加利亚乳杆菌进行复配发酵酸奶时,对于奶制品品质的形成具有重要的作用[30]。此外,本研究对优势菌属在两地奶豆腐中的差异进行了统计分析,结果如6所示。
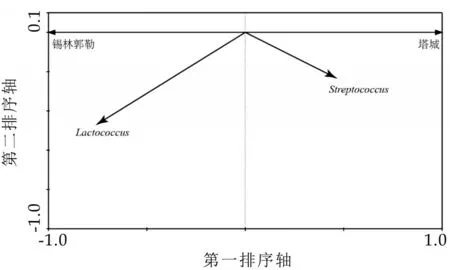
图5 RDA双序图
由图6 可知,12 个优势细菌属中有5 个菌属在两地奶豆腐中存在显著性差异。其中乳球菌属在锡林郭勒奶豆腐中的相对含量显著高于塔城奶豆腐(P<0.05),与RDA 结果一致。而假苍白杆菌属(Pseudo⁃chrobactrum)、节杆菌属(Arthrobacter)、不动杆菌属(Aci⁃netobacter)和罗尔斯顿菌属(Ralstonia)在塔城奶豆腐中的相对含量显著高于锡林郭勒奶豆腐(P<0.05)。乳球菌属的生长与周围环境中水分活度之间存在密切联系,Bassit[31]等人的研究发现,水分活度的降低会显著抑制乳酸乳球菌乳酸亚种diacetylactis 变种(Lactococ⁃cus lactis subsp.lactis var.diacetylactis,原名为Streptococcus diacetylactis)的生长。由于塔城身居欧亚大陆内部,导致该地区气候较锡林郭勒地区更为干燥,这可能是乳球菌属在锡林郭勒奶豆腐中的相对含量显著高于塔城奶豆腐的原因之一。同时,不同地区奶豆腐中菌群群落结构的差异进一步引起各地奶豆腐不同的产品风味和品质。


图6 不同地区奶豆腐中优势菌属的差异分析
3 结论
本研究采用Illumina MiSeq 高通量测序技术,对采自塔城和锡林郭勒地区奶豆腐中的微生物多样性进行了解析,发现不同地区奶豆腐中优势菌属的种类相似,但一些优势菌属相对含量及低丰度物种组成存在较大差异。不同地区奶豆腐中存在大量的乳酸菌类群,且主要以Lactococcus(乳球菌属)、Lactobacillus(乳杆菌属)和Streptococcus(链球菌属)为主,乳球菌属和乳杆菌属是造成塔城和锡林郭勒菌群结构差异的关键菌属,上述两个菌属可以作为后续评估不同地区奶豆腐品质差异的重点关注菌属。
