双酚A(BPA)与黑腹果蝇雌激素相关受体(dERR)的结合模式及对其基因表达的影响
2021-11-06王利超李佳鹏郑相相廖艳凤欧阳霞辉
王利超, 李佳鹏, 郑相相, 王 娟, 廖艳凤, 欧阳霞辉
(西北民族大学生命科学与工程学院, 兰州 730030)
内分泌干扰化学物质(endocrine disrupting chemicals, EDCs)是指能够干扰生物体内天然激素的合成、分泌、运输、结合、作用和代谢的外源性物质或混合物(Atli, 2013),可以通过干扰脊椎动物和无脊椎动物的激素或核受体(nuclear receptor, NR)来干扰正常的激素系统(Canesi and Fabbri, 2015; Bovieretal., 2019)。EDCs可能会通过改变生理控制机制而对人类和环境健康产生不利影响(Elobeid and Allison, 2008)。双酚A(bisphenol A, BPA)是一种属于EDCs家族的非甾体异种雌激素(Nanjappaetal., 2014),作为一种工业用化学品,它通常用作聚碳酸酯合成中的单体,环氧树脂生产中的增塑剂(Koniecznaetal., 2015)。BPA在脊椎动物中起着致畸剂和内分泌干扰物的作用,高剂量(1~10 mg/L)具有致畸作用(Soneetal., 2004),低剂量(在μg/L范围)具有内分泌和多效性效应(Flintetal., 2012)。在毒理学和流行病学研究中表明,长期或高剂量暴露在BPA下,会影响人体肝脏与肾脏功能,同时对生殖、神经、免疫、代谢、心血管系统有潜在影响(Acconciaetal., 2015; Siracusaetal., 2018; Tassinarietal., 2020)。
雌激素相关受体(estrogen-related receptor, ERR)属于核受体(nuclear receptor, NR)超家族。在果蝇Drosophila基因组中发现了一个属于NR3亚家族的新的NR,这表明在后口和原口进化分裂之前就存在一个祖先,这种受体被称为果蝇雌激素相关受体(Drosophilaestrogen-related receptor, dERR)(Östbergetal., 2003)。目前对dERR的研究发现,dERR参与生物体脂类的合成和代谢、线粒体的生物发生及能量代谢的调控。果蝇幼虫发育特征是体重增加约200倍,这种増加需要高效地转换营养物质进入细胞,从而合成氨基酸、脂肪酸、核苷酸等,而正是dERR触发胚胎发生类似Warburg效应的代谢交换机制来支持幼虫生长发育(Tennessenetal., 2011)。迄今为止,虽然在脊椎动物及无脊椎动物ERR研究中均未发现其生理配体,但是仅含两个苯环的小分子BPA却成了ERR配体的最佳候选者。Bannister等(2013) 研究发现蜗牛ERR(MarisacornuarietisERR, mcERR)配体结合域(ligand binding domain, LBD)与BPA之间有弱相互作用。最近的研究也证明BPA与人类ERRγ紧密结合(Liu Xetal., 2014)。Grimaldi等(2019)证明BPA及其类似物可与ERRγ结合并激活其表达。目前对昆虫的研究表明,不同浓度EDCs家族成员能影响不同昆虫ERR基因的表达水平。Martínez-Paz 等(2017)认为目前在昆虫上研究的EDCs对昆虫有相似的影响,Liu T等(2014)认为EP能够模拟BPA的效应。低剂量的双酚A会影响脊椎动物中枢神经系统,这些作用的分子机制可能至少部分涉及BPA与ERRγ的高亲和力结合(Okadaetal., 2008)。同时BPA在果蝇某些发育障碍(如自闭症和注意力缺陷多动障碍)中可能起着致病作用(Kauretal., 2015)。但尚未评估BPA对dERR的影响。昆虫ERR与人类ERRγ直系同源(Jinetal., 2017),由此我们推测BPA可能与dERR存在类似ERRγ的特异性结合,且BPA可能通过dERR发挥其毒性作用。
分子动力学模拟方法由Alder和Wainwight于20世纪50年代提出,可以在分子及原子水平上观察到生物大分子行为(Dodsonetal., 2008),通过对分子、原子在一定时间内运动状态的模拟,以动态观点考察系统随时间演化的行为。本研究拟以黑腹果蝇Drosophilamelanogaster为研究对象,通过AutoDock Vina模拟小分子BPA与Modeller 9.25构建的dERR蛋白的分子对接,基于Gromacs进行分子动力学模拟并结合自由能的计算研究两者的结合模式,同时评估不同浓度BPA在不同时间段对dERR基因转录水平的影响,初步阐明BPA影响dERR的分子作用机理,为进一步研究昆虫ERR的功能及作用机制奠定基础。
1 材料与方法
1.1 黑腹果蝇的培养及处理
选用黑腹果蝇成虫及2龄幼虫将其饲养在培养箱中,温度为24±1℃,相对湿度为50%~60%,光周期为12L∶12D,饲喂标准的玉米粉果蝇培养基(蒸馏水180 mL, 玉米粉16.8 g, 蔗糖13.2 g, 琼脂粉1.5 g, 酵母粉1.5 g, 丙酸1 mL)。将1 mg纯度大于99.0%双酚A结晶(克拉马尔, 上海)固体化合物溶解于100 mL丙酮(>99.5%)中,取1 mL于10 mL容量瓶中逐级定容至10, 1, 0.1 μg/mL制备母液。取1 mL BPA母液,用5%的蔗糖溶液定容至1 L,用以制备BPA处理组所需原液(0.1, 1和10 μg/L)。研究表明丙酮在任何实验中都不会对生物体产生影响(Kwak and Lee, 2005),将1 mL丙酮用5%蔗糖溶液定容至1 L为对照组,处理组与对照组中丙酮浓度相同。黑腹果蝇成虫和2龄幼虫分别放置于含有BPA原液和丙酮对照的玉米粉果蝇培养基中,每隔6 h更换一次含相同浓度BPA原液及丙酮对照的果蝇培养基。
1.2 黑腹果蝇ERR与人类ERRγ氨基酸序列比对
昆虫ERR与人类ERRγ直系同源,为了确定dERR与ERRγ的保守性,基于在线多序列比对工具Clustal Omega(https:∥www.ebi.ac.uk/Tools/msa/clustalo/)对dERR和人类ERRγ氨基酸序列进行比对。采用Jalview展示两者完全一致的氨基酸。
1.3 黑腹果蝇dERR三维结构构建
基于NCBI蛋白质序列数据库中已有的dERR氨基酸序列,使用基本局部比对搜索工具(BLAST)从Protein Data Bank(PDB)数据库中搜索模板。采用Modeller 9.25以4NQA(PDB ID)的A链为模板对dERR蛋白进行同源性建模 (Webb and Sali, 2016),使用PDBsum(http:∥www.ebi.ac.uk/pdbsum)服务器对dERR结构质量进行PROCHECK分析(Peachetal., 2017),计算的Ramachandran图(Rose, 2019)对预测的dERR进行评估和验证。通过3Drefine (Bhattacharyaetal., 2016)改善蛋白结构,使用ModRefiner (Xu and Zhang, 2011)对侧链重新定位。
1.4 BPA与dERR分子对接
使用AutoDockTools(ADT)(版本1.5.6)(Morrisetal., 2009)将极性氢和部分电荷添加到dERR蛋白结构中。通过Discovery Studio Client 2020预测对接位点。小分子化合物BPA从PubChem数据库获取(CID: 6623)。采用AutoDock Vina进行BPA与dERR的分子对接,对接300次(Zahedietal., 2020)。最后将生成的结果直接加载到Discovery Studio Client 2020中进行相互作用力的分析,使用Pymol作图。
1.5 分子动力学模拟及结合能的计算
分子动力学模拟使用Gromacs 5.1.9在内部超级计算平台上进行。采用SPC水模型,将dERR蛋白溶于立方盒中。AMBER99SB力场生成蛋白质拓扑和Ambertools生成配体拓扑。系统显示10个负电荷,因此,添加了10个Na+离子来中和核电荷。为了消除空间碰撞,系统受到最陡峭的能量最小化,使最大作用力低于1 000 kJ·mol·nm。采用粒子网格Ewald方法计算了长程静电力。采用网格法确定邻居列表。在能量最小化后,分别在NVT(固定粒子数、体积和温度)和NPT(固定粒子数、压力和温度)条件下进行了5 000步(每陡峭2 fs)的位置约束模拟。最后,进行了6 ns的MD模拟。用Gromacs均方根偏差(root mean squared deviation, RMSD)和均方根波动(root mean square fluctuation, RMSF)工具计算评价蛋白质结构的重要指标均方根偏差(RMSD)和均方根波动(RMSF)。 使用Gromacs 5.1.9模块的g_mmpbsa,根据轨道法计算dERR-BPA复合物MD轨迹的3-6 ns的结合自由能,结合自由能可以反映各能量分项对BPA亲和力的影响程度。基于单个残基水平对结合自由能进行分解,进一步观察单个氨基酸结合能及其在对接中的贡献。数据可视化均采用R语言的Plot包实现。
1.6 基因表达量的qRT-PCR检测
收集1.1节处理组和对照组的黑腹果蝇成虫和2龄幼虫,采用RNAiso Plus试剂盒(TaKaRa, 大连)提取RNA,每组由3个重复组成(分别由3头成虫和5头幼虫组成)。采用PrimeScriptTM1st Strand cDNA Synthesis Kit (TaKaRa, 大连)合成cDNA。内参基因为Rp49,引物通过在线程序Primer-BLAST设计,由上海生工生物股份有限公司合成。dERR引物ERR-F: 5′-AGAAGGTGCTCAACTCGGAC-3′;ERR-R: 5′-GACATGTTCGTGCCTTGGATAA-3′。Rp49引物rp49-F: 5′-ATGACCATCCGCCCAGCATAC-3′;rp49-R: 5′-GCATCAGATACTGTCCCTTGAAGC-3′。qRT-PCR使用CFX96热循环仪(Bio-Rad, 美国)进行,每个样品设3个技术重复。qRT-PCR反应体系(10 μL): TB Green Premix Ex Taq(Tli RNase H Plus)(2×) 5 μL, 上下游引物(0.5 μmol/L)各0.75 μL, cDNA 1 μL, RNase-Free H2O 2.5 μL。反应程序: 95℃ 3min; 95℃ 5 s, 59.5℃ 15 s, 65℃延伸10 s, 35个循环。
1.7 数据分析
采用2-△△Ct法进行基因相对表达量计算,通过SPSS 22(IBM)软件进行单因素方差分析(ANOVA)(Inetal., 2020),采用t检验分析处理组和对照组数据间的差异显著性。使用GraphPad Prism8.3.0软件实现数据可视化。此外,实验组及对照组之间的相关性分析及数据可视化通过使用R语言(https:∥www.r-project.org/)和可公开获得的R包进行。
2 结果
2.1 黑腹果蝇dERR序列比对结果
采用Clustal Omega 对dERR和人类ERRγ氨基酸序列进行比对,结果显示氨基酸序列相似度为44.61%,一致度是57%(图1),可见两者在进化上是保守的。

图1 黑腹果蝇dERR与人类ERRγ氨基酸序列比对Fig. 1 Amino acid sequence alignment between ERR of Drosophila melanogaster and human ERRγ蛋白质来源物种及GenBank登录号Origin species of proteins and their GenBank accession numbers: 黑腹果蝇Drosophila melanogaster ERR, NP_729340.1; 人Homo sapiens ERRγ, NP_001127757.1.
2.2 黑腹果蝇dERR蛋白建模
采用Modeller 9.25,模拟得到1 000个dERR蛋白模型,并根据它们的离散优化势能(DOPE)得分进行排名,最终选用得分(-40 203.22656)最低的作为最佳模型(图2)。PDBsum计算dERR的Phi/Psi角的残基百分比(1.2%位于Ramachandran不允许区域,84.8%位于最受欢迎区域,1.6%位于慷慨允许区域)(图3)表明预测结构的立体化学质量很好,可进一步用于分子对接。

图2 黑腹果蝇dERR蛋白三级结构预测图Fig. 2 Prediction map of tertiary structure of dERR of Drosophila melanogasterLBD: 配体结合域Ligand binding domain; DBD: DNA结合域DNA-binding domain.

图3 黑腹果蝇dERR蛋白三维模型的Ramachandran图Fig. 3 Ramachandran plot of three-dimensional model of dERR protein of Drosophila melanogaster最受欢迎的区域、不允许的区域和允许的区域分别以红色、米白色和黄色着色,其中,只有1.2%的氨基酸残基位于Ramachandran不允许区域,84.8%位于最受欢迎的区域,1.6%位于慷慨允许区域。The most popular area, the disallowed area, and the allowed area are in red, off-white, and yellow, respectively, among which only 1.2% amino acid residues are located in Ramachandran disallowed areas, 84.8% in the most popular areas, and 1.6% in generously allowed areas.
2.3 BPA与dERR对接
采用Discovery Studio Client 2020预测对接位点,通过AutoDock Vina进行对接,结果显示(图4),BPA能够与dERR进行对接,能够完全进入dERR蛋白活性口袋。综合能量越低越稳定的原理及对接位点等因素,确定BPA与dERR结合最有可能的构象最优对接分数为-8.2 kcal/mol,下线偏差(rmsd u. b.)及上限偏差均为(rmsd l. b.)均为0;BPA和dERR之间结合不存在氢键作用力,而BPA的作用靶点与侧链氨基酸残基Leu338, Arg458和ILe289分别形成Pi-Alkyl键、Pi-Cation键和Alkyl键,键长分别为5.49, 2.96和4.07 Å。

图4 BPA与dERR 分子对接结果模拟图Fig. 4 Simulation diagram of molecular docking results of BPA and dERR图中Leu338, Arg458和ILe289残基分别用红色、绿色和蓝色表示,与之对应3种颜色虚线分别代表Pi-Alkyl, Pi-Cation和Alkyl键,淡紫色示小分子BPA。In the figure, the Leu338, Arg458, and ILe289 residues are represented by red, green and blue, respectively. The corresponding three color dotted lines represent Pi-Alkyl, Pi-Cation, and Alkyl bonds, respectively. Lavender indicates the small molecule BPA.
2.4 BPA-dERR分子动力学模拟及结合能
对BPA-dERR复合物进行了6 ns的MD模拟,运用主链原子均方根偏差(RMSD) 值监控体系的平衡,结果如图5(A)所示,dERR-BPA复合物和dERR蛋白单体变化趋势几乎一致,均在3 ns左右达到平衡,RMSD值在0.95 nm左右波动,这些数据表明BPA与dERR结合稳定,说明分子对接的结果是可靠的。哺乳动物ERRγ保持其活化构象,其中α-螺旋12必须位于配体结合口袋上的适当位置。BPA可以留在ERRγ-LBP (配体结合口袋)内,而无需更改此激活构象(Starovoytovetal., 2014)。可见我们的MD模拟结果是可靠的。BPA与脊椎动物和无脊椎动物的ERR作用方式可能是一样的,并不会改变ERR蛋白的构象而去影响ERR。在模拟过程中对根据蛋白质的骨架计算的蛋白质中每个残基的均方根波动(RMSF)进行了评估,以解决灵活性增加的问题,如图5(B)所示DNA结合域显示出较高的灵活性,而配体结合域二级结构元件显示出相对较小的偏差。BPA与dERR结合后可能通过影响dERR-DBD与雌激素相关受体反应元件(estrogen-related receptor response element, ERRE)的结合活性从而影响dERR的转录水平。g_mmpbsa计算的BPA与dERR的结合能为-66.799 kJ/mol,说明BPA与dERR能够有效和稳定的结合,同时计算静电作用能(-42.123 kJ/mol)、范德华能(-128.214 kJ/mol)、非极性溶剂化能(-13.786 kJ/mol)、极性溶剂化能(117.324 kJ/mol),可以看出范德华能为BPA的结合提供了主要驱动力。为定量获得在dERR-BPA复合物形成过程中关键能量贡献的具体细节,基于单个残基水平对结合自由能进行分解。通过极性溶剂化能我们发现Phe370(-5.0868 kJ/mol)及Leu334(-4.7107 kJ/mol)等侧链氨基酸是BPA和dERR两者结合的关键氨基酸。

图5 dERR-BPA复合物主链RMSD随时间变化曲线(A)和dERR蛋白残基随时间变化曲线(B)Fig. 5 Time-dependent curves of the RMSD in complex dERR-BPA (A) and the main protein residues of dERR (B)
2.5 3种浓度BPA对黑腹果蝇dERR基因表达影响及不同的时间和BPA浓度与dERR基因表达量间相关性
结果发现(图6),在6 h时,3种浓度BPA暴露使成虫及2龄幼虫dERR转录水平都发生下降,其中0.1 μg/L浓度显著下调成虫和2龄幼虫dERR基因表达(P<0.0001);在12 h时,3种浓度BPA暴露均使成虫及2龄幼虫dERR转录水平上调,其中0.1 μg/L浓度显著上调2龄幼虫和成虫dERR基因的表达(P<0.0001);在24 h时,3种浓度BPA暴露对成虫及2龄幼虫的dERR基因表达有不同的影响,其中0.1 μg/L处理后,2龄幼虫和成虫dERR基因表达几乎接近对照水平,1 μg/L处理后,2龄幼虫(P<0.001)和成虫(P<0.05)dERR基因表达均呈显著上升趋势,而10 μg/L处理后,成虫dERR基因表达呈显著上调趋势(P<0.05),而2龄幼虫呈显著下调趋势(P<0.05)。此次BPA浓度范围的实验结果通过皮尔逊相关性系数观察剂量和效应的关系,我们发现成虫和2龄幼虫dERR基因表达量在处理组及对照组的相关性是一致的(图7),其中6 h时,dERR基因表达量与3种BPA浓度间均呈正相关,仅12 h(1 μg/L)和24 h(10 μg/L)处理组分别与12和24 h对照组呈正相关,其余均呈负相关。

图6 不同浓度BPA处理不同时间对黑腹果蝇成虫(A)和2龄幼虫(B)dERR基因表达量Fig. 6 Expression levels of dERR gene in adults (A) and the 2nd instar larvae (B) of Drosophila melanogasterafter exposed to different concentrations of BPA for different time图中数据为平均值±标准误;柱上星号表示同一处理时间与对照有显著差异(*P<0.05, **P<0.01, ***P<0.001和****P<0.0001, t检验)。Data in the figure are mean±SE. Asterisks above bars indicate significant difference from the control at the same treatment time (*P<0.05, **P<0.01, ***P<0.001, and ****P<0.0001, t-test).
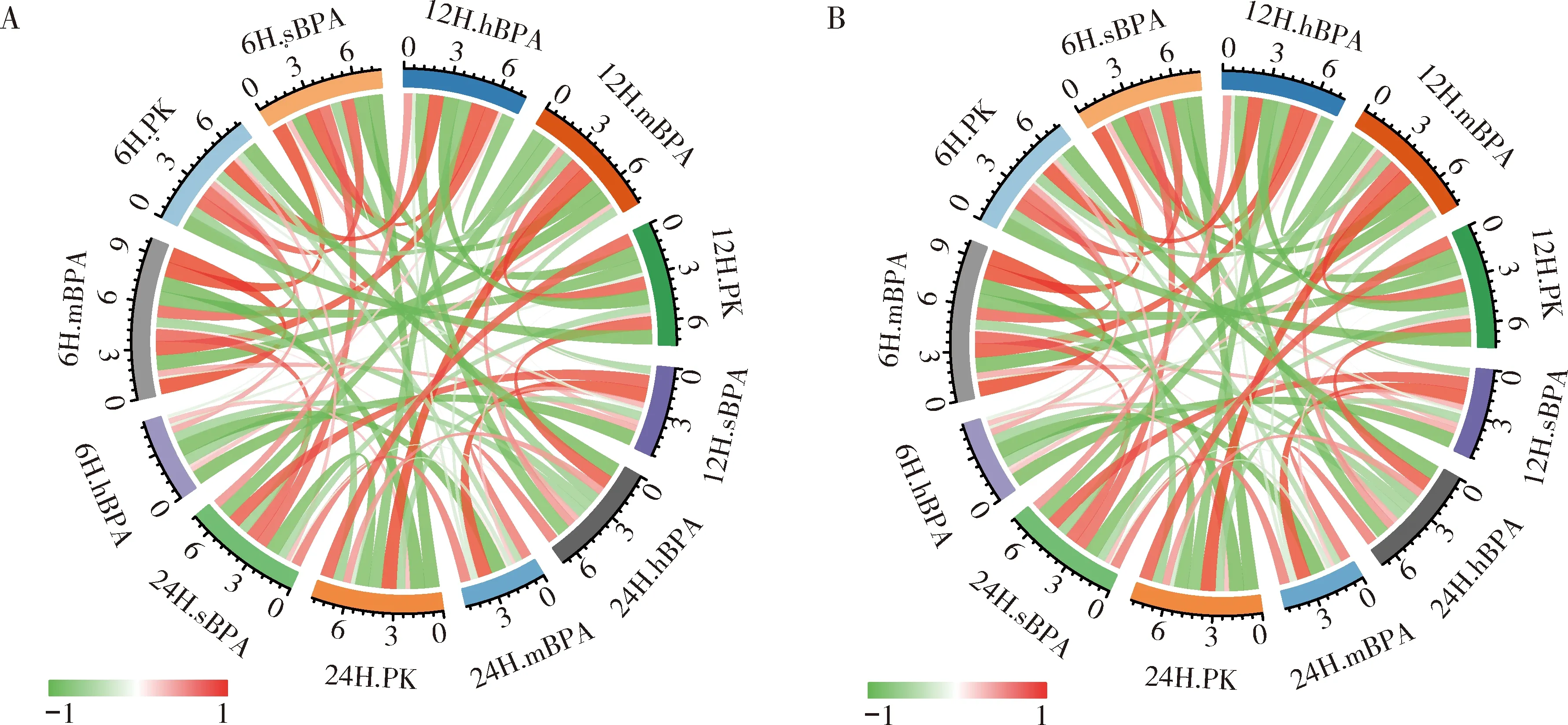
图7 黑腹果蝇成虫(A)和2龄幼虫(B)dERR基因表达量与不同的时间和BPA浓度的相关性圈图Fig. 7 Circle diagram of correlation of expression level of dERR gene in adults (A) and the 2nd instar larvae (B) of Drosophila melanogaster with BPA concentration6H.sBPA: 0.1 μg/L BPA处理6 h Treatment with 0.1 μg/L BPA for 6 h; 6H.mBPA: 1 μg/L BPA处理6 h Treatment with 1 μg/L BPA for 6 h; 6H.hBPA: 10 μg/L BPA处理6 h Treatment with 10 μg/L BPA for 6 h; 12H.sBPA: 0.1 μg/L BPA处理12 h Treatment with 0.1 μg/L BPA for 12 h; 12H.mBPA: 1 μg/L BPA处理12 h Treatment with 1 μg/L BPA for 12 h; 12H.hBPA: 10 μg/L BPA处理12 h Treatment with 10 μg/L BPA for 12 h; 24H.sBPA: 0.1 μg/L BPA处理24 h Treatment with 0.1 μg/L BPA for 24 h; 24H.mBPA: 1 μg/L BPA处理24 h Treatment with 1 μg/L BPA for 24 h; 24H.hBPA: 10 μg/L BPA处理24 h Treatment with 10 μg/L BPA for 24 h; 6H.PK: 丙酮处理6 h, 作为对照Treatment with acetone for 6 h as the control; 12H.PK: 丙酮处理12 h, 作为对照Treatment with acetone for 12 h as the control; 24H.PK: 丙酮处理24 h, 作为对照Treatment with acetone for 24 h as the control. 连线的颜色代表相关性(红色代表正相关,绿色代表负相关);连线的宽度代表相关系数。The color of line stands for correlation (positive correlation in red, and negative correlation in green) and the width of line represents the correlation coefficient.
3 讨论
本研究确定了dERR与人类ERRγ蛋白的同源性(图1);使用Modeller 9.25构建了dERR基于同源性的3D模型(图2),Phi/Psi角的残基百分比表明了预测的同源建模结构具有很好的立体化学质量;基于AutoDock Vina实现了BPA进入与(9cis)-视黄酸(模板4NQA的配体)一样的结合口袋。但是非生理配体小分子与蛋白质的相互作用不仅发生在活性口袋处,同时可能发生在蛋白质表面的其他口袋,它们的相互作用受能量匹配、空间匹配等多方面影响。其次,通过Discovery Studio Client 2020分析了两者的疏水相互作用、成键数目及键长,研究了两者的结合模式。有趣的是,人类ERRγ-LBD/BPA复合物的X射线晶体分析表明,ERRγ受体残基Leu342, Leu345, Asn346和ILe349(Liu Xetal., 2014)以及GLu275和Arg316(Okadaetal., 2008)充当BPA的固有结合位点。虽然dERR蛋白晶体结构仍未发现,但是从结果来看,ERR-LBD的Leu,ILe和Arg极可能是两者结合的关键氨基酸。此外,进一步从6 ns的轨道上对蛋白质-配体复合物进行了分子动力学模拟分析,结果显示dERR-BPA 复合物的结构较稳定(图4),说明分子对接的结果很可靠。利用g_mmpbsa定量获得复合物形成过程中关键能量贡献的具体细节,基于单个残基水平对结合自由能进行分解发现Phe370和Leu334等侧链氨基酸是BPA和dERR两者结合的关键氨基酸,在分子和原子水平上解释了两者相互作用的机制。EDCs在极高和极低剂量下可能会引发可观察到的效应,但在中等剂量下几乎没有效果(Flintetal., 2012)。为了进一步明确BPA对dERR基因表达的影响,我们测定了3个时间段(6, 12和24 h) 3种浓度BPA(0.1, 1和10 μg/L)对黑腹果蝇成虫和2龄幼虫dERR基因表达的影响,发现dERR基因表达呈现下调、上调或几乎接近对照表达水平的现象(图5),而且黑腹果蝇2龄幼虫对低剂量BPA较成虫更敏感。正如Flint等(2012)提到的一些无脊椎动物如昆虫幼虫可能对低剂量BPA的暴露更加敏感。
分子水平上的大量研究已经揭示了不同EDCs在不同程度上对昆虫ERR有不同的影响,但是无论是剂量效应还是作用机理方面都仍是知之甚少。这些研究中有3个推测观点几乎是得到认可:其一,EDCs直接或间接地与受体结合;其二,EDCs可能是一种蜕皮激素(20-hydroxyecdysone, 20E)模拟物,能够以非单一的方式影响ERR;其三,EDCs可能扰乱保幼激素(juvenile hormone, JH)和20E之间的平衡(Liu Tetal., 2014, Bovieretal., 2018)。在哺乳动物的研究中发现,BPA能够与ERRγ特异性结合(Liuetal., 2019)。有趣的是,昆虫ERR与人的ERRγ直系同源(Jinetal., 2017)。我们初步认为BPA影响dERR可能基于3点:其一是BPA与dERR有弱结合;其二是BPA可能是一种类固醇激素的模拟物,从而引起类似激素的生理效应;其三是BPA可能导致蜕皮类固醇激素紊乱,而这一点可能有ECR的参与。 奚耕思团队发现TeERR(TeleogryllusemmaERR)和TeEcR(TeleogryllusemmaEcR)彼此可以相互调控基因表达(Jinetal., 2017)。Martinez等(1991)早期的研究发现,序列基序5′-GGTCA-′3存在于无脊椎动物的ECRE中,实际上与雌激素相关受体反应元件(ERRE)基序5′-AGGTCA-′3非常相似。ERR是转录调节因子,能够与目标基因上游的回文反向重复序列5′GGTCAnnnTGACC-3′相互作用,调节转录起始速率。
基于不同浓度BPA处理6 h的结果,我们认为BPA在不改变dERR蛋白结构的情况下,与侧链氨基酸残基作用达到弱结合,与对应基因序列相互作用,调节自身转录起始速率,从而下调dERR基因表达水平。但这种影响不会一直持续。同时BPA可能造成蜕皮类固醇激素紊乱。随着时间的增加,当机体感受到这种胁迫后,20E的大量合成除了减弱了BPA对内激素的影响,进而可能通过ECR间接上调dERR基因表达。当然也不能排除低剂量BPA会被生物降解或代谢的因素(Staplesetal., 1998; Pritchettetal., 2002; Kangetal., 2007),如同非甾体药物的首过效应(Herman and Santos, 2021)。同时在哺乳动物的研究发现,双酚类物质(BPA, BPC和BPE)对NR的活性有不同程度的调节作用,ERRγ一般被双酚类物质激活(Thouennonetal., 2019)。BPA上调ERRγ mRNA和蛋白表达及其核定位是由ERK1/2磷酸化介导的(Hafezi and Abdel-Rahman, 2019)。因此,在果蝇中这种潜在作用可能存在。在不同浓度BPA处理12 h时,我们观察到dERR基因表达在幼虫和成虫中呈上调趋势。最终当机体恢复到初始状态后,dERR基因表达水平有接近对照水平的趋势。由于不同浓度的BPA及无法消除个体差异,我们在24 h也观察到不同的结果。
本研究首次阐述了BPA能够影响黑腹果蝇体内dERR基因的表达,通过由静态到动态的分子对接和分子动力学模拟为BPA影响dERR基因表达提供了可能的解释,初步阐明BPA影响dERR的分子作用机理,为昆虫ERR的研究奠定了一定的基础。
