从江香猪miR-146b-3p 的组织表达、靶基因预测及其初步研究
2021-11-09崔小芝许厚强许家利谌颖莲倪萌萌
崔小芝,许厚强,*,熊 讯,陈 晨,许家利,谌颖莲,敖 政,倪萌萌,阮 涌,陈 伟
(1.贵州大学高原山地动物遗传育种与繁殖教育部重点实验室/贵州省动物遗传育种与繁殖重点实验室,贵州贵阳 550025;2.贵州大学动物科学学院,贵州贵阳 550025;3.贵州大学医学院,贵州贵阳 550025)
从江香猪(Sus scrofa)作为我国特有的地方猪种资源,具有肉质优良的种质特性;而大白猪(Sus scrofa)虽然在生长速度上具有优势,但其肉品质达不到消费者的要求[1]。从江香猪较大白猪生长迟缓,了解从江香猪生长迟缓的成因机制,对提高从江香猪养殖效率具有重要意义。因此,解析从江香猪生长缓慢的分子机制,是目前相关课题亟需攻克的难点。microRNA(miRNA)是一类在转录后水平调控基因表达的内源性非编码RNA,其大小长约18~25 个核苷酸,其通过与靶mRNA 的3'UTR 端区域结合来抑制靶基因的翻译或降解靶基因的表达[2],发挥其生物学功能。目前越来越多的研究发现miR-146b-3p 在细胞增殖、分化、迁移、侵袭、凋亡与肿瘤等生物过程中发挥重要的调控作用[3]。首先,在人体中miR-146b 基因位于10 号染色体q24-32,miR-146b-3p 和miR-146b-5p 是miR-146b 的不同成熟产物,miR-146b-3p 通过抑制靶基因NF2 促进甲状腺乳头状癌侵袭和转移[3]。而涂镇波[5]用肿瘤细胞功能学实验验证了超表达miR-146b-5p 能抑制急性T 淋巴细胞白血病细胞的侵袭与浸润,miR-146b-5p 还可调控树突状细胞的发育成熟和间充质干细胞成脂的分化[6]。其次,在探索博来霉素所致肺纤维化(Bleomycin-Induced Pulmonary Fibrosis)作用机制中,发现博来霉素诱导可以促进小鼠肺组织中miR-146b 的表达上调[7],miR-146b 上调后,还可通过降低细胞色素P450 芳香化酶(CytochromeP450-19A1,CYP19A1)促进猪卵巢颗粒细胞凋亡[8]。彭永东[9]还发现miR-146 能调控脂肪细胞分化和脂质代谢。此外,还有研究显示miR-146b-3p在鸡(Gallus gallus)生长发育过程中具有重要的调控作用,如miR-146b-3p 能直接抑制PI3K/AKT 通路和MDFIC,进一步抑制鸡成肌细胞的增殖与分化[10]。但miRNA 在不同物种之间具有高度的保守性[11],且miR-146b-3p 在猪生长发育过程中的作用机制尚不清楚。因此,本文以上述研究为基础,通过实验进一步探究miR-146b-3p 在猪生长发育中的作用机制,旨在为从江香猪分子遗传育种工作提供新的切入点。
1 材料与方法
1.1 实验材料 选取饲养环境相同的240 日龄健康从江香猪与大白猪各3 头,采集心脏、脾脏、肝脏、肺脏、肾脏、大肠、小肠和背最长肌8 个组织样,用0.9%的生理盐水、DEPC 水处理后,锡箔纸包装,做好标记,先放置液氮中,带回实验室转至-80℃冰箱保存。用于各组织RNA 与DNA 的提取。实验动物由贵州大学种猪场提供。
载体pMD-19-T 购自宝生物工程(大连)有限公司;PM-U6 微小miRNA 超表达载体由本实验团队构建(图1);大肠杆菌(Escherichia coli)DH5α感受态细胞、从江香猪肌细胞均由贵州大学高原山地动物遗传育种与繁殖教育部重点实验室保存。
图1 PM-U6 真核表达载体
1.2 试剂与仪器 DM 2000 Marker 购自擎科生物技术有限公司(北京);2×EsTaq Master Mix 购自康为世纪生物科技有限公司(北京);琼脂糖凝胶回收试剂盒、氯仿、异丙醇、无水乙醇、75%的酒精购自宏达尔生物科技有限公司(贵州);DNA 提取试剂盒Tissue DNA Kit、逆转录试剂盒购自赛默飞世尔生物科技有限公司(美国);质粒抽提试剂盒、Trizol 试剂购自英骏生物技术有限公司(Gibco,美国);超微量紫外分光光度计(型号为Thermo ManoDrop 2000,美国)、梯度PCR仪(型号为ABI VeritiTM,德国)。
1.3 实验方法
1.3.1 组织RNA 的提取 从-80℃冰箱中取出组织样品置于冰盒中,用已灭菌好的剪刀、镊子取50~150 mg组织于研钵中,加入液氮研磨组织成粉末,转移至新的1.5 mL 离心管中,根据Trizol 试剂盒说明书提取从江香猪和大白猪各组织的总RNA,对提取的总RNA 浓度进行测定后各取2 μg 总量,再根据逆转录试剂盒说明书进行逆转录获取cDNA。
1.3.2 DNA 的提取 组织块解冻,用生理盐水洗去血污,剪取约0.5 g 组织,放入1.5 mL 离心管中,剪碎,按照Tissue DNA Kit(Omega Biotek,美国)试剂盒说明书提取模板DNA,将所提取的DNA 储存在-20℃备用。
1.3.3 扩增引物的设计 根据miR-146b-3p 的序列,利用Primer 5.0 软件设计序列扩增引物和miR-146b-3p 颈环引物(表1),由生工生物工程(上海)股份有限公司合成。

表1 引物信息
1.3.4 miR-146b-3p 靶基因预测 本文采用miRNA22.2与TargetScan 靶基因在线软件预测miR-146b-3p 的靶基因(http://cbcsrv.watson.ibm.com/rna22_targets.html)。miR-146b-3p 序列来源于miRBase 数据库(http://www.mirbase.org/),miR-146b-3p 的成熟序列为:GCCCT GTGGACTCAGTTCTGGT。靶基因(Growth hormone receptor,GHR)3'UTR(GenBank NO:XM_01399063 6.2)序列来源于NCBI(https://www.ncbi.nlm.nih.gov/)。
1.3.5 实时荧光定量PCR(qRT-PCR)以逆转录获得的各组织cDNA 为模板,利用qRT-PCR 检测miR-146b-3p 在各样本中的相对表达情况,以GAPDH为内参基因,以大白猪心脏组织为对照样本,运用2-ΔΔCt[12]计算出miR-146b-3p 在不同样本中的相对表达量。qRTPCR 采用10 μL 反应体系:2×UltraSYBR Mixture(High ROX)5 μL、上下游引物(10 μmol/L)各0.3 μL、cDNA 0.5 μL、ddH2O 4 μL;反应程序:95 ℃预变性3 min,95℃变性30 s,57℃退火40 s,40 个循环;扩增反应结束后通过溶解曲线来判断PCR 反应的特异性。
1.3.6 miR-146b-3p 前体序列扩增及超表达载体的构建以提取的DNA 为模板对miR-146b-3p 前体序列进行扩增。PCR 扩增体系20 μL:Taq DNA 聚合 酶10 μL,上下游引物(10 pmol/ L)各1 μL,DNA 模板1 μL,ddH2O 7 μL。扩增程序:95℃预变性5 min;95℃变性30 s,57℃退火30 s,35 个循环;72℃补偿延伸10 min。对PCR 扩增产物以1.0%琼脂糖凝胶电泳进行检测。将片段大小相符的PCR 产物根据胶回收试剂盒说明书进行回收,回收产物与pMD-19-T 载体在16℃条件下连接16 h 后转化至大肠杆菌DH5α感受态细胞,挑取单菌落进行扩繁及PCR 检测,将阳性样本交送上海生工生物工程股份有限公司测序验证;T 克隆产物与PM-U6空载体利用SalI 和HandIII 双酶切,纯化后在16℃条件下连接16 h 后转化至大肠杆菌DH5α感受态细胞,挑取单菌落进行扩繁及PCR 检测,将PM-U6-miR-146b-3p 超表达阳性样本交送上海生工生物工程股份有限公司测序验证。
1.3.7 统计分析 通过2-ΔΔCt对qRT-PCR 数据进行计算分析,利用软件SPSS 19.0 对实验数据进行单因素方差分析,不同字母表示差异显著(P<0.05);*为差异显著(P<0.05);** 为差异极显著(P<0.01);样本重复数n=3。
2 结果
2.1 miR-146b-3p 在从江香猪及大白猪各组织中的表达分析 以大白猪的心脏组织为对照,利用qRT-PCR 检测到miR-146b-3p 在从江香猪和大白猪各个组织均有表达,结果如图2 所示。在从江香猪中,miR-146b-3p 在小肠中的表达量最高,极显著高于其余组织,其余组织表达量由高到低依次为大肠、肝脏、脾脏、心脏、肾脏、肺脏、背最长肌均有不同程度的表达,背最长肌表达量最低(P<0.01);在大白猪中,miR-146b-3p 在肺脏中表达量最高,在其他组织中的表达量由高到底依次为小肠、大肠、脾脏、背最长肌、肝脏、肾脏、心脏。miR-146b-3p 在从江香猪小肠、肝脏、脾脏、肾脏、肺脏和背最长肌中的表达量显著高于大白猪。
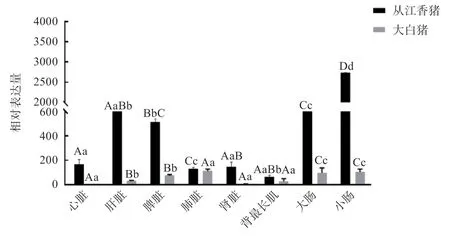
图2 miR-146b-3p 在从江香猪和大白猪不同组织中的表达
2.2 miR-146b-3p 靶基因预测 采用RNA22.2 在线软件,将猪GHR、IGF-1序列与miR-146b-3p 的序列进行预测分析。结果显示:GHR的3'UTR 序列中含有miR-146b-3p 的结合位点(图3),可能是miR-146b-3p 的候选靶基因,IGF-1的3'UTR 序列没有任何结合位点。

图3 miR-146b-3p 在候选靶基因中的靶位点
2.3 miR-146b-3p 克隆载体和重组质粒的构建与鉴定miR-146b-3p 克隆载体菌液PCR 鉴定结果(图4)与双酶切结果(图5)均显示条带为99 bp,测序结果(图6)显示与miRBase 数据库中的序列一致,没有突变位点,由此鉴定了miR-146b-3p 在从江香猪组织中存在。

图4 miR-146b-3p 前体序列的PCR 扩增

图5 PM-U6-miR-146b-3p 双酶切图
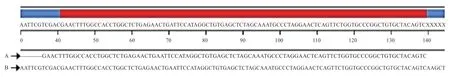
图6 PM-U6-miR-146b-3p 产物序列分析
2.4 miR-146b-3p 超表达载体转染从江香猪肌细胞 将miR-146b-3p 超表达载体转染至从江香猪肌细胞,48 h后提取细胞总RNA,在转录水平检测miR-146b-3p 及GHR、IGF-1的表达情况。qRT-PCR 检测结果表明(图7),miR-146b-3p 超表达载体转染从江香猪肌细胞后miR-146b-3p 的转录水平极显著上调,GHR的转录水平极显著下调,IGF-1也有所下调但差异不显著。
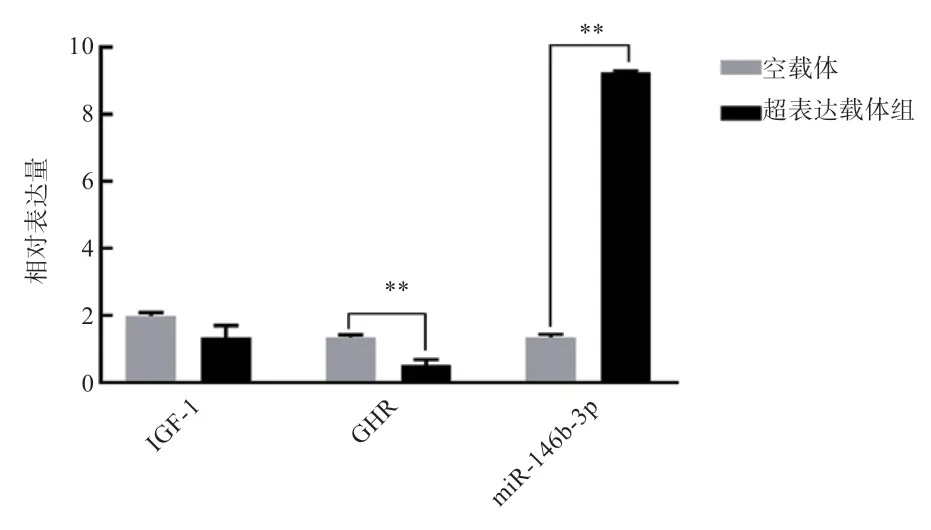
图7 miR-146b-3p 在从江香猪肌细胞中超表达后对GHR 基因表达的影响
3 讨论
miRNA 是一类在转录后水平调控基因表达的内源性非编码RNA。关于miRNA 在不同物种上的功能与机制研究取得了很多研究成果,但是关于猪miRNA 的研究以及其数据相对于人类miRNA 来说还有很大的探索空间。已有研究证实miR-146b-3p 参与细胞增殖分化,进一步调控动物的生长发育[13],但miR-146b-3p 在从江香猪生长发育过程中的作用机理尚不清楚。之前关于miR-146b-3p 都是在特定的组织与细胞中进行研究[14],miRNA 在不同物种和组织中的表达模式不同,功能也会随之不同[15]。构建miRNA 组织表达谱也是研究其功能的一种有力方法,其成本低、简单易行,且能够对miRNA 进行定量分析。因此,本实验首先检测miR-146b-3p 在各组织中的表达差异性,发现miR-146b-3p在从江香猪和大白猪心脏、肝脏、脾脏、肺脏、肾脏、大肠、小肠、背最长肌8 个组织中均有表达;miR-146b-3p 在从江香猪小肠和大肠中的表达量显著高于大白猪,这与miR-let-7a 在大肠和小肠均有高表达量一致,miR-let-7a 对猪在肠道吸收营养物质具有调控作用[16],这是否揭示miR-146b-3 也在猪肠道吸收营养过程有重要的作用还需要进一步验证。而miR-146b-3p 在从江香猪肝脏和脾脏中的表达量也高于大白猪,肝脏在动物体的生物学过程中具有非常重要的作用,如参与三大营养物质、维生素、激素等代谢过程,还参与分泌、排泄和生物转化等过程,脾脏是机体最大的免疫器官,且肝脏和脾脏会严重影响猪的采食量、生长速度、抗病能力、生产性能[17],miR-146b-3p 在从江香猪和大白猪中存在差异性,其可能是从江香猪和大白猪生长速度存在差异的调控基因。基于本课题组前期研究GHR在从江香猪与大白猪各组织中均有表达的基础[18],通过生物信息学分析发现miR-146b-3p 与GHR3'UTR 存在结合位点,而与IGF-1的3'UTR 没有结合位点。根据miRNA的竞争性作用机制[19],有类似的研究验证miR-139 与miRNA let-7b 能靶向负调控GHR3'UTR,进一步介导动物的生长发育[20],所以miR-146b-3p 也有可能通过结合GHR3'UTR 靶位点来抑制GHR的表达下调,进一步影响生长基因IGF-1的表达,从而导致从江香猪的生长速度低于大白猪。此外,GHR在大白猪心脏、肾脏、大肠、小肠、背最长肌、脂肪中的表达量均高于从江香猪,IGF-1在心脏、肝脏、脾脏、肺脏、肾脏、大肠、小肠、背最长肌中均显著高于从江香猪[21],而本研究miR-146b-3p 的表达量相反,在从江香猪各组织中的表达量均显著高于大白猪,进一步显示miR-146b-3p 可能是生长速度相关的差异基因。
此外,本实验将构建的miR-146b-3p 真核载体转染至从江香猪肌细胞中,从细胞分子水平检测miR-146b-3p 对GHR表达的影响,发现miR-146b-3p 在从江香猪肌细胞中超表达后,能够使GHR的表达极显著下调,而IGF-1表达量下调不显著,这与Ouyang[13]发现miR-146b-3p 通过靶向结合GHR的3'UTR 使GHR下调的结果一致,表明miR-146b-3p 能够通过影响生长相关GHR与IGF-1的表达来影响猪的生长。
综上,miR-146b-3p 在从江香猪生长发育过程中可能扮演着至关重要的角色,对进一步探究miR-146b-3p的功能及挖掘新的特色基因奠定基础,为解析从江香猪生长缓慢、个体矮小成因机制提供了理论支撑。
4 结论
除心脏与大肠外,miR-146b-3p 在从江香猪其余组织中的表达量均显著高于大白猪,并在从江香猪肌细胞中超表达后使GHR表达量极显著下降,揭示miR-146b-3p 可能对GHR存在负向调控作用,进一步调控猪生长发育。本研究为解析从江香猪生长缓慢与地方猪种的遗传改良提供了重要参考数据。
