昆虫口腔分泌物效应子及其在害虫防治中基于RNAi技术的应用展望
2021-11-06孙丽丽曹传旺
高 源, 孙丽丽, 曹传旺,2,*
(1. 东北林业大学林学院, 哈尔滨 150040; 2. 东北林业大学, 森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040)
昆虫与植物之间的斗争已有3.5亿多年的历史,目前有记载的昆虫种类达100万余种,其中50%以上为植食性昆虫(Wu and Baldwin, 2010)。昆虫对植物的成功定殖与其多样化的适应策略密切相关,口腔分泌物(oral secretion, OS)在其中起到了至关重要的作用(Consalesetal., 2012; Elzinga and Jander, 2013)。口腔分泌物,又称返吐液,是昆虫唾液腺分泌的唾液和肠道分泌物的混合液,其中含有多种调节植物激素反应的蛋白质,并在取食过程中与植物相互作用,从而影响植物直接防御和间接防御(殷海娣等, 2006; Chen, 2008; Hogenhout and Bos, 2011)。近20年来,RNA干扰(RNA interference, RNAi)成为研究者探究昆虫OS功能的主要手段之一。RNAi是研究昆虫基因功能的有价值的反向遗传学工具,也是目前最有可能应用于害虫防治的新技术(Pitinoetal., 2011; 胡少茹等, 2019)。基于昆虫OS与植物的相互作用关系,通过RNAi技术可以筛选出昆虫OS中直接用于害虫防治的靶标基因。因此,本文主要综述了昆虫口腔分泌物对植物防御的影响、口腔分泌物效应子(effector)的功能以及基于RNAi技术口腔分泌物效应子在害虫防治中的可行性探索,并对以口腔分泌物效应子基因为靶标的RNAi生物防治的应用前景和今后的研究方向进行了展望。
1 昆虫口腔分泌物对寄主植物防御的影响
植食性昆虫OS伴随昆虫的取食分泌至植物中,且OS中含有影响植食性昆虫相关分子模式(herbivore associated molecular patterns, HAMPs)或植物防御反应的各种激发子(elicitor),然而它们在植物中引起的防御反应不尽相同,所涉及的生物学过程也有所差异(Schmelz, 2015)(图1)。
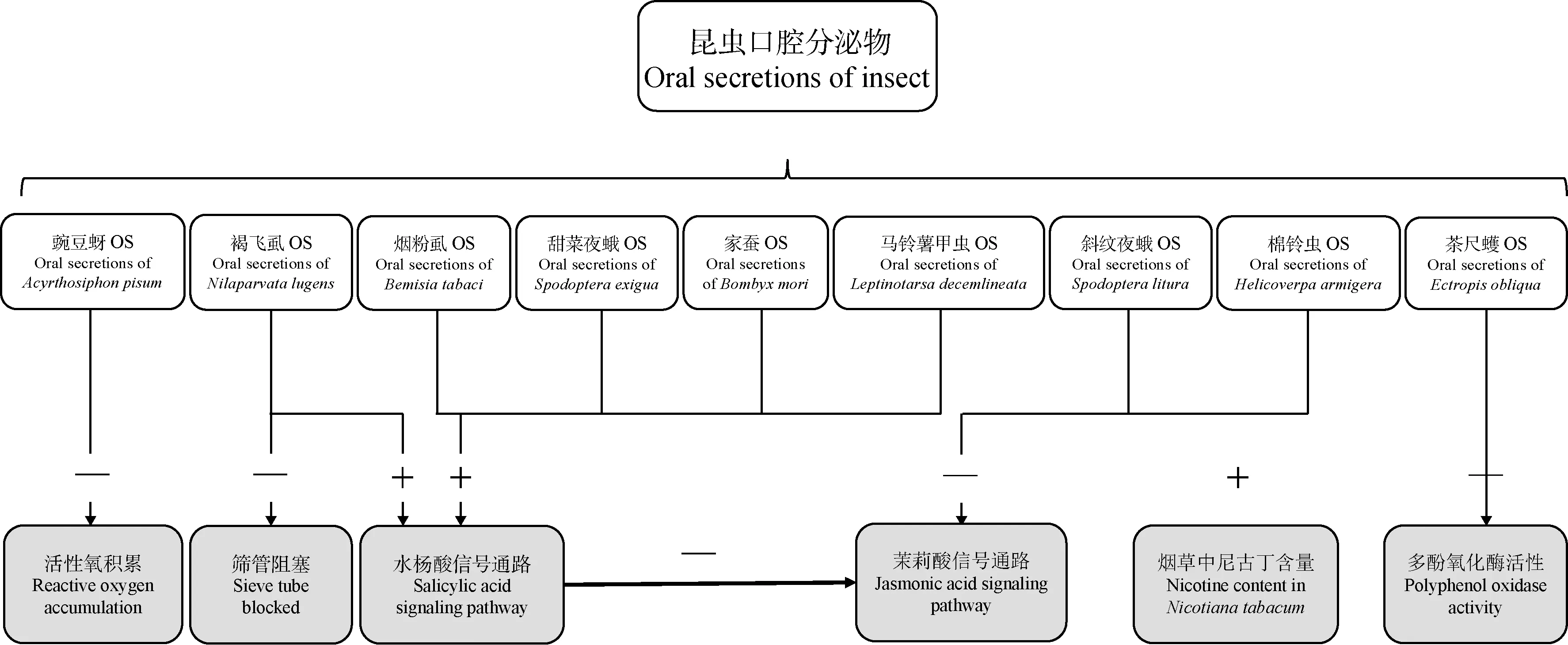
图1 昆虫口腔分泌物对植物防御反应的影响Fig. 1 Effects of insect oral secretions on plant defense responses+: 正调控植物的防御反应 Positive regulation of plant defense response; -: 负调控植物的防御反应 Negative regulation of plant defense response.
1.1 对寄主植物直接防御反应的影响
植物被机械损伤或被取食时均会发生一系列的防御反应,而膜电位(Vm)去极化和Ca2+升高的现象仅在昆虫OS与植物伤口接触时才会发生(Maffeietal., 2007)。Ca2+作为第二信使,细胞内Ca2+水平变化在介导应激反应中起关键作用,而钙调蛋白样蛋白(calmodulin-like proteins, CLMs)作为Ca2+的传感器会参与多种胁迫反应(Waretal., 2012)。拟南芥Arabidopsisthaliana钙调蛋白样蛋白CLM37和CML42只在OS的作用下才会表达,在拟南芥中抑制CLM42的表达可降低斜纹夜蛾Spodopteralitura的取食量,而抑制CLM37的表达可促进斜纹夜蛾取食,表明CLM42负调节拟南芥的防御反应;在植物对昆虫的防御反应中茉莉酸(jasmonic acid, JA)信号通路起到重要的作用,CML42则通过降低冠菌素不敏感1(coronatine insensitive 1, COI1)介导的JA敏感性及JA响应基因的表达而充当植物防御的负调节剂(Howe and Jander, 2008);而CLM37基因的表达会增强jasmonate-insensitive 1(JAR1)基因的转录,使茉莉酸-异亮氨酸(jasmonoyl-isoleucine, JA-Ile)含量升高,从而负调控JAZ(jasmonate ZIM-domain)家族基因JAZ10和JAZ12的表达量,表明Ca2+与JA信号通路有直接联系(Vadasseryetal., 2012; Sandraetal., 2014)。Verhage等(2011)发现菜青虫Pierisrapae幼虫取食拟南芥时,JA防御反应主要依赖于激活髓细胞组织增生蛋白2(myelocytomatosis protein 2, MYC2)等效应蛋白的表达,但在拟南芥机械损伤位置涂抹幼虫的OS,会优先启动JA信号通路的乙烯响应因子(ethylene-responsive factor, ERF)分支,说明菜青虫的OS可能存在激活ERF分支的效应蛋白,利用植物JA信号通路的MYC2分支会与ERF分支发生串扰作用,以抵抗植物的防御反应。在褐飞虱Nilaparvatalugens和棉铃虫Helicoverpaarmigera的OS中,也发现可以规避JA引起的植物防御反应的效应子(Jietal., 2017; Chenetal., 2019)。
众所周知,植物可通过筛管阻塞机制来防御蚜虫的摄食行为,而为了顺利获取食物,蚜虫唾液具有调控植物防御反应和降解植物次生物质的作用(尚哲明和刘德广, 2019)。蚜虫唾液由胶状唾液和水状唾液组成(严盈等, 2008),胶状唾液产生的寡半乳糖醛酸(oligogalacturonides, OG)作为激发子会触发植物Ca2+通路的防御,使Ca2+涌入筛元(sieve element, SE)中,发生筛管阻塞,阻止蚜虫进食。已有研究显示蚜虫的水状唾液中含有Ca2+结合蛋白,与Ca2+结合后抑制SE中的防御反应,但水状唾液仅短暂分泌至植物细胞中,因此OG诱导的植物防御不大可能被抑制(Willetal., 2007; Will and van Bel, 2008; Rüpingetal., 2010)。胞质Ca2+浓度的瞬时升高是植物感知环境的主要信号之一,蚜虫取食会被植物的共受体相关激酶BRI1-associated kinase 1(BAK1)识别,引起筛管的胼胝质沉积。而蚜虫唾液抑制筛管的阻塞是普遍存在的,通过对10种蚜虫取食试验,发现其中的7种蚜虫水状唾液蛋白的组成都不相同,均可抵消植物的筛管阻塞,但蚜虫唾液阻止胼胝质沉积的分子机理尚不清楚(Willetal., 2009; Vincentetal., 2017)。Ca2+结合蛋白普遍存在于昆虫OS中,如褐飞虱和烟粉虱Bemisiatabaci等,其沉默后会对昆虫的存活率或繁殖力产生影响(周涵宇, 2018; 李世香, 2019)。
来自口腔分泌物的不同化合物可被宿主植物识别,是诱导植物防御食草动物的激发子(Vadasseryetal., 2012)。在草地贪夜蛾Spodopterafrugiperda的OS中发现一种由植物叶绿体ATP酶γ亚基(cATPC)水解形成的二硫键桥接肽(+ICDINGVCVDA-),命名为inceptin,当其取食豇豆Vignaunguiculata时,摄入的cATPC在中肠中裂解而形成inceptins,少量分泌至豇豆中的受体蛋白可激活乙烯(ethylene, ET)、JA和水杨酸(salicylic acid, SA)防御通路(Schmelzetal., 2006)。在美洲沙漠蝗Schistocercaamericana中又鉴定到一种由饱和的和单不饱和的硫酸化α-羟基脂肪酸组成的化合物caeliferins,在蝗虫取食玉米后会诱导挥发性有机化合物(volatile organic compounds, VOCs)的释放(Albornetal., 2007)。
1.2 对寄主植物的间接防御反应的影响
昆虫OS可引起虫害诱导植物挥发物(herbivore-induced plant volatiles, HIPVs),吸引植食性昆虫的天敌,起到间接防御的作用(Danneretal., 2018)。含有18碳不饱和脂肪酸和L-谷氨酰胺(或L-谷氨酸)缀合物(fatty acid amino acid conjugates, FACs)作为OS的主要成分会诱导植物的防御反应。甜菜夜蛾Spodopteraexigua幼虫OS中分离出的N-(17-hydroxylinolenoyl)-L-glutamine是一种羟基FACs,命名为“挥发性诱导素”(volicitin),会引起玉米Zeamays释放出吸引天敌的挥发性化合物,在植物防御中起到正调控的作用(Albornetal., 1997)。挥发性诱导素由来源于植物的17-hydroxylinolenoyl与昆虫中肠合成的glutamine结合而产生,FACs水解为脂肪酸和谷氨酰胺并被昆虫的组织和血淋巴重新吸收,促进昆虫的氮代谢,但昆虫却无法控制FACs产生的威胁(Paréetal., 1998; 张一宾, 2007; Yoshinagaetal., 2008)。目前发现FACs存在于草地贪夜蛾、斜纹夜蛾、北京油葫芦Teleogryllusmitratus和黑腹果蝇Drosophilamelanogaster中(Spiteller and Boland, 2003; Spitelleretal., 2004; Yoshinagaetal., 2007)。
由于HAMPs具有化学复杂性和多样性,昆虫OS中还含有诱导其他植物反应的激发子 (Acevedoetal., 2015; Watermanetal., 2019),例如葡萄糖氧化酶(glucose oxidase, GOX)在很多鳞翅目昆虫OS中被鉴定,棉铃虫、斜纹夜蛾、烟夜蛾Helicoverpaassulta和烟芽夜蛾HeliothisvirescensOS中的GOX均可抑制烟草Nicotianatabacum中尼古丁和HIPVs的释放,并且棉铃虫幼虫OS中的GOX能够通过抑制烟草中的尼古丁而降低植物的防御反应(Zong and Wang, 2004; Musseretal., 2005; Delphiaetal., 2006; de la Paz Celorio-Manceraetal., 2011),而甜菜夜蛾和家蚕Bombyxmori等昆虫OS中的GOX则会通过引发SA和JA的串扰作用而降低植物防御(Diezeletal., 2009; 肖丽蓉, 2012)。此外,菜青虫和斜纹夜蛾的OS会降低拟南芥ethylene-responsive factor/APETALA2 transcription factor (ERF/AP2 TF)和蛋白酶抑制剂(protease inhibitor, PI)基因的表达,并减少叶片的水分流失,但与SA和JA通路无关(Weechetal., 2008; Consalesetal., 2012)。Kinoshita和Betsuyaku(2018)又通过模拟斜纹夜蛾对拟南芥的取食,发现斜纹夜蛾OS可抑制拟南芥伤口的愈合,并且拟南芥会转移营养物质使受损叶片组织衰老,有利于抵御虫害,而昆虫OS与植物衰老相关基因(senescence-associated genes, SAGs)的互作机理可作为今后的研究方向。
虽然昆虫口腔分泌物可引发植物对昆虫的间接防御,但一些昆虫在生物进化中为了更好地适应寄主,其OS会通过抑制HIPVs的释放来进行自我保护。绿叶挥发物(green leaf volatiles, GLVs)是HIPVs中一类重要的植物挥发物,机械损伤、昆虫取食、非生物胁迫等行为都会引起GLVs的释放,并具备排斥或吸引植食性昆虫天敌的功能(Matsui and Koeduka, 2016; Ameyeetal., 2018)。拟南芥受到机械损伤时会迅速合成并释放GLVs,而在伤口处涂抹菜青虫或甜菜夜蛾的OS时,会抑制氢过氧化物裂解酶(hydroperoxide lyase, HPL)相关基因的表达和GLVs的释放(Savchenkoetal., 2013)。家蚕OS在取食时从喷丝头位置分泌出来,通过消融喷丝头的方式使家蚕无法分泌OS,发现桑叶GLVs的释放量也随之减少,表明家蚕OS抑制GLVs的产生(Takaietal., 2018)。通过模拟昆虫取食,发现涂抹粉纹夜蛾Trichoplusiani、甜菜夜蛾、草地贪夜蛾和烟草天蛾ManducasextaOS的植物叶片与机械处理的植物叶片相比会减少GLVs的释放,从而减少植物对于昆虫的防御反应(Janesetal., 2019)。
1.3 共生微生物对植物防御的影响
昆虫OS中含有肠道的混合液,因此一些共生微生物在昆虫取食过程中也会与植物细胞接触,进而影响植物对昆虫的防御。Guo等(2013)研究发现斜纹夜蛾OS的共生细菌中含有孔蛋白样蛋白(porin-like protein, PLP),PLP会在植物细胞膜上形成通道而产生极化现象,跨膜离子通量发生变化,是植物对于昆虫防御的早期防御反应,并可能会激活SA和JA防御通路。Oliver等(2010)研究显示蚜虫内共生细菌不仅可为其提供必需的营养或维生素,还会对天敌起到防御作用。当植食性昆虫取食时,内共生细菌还会干扰植物的防御反应(Barretal., 2010)。Chung等(2013)发现科罗拉多马铃薯甲虫Leptinotarsadecemlineata幼虫OS中的内共生细菌能够诱导番茄SolanumlycopersicumSA的积累,从而干扰JA信号通路,使马铃薯甲虫的取食量升高;通过进一步对内共生细菌的分离,发现寡养单胞菌属Stenotrophomonas、假单胞菌属Pseudomonas和肠杆菌属Enterobacter都会影响番茄对马铃薯甲虫的防御反应,并鉴定假单胞菌属中纯化的鞭毛蛋白起主要调控作用。
大量研究证明,昆虫OS影响植物的防御,而有关昆虫OS效应子的功能知之甚少(De Vos and Jander, 2009)。功能基因组学技术的发展为深入研究昆虫OS效应子种类和功能提供了有效的方法,特别是第一个唾液蛋白C002的鉴定(Muttietal., 2008),让我们了解到蚜虫、粉虱等许多昆虫OS效应子均能诱导植物防御系统。随着关于植食性昆虫OS效应子及其功能的广泛而深入研究,将会进一步揭示昆虫OS效应子与植物防御系统相互作用的分子机理。
2 昆虫口腔分泌物效应子的鉴定及其功能
2.1 昆虫口腔分泌物效应子的鉴定
越来越多的口腔分泌物蛋白被归类为效应子,并且不同物种的OS效应子有所不同,表明昆虫已进化出针对不同宿主植物取食的策略(Will and van Bel, 2008; Raoetal., 2019)。目前,唾液腺转录组(RNA-seq)或OS的蛋白质组学(LC-MS/MS)测序是鉴定昆虫OS效应子主要途径。昆虫唾液腺的转录组测序能对无参考基因组昆虫的OS效应子进行分析鉴定,而蛋白组学可以确定与植物直接相互作用的效应子,包括没有激活植物防御的无信号肽链或来源于微生物的蛋白质,例如:马铃薯长管蚜Macrosiphumeuphorbiae的OS含有内共生菌Buchneraaphidicola,并从中鉴定到一种诱导植物模式触发免疫(pattern-triggered immunity, PTI)的伴侣蛋白GroEL,进而增强了植物的防御能力(Chaudharyetal., 2014; 苗雨桐等, 2017; Huangetal., 2019)。
功能基因组学技术的发展推动了植食性昆虫OS效应子的研究,近年来已对豌豆蚜Acyrthosiphonpisum、桃蚜Myzuspersicae、麦二叉蚜Schizaphisgraminum、马铃薯长管蚜、禾谷缢管蚜Rhopalosiphumpadi、角倍蚜Schlechtendaliachinensis、褐飞虱、烟粉虱、白背飞虱Sogatellafurcifera、伊氏叶螨Tetranychusevansi、茶尺蠖Ectropisobliqua、棉铃虫等20多种昆虫的唾液腺或OS进行了分析,并从中鉴定出51种与植物相互作用的效应子(Harmeletal., 2008; Carolanetal., 2009, 2011; 张帅等, 2012; 杜琴, 2014; Nicholson and Puterka, 2014; Chaudharyetal., 2015; 马琳等, 2015; Thorpeetal., 2016; Boulainetal., 2018; Huangetal., 2018a, 2018b; 苗雨桐, 2018; Yangetal., 2018)(表1)。目前已报道的OS效应子多数来自刺吸式口器昆虫,并且在植物对蚜虫OS效应子的识别中,发现其作用机理与细菌效应蛋白的识别存在很多相似之处(Jones and Dang, 2006)。
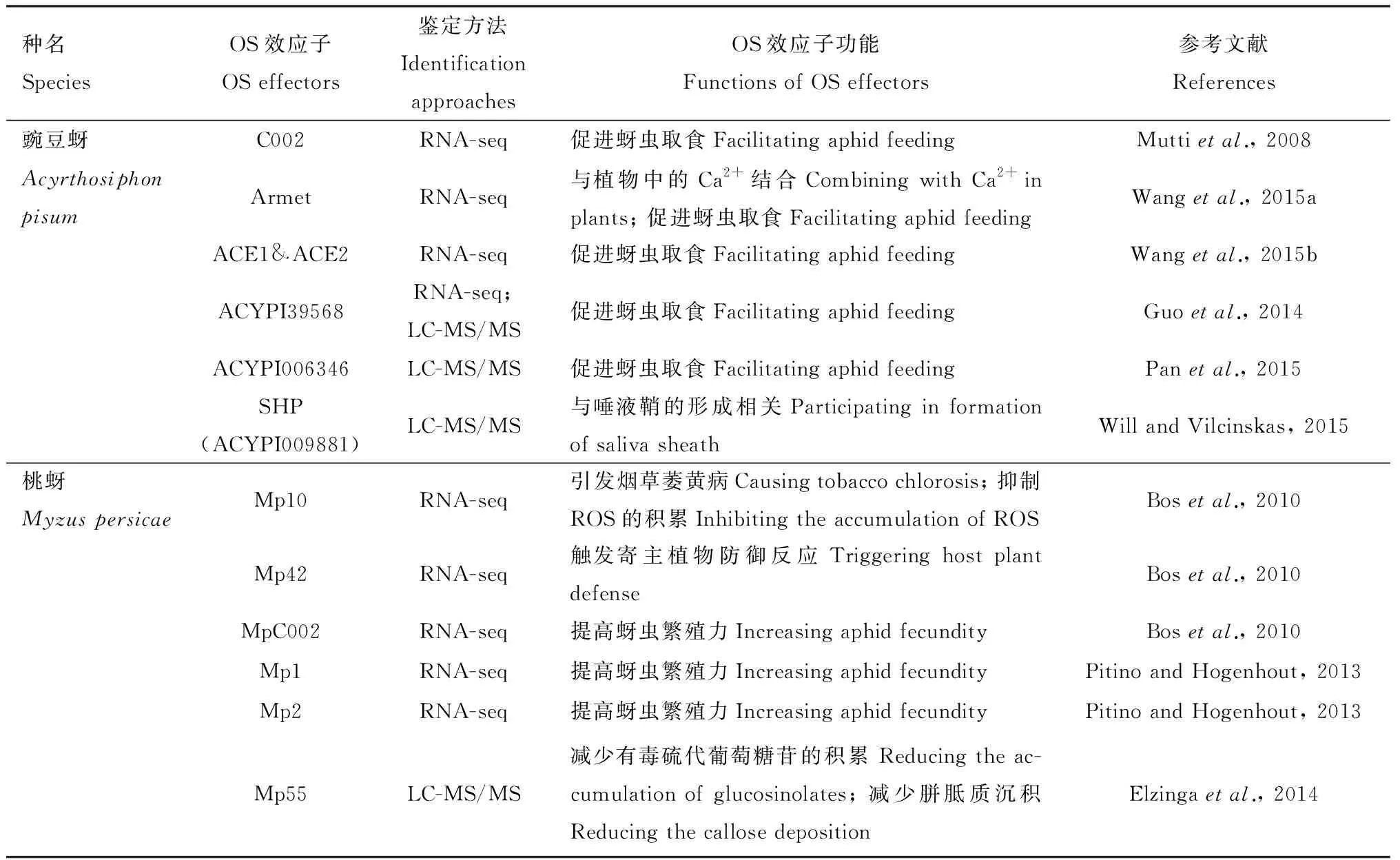
表1 已鉴定的昆虫口腔分泌物效应子Table 1 Identification of effectors in insect oral secretions

2.2 刺吸式口器昆虫口腔分泌物效应子的功能
2.2.1蚜虫口腔分泌物效应子:Mutti等(2008)在豌豆蚜唾液腺中发现的第一个OS效应子C002会随蚜虫取食分泌至宿主植物中,并在植物的韧皮部表达,表明C002是蚜虫取食过程中必需的蛋白。在桃蚜、麦长管蚜Sitobionavenae、麦二叉蚜、棉蚜Aphisgossypii和禾谷缢管蚜的基因组中也发现了ApC002的同源编码基因(Bosetal., 2010; 李雪峰等, 2014; Zhangetal., 2015; 任柯昱等, 2018; Escudero-Martinezetal., 2020)。蚜虫中C002的功能因物种而异,且效应子C002对于植物信号通路的影响尚不清楚(Pitino and Hogenhout, 2013)。除了C002,蚜虫OS中存在影响其摄食能力的其他效应子。在豌豆蚜的唾液中鉴定了一种蛋白酶——血管紧张素转换酶(angiotensin-converting enzyme, ACE),ApACE1和ApACE2基因在唾液腺中高表达,当豌豆蚜取食植物时,ACE1和ACE2会分泌至植物中并促进其取食(Wangetal., 2015b)。豌豆蚜OS中还具有一种Ca2+结合能力的水溶性蛋白Armet (arginine-rich, mutated in early stage of tumors),该蛋白的表达促进豌豆蚜取食(Wangetal., 2015a)。效应子Armet具有特异性,在烟草中过表达ApArmet基因会诱导SA的积累,这种SA积累仅对细菌病原体具有抵抗力,但不影响蚜虫的存活率和产卵量,这是由于蚜虫在摄食过程中误导植物抵御病原体,从而躲避防御(Cuietal., 2019)。虽然在其他蚜虫OS中也发现了ACE和Armet的同源蛋白,但这些效应子在蚜虫与植物相互作用中的功能尚不清楚(Zhangetal., 2017; 任柯昱等, 2018)。
桃蚜OS中鉴定了多种调节植物的防御反应的效应子。研究发现,桃蚜Mp10会诱导烟草局部叶片细胞死亡和萎黄,并抑制细菌flg22介导的活性氧(reactive oxygen species, ROS)反应,但蚜虫取食过表达Mp10转基因烟草会降低繁殖力,这种相互矛盾的结果说明Mp10可能会诱导效应子触发的免疫(effector-triggered immunity, ETI),从而降低蚜虫的性能(Bosetal., 2010; Rodriguezetal., 2014)。Elzinga等(2014)发现桃蚜效应子Mp55会减少植物中有毒硫代葡萄糖苷的积累,抑制胼胝质沉积,提高桃蚜的繁殖力,而Mp56, Mp57和Mp58会降低桃蚜的繁殖力。桃蚜效应子Mp1和Mp2对植物防御起负调控作用,进而提高桃蚜的繁殖力,并且在禾谷缢管蚜OS中也发现具有相同功能的直系同源效应子Rp1(Pitino and Hogenhout, 2013; Mugfordetal., 2016; Escudero-Martinezetal., 2020)。Rodriguez等(2017)进一步研究Mp1与寄主植物的互作关系,确定液泡蛋白分选关联蛋白52(vacuolar protein sorting associated protein 52, VPS52)是Mp1的作用靶标,为OS效应子在寄主植物中作用靶标的鉴定提供理论基础。
在马铃薯长管蚜OS效应子研究中,Me10和Me23的表达能够提高桃蚜在烟草上的繁殖力,但在寄主植物番茄中,只有Me10的表达提高蚜虫的繁殖能力(Atamianetal., 2013)。而效应子Me10与番茄防御信号的正调控因子14-3-3蛋白异构体7(tomato 14-3-3 isoform 7, TFT7)相互作用,说明蚜虫通过Me10干扰TFT7的功能,从而降低宿主植物对其防御能力(Chaudharyetal., 2019)。马铃薯长管蚜OS中还鉴定了一种谷胱甘肽-S-转移酶(glutathione S-transferases, GST)家族蛋白Me47,并通过植物的遗传转化发现Me47具有宿主依赖性(Kettles and Kaloshian, 2016)。蚜虫取食过程中除水状唾液的效应子会影响植物的防御反应外,胶状唾液中的一些效应子也会与植物防御反应进行对抗,使蚜虫顺利取食,例如桃蚜OS效应子Mp1(Will and van Bel, 2008; Pitino and Hogenhout, 2013)。蚜虫探针插入植物中会形成唾液凸缘和唾液鞘,唾液鞘则需要在有氧条件下形成,鞘蛋白(sheath protein, SHP)通过硫代基氧化形成的二硫键交联而形成(Willetal., 2012)。在豌豆蚜和麦长管蚜中均鉴定了一种参与唾液鞘形成的SHP,经验证,SHP在取食过程中发挥一定作用,但其对植物防御相关通路的作用尚不清楚(Raoetal., 2013; Abdellatefetal., 2015; Will and Vilcinskas, 2015)。
2.2.2褐飞虱口腔分泌物效应子:由于很多褐飞虱口腔分泌物效应子在与植物相互作用中的功能均未知,为探究褐飞虱唾液的成分和功能,人们利用功能基因组学对其效应子进行鉴定,研究表明为了适应不同宿主,不同种类的褐飞虱效应子也有一定的差异(Huangetal., 2017)。褐飞虱OS效应子NlANX-L5与典型的昆虫膜联蛋白表现出不同的结构域和表达特异性,但与含有EF-hand结构的钙调蛋白(calmodulin, CaM) NlCaM相似,推测这种效应子具有Ca2+结合能力;而另一效应子NlCA可能与CO2的水合和电离的活性相关,介导CO2的代谢(黄晓慧, 2016; Huangetal., 2016)。结合褐飞虱唾液腺的RNA-seq预测出一种具有Ca2+结合活性的唾液蛋白NlSEF1,NlSEF1表达会影响水稻中H2O2的含量从而介导水稻的防御反应(Yeetal., 2017)。在褐飞虱的OS中还鉴定一种编码内切β-1,4-葡聚糖酶(endo-β-1,4-glucanase)的基因NlEG1,NlEG1的表达能够降解水稻细胞壁中的纤维素使探针能够到达韧皮部,还可规避SA, JA以及JA-Ile引起的植物防御反应(Jietal., 2017)。
通过对褐飞虱胶状唾液效应子的分离鉴定,发现褐飞虱胶状唾液中最丰富的蛋白是黏蛋白样蛋白(mucin-like protein, Mul),效应子NlMul的表达能够保持唾液鞘润滑,并抵御植物的防御反应,但不参与唾液鞘的固化(Huangetal., 2017)。唾液蛋白Nlsalivap-3、鞘蛋白NlShp和粘蛋白样蛋白(mucin-like protein, MLP)NlMLP在唾液鞘的形成过程中均起到关键的作用,并且NlMLP在植物中表达会诱导植物细胞死亡、胼胝质沉积和JA等信号通路的防御反应(Huangetal., 2015, 2016; Shangguanetal., 2018)。
2.2.3其他刺吸式口器昆虫口腔分泌物效应子:除了对蚜虫和褐飞虱口腔分泌物效应子功能进行鉴定外,在其他刺吸式口器昆虫如黑尾稻叶蝉Nephotettixcincticeps和烟粉虱也进行了类似的研究,并取得了一定进展(Hattorietal., 2012; 邵若玄, 2017; 邵若玄等, 2018)。Hattori等(2012)在黑尾稻叶蝉OS中发现效应子NcSP84,功能分析显示NcSP84随昆虫取食分泌至植物筛管中,具有Ca2+结合能力,能够促进稻叶蝉连续取食,并推测此效应子会抑制SE的阻塞。效应子BtQ0F1和BtQ2477在烟草中的瞬时表达均会促进烟粉虱的存活,并发现BtQ2477干扰烟草的JA、SA和油菜素甾醇(brassinosteroid, BR)信号通路(洪越, 2016)。在烟粉虱OS中还鉴定了一种效应子Bt56,Bt56会通过SA-JA的串扰作用抑制烟草的防御反应,并确定Bt56在植物中的靶标NTH202转录因子,说明NTH202能够抑制烟粉虱诱导的SA累积(Xuetal., 2019)。
2.3 咀嚼式口器昆虫口腔分泌物效应子的功能
目前已鉴定的OS效应子大多数来源于刺吸式口器昆虫,而在咀嚼式口器昆虫中鉴定的效应子还比较少。杨子威(2013)发现茶尺蠖的OS可通过降低茶树中多酚氧化酶(polyphenol oxidases, PPOs)的活性而抑制植物的防御反应,但未鉴定到起作用的相关效应子。而茶尺蠖效应子EoGL虽然会抑制其生长,但在植物中的分子机理还尚不清楚(杜琴, 2014)。此前,虽已经对棉铃虫的唾液腺进行了RNA-seq分析,但尚未发现与寄主相互作用的效应子(张帅等, 2012)。最近,在棉铃虫OS中鉴定出一种类似于食肉昆虫毒R样蛋白的效应子,命名为H.armigeraR-like protein 1 (HARP1), HARP1效应子在植物细胞中的表达会下调JA通路相关基因的表达量,并与植物茉莉酸通路的主要负调控因子JAZ蛋白相互作用,从而阻止COI1-JAZ的相互作用和JAZ的降解,抑制植物JA防御反应(Chenetal., 2019)。HARP1的鉴定为咀嚼式口器昆虫OS效应子的研究奠定了坚实的基础,对今后昆虫OS的研究提供了新思路。
3 基于RNAi技术OS效应子在害虫防治中的可行性及应用前景
RNAi发展不仅促进昆虫基因功能和调控方面的研究,还为害虫的防治开辟了一条新途径(王聪等, 2018)。根据OS效应子的功能分析,在多种昆虫的OS效应子基因中鉴定到可以作为RNAi靶标的基因,并发现沉默OS效应子基因的表达会影响昆虫在寄主植物上的取食量、繁殖力和存活率等(胡少茹等, 2019)(表2)。昆虫OS效应子的RNAi研究中,主要通过显微注射法、饲喂法和喂食特定dsRNA转基因植物3种方式对昆虫靶标基因进行沉默,以探究OS效应子基因作为RNAi靶标在害虫防治中的可行性。

表2 昆虫口腔分泌物效应子在害虫防治中应用前景Table 2 Application prospects of effectors in insect oral secretions in pest control

续表2 Table 2 continued
3.1 显微注射法
在蚜虫的生物防治中,C002等效应子的基因均可作为靶标基因,Mutti等(2006)对豌豆蚜注射siC002,发现豌豆蚜无法在豌豆上摄食,但能够取食人工饲料。虽然无法确定C002的功能,但RNAi处理对豌豆蚜的存活率和摄食行为产生影响,说明C002在蚜虫的防治方面具有可应用性(Muttietal., 2008)。通过注射dsRNA的方法对豌豆蚜的ApACE1和ApACE2基因进行干扰,发现只有同时干扰两个基因时,豌豆蚜在植物上的存活率才会降低,而单独沉默某一基因不会影响其存活率(Wangetal., 2015b)。
唾液鞘结构的完整保证了蚜虫的正常取食,在豌豆蚜中注射dsApShp会完全破坏唾液鞘的形成,并降低豌豆蚜的繁殖率,但对存活率没有影响(Will and Vilcinskas, 2015)。褐飞虱的效应子NlShp, Nlsalivap-3和NlMLP与蚜虫SHP的功能相似,注射dsNlShp的褐飞虱无法形成唾液鞘,取食能力下降,并在取食4 d后出现死亡现象;沉默Nlsalivap-3的表达也会导致褐飞虱无法形成完整的唾液鞘,从而因取食异常导致蜜露排泄量显著减少,死亡率升高;而注射dsNlMLP的褐飞虱产生的唾液鞘短,大多数都无法到达韧皮部,探针位置产生的沉积物极少,进而影响褐飞虱在水稻韧皮部的取食(Huangetal., 2015, 2016; Shangguanetal., 2018)。沉默NlANX-L5,NlCA和NlCaM中的任何一个基因都会降低褐飞虱在植物上的存活率,并且沉默NlANX-L5的褐飞虱在韧皮部的摄食时间缩短,蜜露排泄量显著减少;注射dsNlCaM的褐飞虱表现出无法正常羽化、体色发黑等表型,并且褐飞虱的摄食能力受到影响,发育历期延长(黄晓慧, 2016; Huangetal., 2016)。在褐飞虱OS中还发现一种与唾液鞘形成相关的效应子NlMul,虽然沉默NlMul基因的表达不会影响唾液鞘的形状,但会导致唾液鞘变短且分支减少,影响褐飞虱的摄食和存活率(Huangetal., 2017)。褐飞虱NlEG1的表达会影响植物的防御,而沉默NlEG1的褐飞虱难以从韧皮部摄入汁液,在水稻植物上的取食量、存活率和繁殖能力也随之降低(Jietal., 2017)。同样,注射dsNlSEF1的褐飞虱会引起水稻更强烈的防御反应,并具有更高的死亡率和更低的摄食能力(Yeetal., 2017)。虽然褐飞虱的钙结合蛋白NlCBP1和NlCBP2与宿主的相互作用机理还尚不清楚,但通过注射dsRNA的方法沉默靶基因会降低褐飞虱的存活率(周涵宇, 2018)。
3.2 饲喂法
昆虫取食含有dsRNA的人工饲料与显微注射相比,该方法便于实际操作与应用,并且不会损伤昆虫。鳞翅目昆虫取食含有dsRNA的人工饲料时,dsRNA易被OS和血淋巴降解,导致沉默效率降低,而一些刺吸式昆虫在取食含有dsRNA的人工饲料后会使OS靶基因达到较好的沉默效果(Christiaensetal., 2014; 李晨雨等, 2021)。研究表明分泌蛋白漆酶1(laccase 1, LAC1)存在于烟粉虱及麦长管蚜OS中,当取食含有dsLAC1的蔗糖溶液时,烟粉虱及麦长管蚜的存活率均有所下降,说明LAC1具有克服植物抗性的作用(Yangetal., 2017)。烟粉虱OS中具有影响植物防御反应的效应子Bt56,通过取食含有dsBt56的人工饲料2 d,再取食烟草3 d,发现烟粉虱的存活率和繁殖能力均显著下降(Xuetal., 2019)。与Bt56相似,饲喂法沉默BtQ0F1基因也会导致烟粉虱的存活率降低(洪越, 2016)。
3.3 植物介导的昆虫RNAi
植物介导的昆虫RNAi也叫宿主诱导的基因沉默(host-induced gene silencing, HIGS),是指利用表达了昆虫特定基因的dsRNA的寄主植物喂食该昆虫,可以沉默昆虫特定靶基因的表达,并引起昆虫生长发育的异常(Nowaraetal., 2010; 傅淑等, 2019; Liuetal., 2020)。桃蚜的候选效应子MpC002, Mp1, Mp2和Mp55在植物中的表达都会提高桃蚜的繁殖力,然而通过农杆菌介导法将这几种效应子的dsRNA导入烟草或拟南芥中,发现MpC002, Mp2和Mp55通过植物介导的RNAi均会降低桃蚜的繁殖力,并且这种影响具有持久性,沉默效应稳定遗传给后代,大大降低桃蚜种群的增长;而Mp1通过植物介导的RNAi则不会对桃蚜产生影响,可能是由于Mp1与MpC002和Mp2在植物中的作用位点不同导致的(Pitinoetal., 2011; Elzingaetal., 2014; Colemanetal., 2015; Mugfordetal., 2016)。Mp1通过干扰VPS52的表达可能会降低植物对蚜虫的免疫力或负调控韧皮部的养分利用,因此,桃蚜取食过表达StVPS52转基因马铃薯降低其繁殖力(Rodriguezetal., 2017)。棉铃虫的效应子HARP1通过与JAZ蛋白相互作用进而干扰植物的JA防御反应,当棉铃虫取食35S:dsHARP1转基因拟南芥后,OS中的HARP1积累明显减少,并且在幼虫阶段表现出生长迟缓的现象(Chenetal., 2019)。
研究发现通过显微注射沉默豌豆蚜的ApShp基因会影响豌豆蚜的生长发育。Abdellatef等(2015)将SHP整合到大麦基因组中,获得dsSaShp转基因大麦;麦长管蚜取食转基因大麦能有效地沉默SaSHP的表达,并显现出唾液鞘变小,繁殖率降低,发育历期延长的现象,同时这种沉默效应连续遗传给后代。褐飞虱OS效应子NlMLP作为唾液鞘形成的调控因子,褐飞虱取食表达dsNlMLP的转基因水稻后,其存活率和取食量显著降低,说明以dsNlMLP转基因水稻为食会触发褐飞虱NlMLP基因沉默,并丧失机能(Shangguanetal., 2018)。
目前研究表明OS效应子在昆虫的生长发育过程中起关键作用,在不同物种中具有专一性,并且以OS效应子基因为靶标的HIGS可将这种RNAi效果传递至下代,能够产生类似的影响(Elzingaetal., 2014; Mugfordetal., 2016; Liuetal., 2020)。通过显微注射、饲喂和植物介导的RNAi的方法沉默昆虫OS靶基因的表达会影响昆虫的取食量、繁殖率和死亡率,由此说明以OS效应子基因作为靶标的RNAi技术在害虫防治中具有可行性和广阔的应用前景。
4 小结与展望
在昆虫取食过程中,昆虫的OS可通过多种方式干扰植物的防御反应,进而促进昆虫对寄主植物的取食。随着高通量测序技术的发展,多组学联合分析已成为当今研究热点。近期,Si等(2020)对棉铃虫和斜纹夜蛾的OS处理的棉花进行了转录组学和代谢组学分析,发现OS能够抑制棉花防御通路相关基因的表达并引起的棉酚含量升高。多组学的联合分析促进了昆虫OS与寄主植物在转录、蛋白和代谢水平中的作用机理研究更加深入,这也为今后昆虫OS效应子的功能研究提供了新方向。
目前,人们已经对蚜虫、粉虱、棉铃虫等多种昆虫的OS进行了功能基因组学分析,并确定部分OS效应子在昆虫取食中的功能,但与植物相互作用中,仅鉴定了Bt56和HARP1等少数几种OS效应子的作用机理。昆虫OS效应子在不同植物中的表达量不同,并且在不同龄期表达量也有所差异。很多OS效应子能够诱导植物相关防御通路标记基因表达,但防御激素却没有明显的变化,表明还有一些未知的OS蛋白的表达会影响植物的防御反应,这些未知的OS蛋白与寄主植物的相互作用机制有待深入研究。与以往OS效应子的研究不同,Guo等(2020)对烟草上饲养的烟草适应型蚜虫(tobacco-adapted, TA)和非烟草适应型(non-tobacco-adapted, NTA)蚜虫的OS进行分析,鉴定了一种在NTA蚜虫中上调表达的半胱氨酸组织蛋白酶Cathepsin B3(CathB3),CathB3与烟草EDR1(enhanced disease resistance 1-like)结合会触发ROS的积累,抑制蚜虫取食。CathB3的鉴定说明昆虫可通过控制OS效应子的表达提高对植物的适应性,而效应子在植物中的表达会降低昆虫对寄主的适应性,这为昆虫的生物防治提供了新思路。
目前害虫的防治主要以化学农药为主,化学农药的长期使用导致昆虫形成抗药性,大大降低了防治效果。RNAi技术应用在害虫防治中具有专一性,不会影响害虫天敌,解决了害虫对杀虫剂产生抗药性的问题。沉默昆虫中OS效应子的表达会影响昆虫的生长发育,并且可以传递至后代,具有稳定性。尽管目前的研究还仅停留在实验室阶段,但以MpC002,Mp2和HARP1等作为靶标基因的dsRNA转基因植物已经证实这种方法的可行性,说明OS效应子基因已具备作为RNAi靶标的条件,在害虫生物防治中具有良好应用前景,进而推动RNAi技术发展。
