亚甲蓝对机械性脑损伤小鼠的神经保护作用及其机制研究
2021-10-30赵红喜柳杨
赵红喜,柳杨
机械性脑损伤是一种常见的中枢神经系统损伤性疾病,随着工业、交通等领域的快速发展,机械性脑损伤发生率逐年增加。脑损伤的病理过程包括原发性损伤和继发性损伤,原发性损伤包括组织撕裂、细胞破碎及出血等,由原发性脑损伤引起的继发性脑损伤包括水肿、炎症反应、氧化应激及细胞坏死凋亡等。目前脑损伤的治疗均以减轻继发性脑损伤为主,目前临床大多使用阿司匹林等非甾体类抗炎药物或者激素类药物以减轻炎症反应,但由于毒副作用大,不适于长期服用。
亚甲蓝(methylene blue,MB)是一种吩噻嗪类化合物,水溶性强,分子式为CHNS·Cl⋅3HO,相对分子质量为373.90。亚甲蓝进入脑组织后,主要聚集于线粒体内部,从而调节氧化应激、炎症反应及细胞凋亡进程等。多项研究发现亚甲蓝对阿尔茨海默病、缺血性脑损伤及高原低氧所致脑损伤等神经系统疾病具有较好的治疗作用。也有研究表明亚甲蓝能够通过促进自噬改善创伤性脑损伤。但是对于脑损伤所引起的细胞凋亡及炎症反应的影响及其作用机制研究较少。核转录因子-κB(Nuclear Fctor-κB,NF-κB)是机体内具有调节炎症与免疫功能的细胞因子,能够激活促炎因子、影响细胞增殖、迁移及凋亡过程,能够促进白细胞介素(IL)-1β,IL-6 及肿瘤坏死因子α(TNF-α)的表达。本实验自2019 年3―8 月通过建立机械性脑损伤模型,探讨MB对机械性脑损伤小鼠的治疗作用与NFκB信号通路的关系。
1 材料与方法
1.1 药材及试剂
亚甲蓝、吡咯烷二硫代甲酸铵(pynolidine dithiocarbamate,PDTC)购自Sigma 公司,IL-1β、IL-6、TNF-α 试剂盒购自默沙克生物公司,小鼠抗小胶质细胞(Iba-1)、β-actin、IκB-α 抗体、兔抗NF-κB p65、p-IκB-α 抗体购自Cell Signaling Technol‑ogy(CST 公司),兔抗星形胶质细胞(GFAP)抗体购自北京博奥森,辣根过氧化物酶(HRP)标记羊抗小鼠、HRP 标记驴抗兔二抗购自北京鼎国,凋亡试剂盒购自南京凯基,全蛋白提取试剂盒、BCA 蛋白定量试剂盒购自碧云天生物。1.2 仪器
石蜡切片机(德国徕卡公司)、紫外可见分光光度计(深圳迈瑞)、酶标仪(深圳迈瑞)、倒置荧光显微镜(日本尼康公司),水平核酸电泳仪、半干式蛋白转膜仪(Bio-Rad)。1.3 实验动物与分组
SPF级C57BL/6雄性小鼠50只,体质量范围为20~25 g,购自辽宁长生生物有限公司[SCXK(辽):2013-0001],小鼠分笼饲养,自由饮食水,室温20~25 ℃,湿度50 %。本研究中对于小鼠的处理符合动物伦理学相关标准。小鼠适应性饲养1周后,采用随机数字表法分为以下五组:假手术组(Sham 组)、机械性脑损伤组(SWI 组)、亚甲蓝组(MB组)、NF-κB抑制剂组(PDTC组)、亚甲蓝联合NF-κB抑制剂组(MB+PDTC组),每组10只。1.4 机械性脑损伤小鼠模型的制作
利用10 %水合氯醛麻醉小鼠后,利用脑立体定位仪将其固定,酒精消毒皮肤、被毛,旋转定位仪旋钮至损伤位点:颅骨中线左侧2 mm、人字缝前2 mm。利用颅骨钻钻开一圆孔,将直径为1.0 mm 针头垂直插入脑组织中至2 mm 处,针头停留5 min 后拔出,止血,缝合伤口,消毒。将小鼠重新放于鼠笼中继续饲养。Sham组只破坏颅骨,不进行刺伤。手术1 h 后腹腔注射给予MB 1 mg/kg,每天一次;Sham 组给予等体积的蒸馏水;为了抑制NF-κB 信号通路活性,PDTC 组及MB+PDTC 组分别于造模前腹腔注射100 mg/kg PDTC溶液。1.5 小鼠神经功能评分评价
利用神经功能缺损评分(neuological severity scores,NSS)对各组小鼠于模型制备成功3、7、14 及21 d 后进行神经功能缺损评价,NSS 评分总分为18 分,正常为0 分,随着损伤严重程度增加,分数逐渐升高。1.6 TUNEL 检测细胞凋亡
于造模完成后7 d,将小鼠麻醉,心脏灌流后取脑,包埋,冰冻切片机上进行连续冠状切片,厚度为7 μm,利用原位末端转移酶标记(terminal deoxynucleotidyl transferase-mediat‑ed dUTP-biotin nick end labeling,TUNEL)试剂盒法检测小鼠脑内神经细胞凋亡情况,具体步骤按照试剂盒说明书进行操作。每组小鼠选取6 张切片标本,显微镜下观察凋亡情况,以Image J 软件对照片中阳性细胞进行计数。1.7 苏木精-伊红(HE)染色
利用HE 染色检测脑组织病理损伤,切片置于梯度乙醇溶液水化,蒸馏水冲洗1 min;苏木精染液加热染色2 min,流水洗去苏木精液;l%盐酸乙醇分化30 s,流水洗;1% 氨水反蓝30 s,流水洗;伊红染液染色2 min,流水洗;梯度乙醇溶液脱水:80 %乙醇1 min,95% 乙醇1 min,无水乙醇1 min;切片晾干,中性树胶封片,光镜下观察组织学改变。1.8 酶联免疫吸附测定(ELISA)检测IL-1β、IL-6及TNF-α水平
我们于损伤后3 d,利用ELISA检测各组小鼠脑组织中炎性因子IL-1β、IL-6及TNF-α 的水平变化。各组取损伤周围脑组织100 mg,加入预冷的生理盐水,制备成10% 脑组织匀浆液,电动匀浆器匀浆,8 000 r/min,离心20 min,取上清,随后根据试剂盒说明书进行操作。酶标仪450 nm 波长处检测各孔吸光度值,根据标准品吸光度,绘制标准曲线,再根据标准曲线求出各组小鼠脑组织中IL-1β、IL-6及TNF-α的含量。1.9 蛋白质印迹法(Western blotting)检测NF-κB信号通路相关蛋白
各组取损伤周围脑组织100 mg,加入RIPA 裂解液,电动匀浆器匀浆,10 000 r/min,离心30 min,取上清;随后利用BCA 蛋白定量试剂盒进行蛋白定量,加入上样缓冲液,加热10 min变性,之后进行SDS-PAGE 电泳(100 V,90 min)、转膜(250 mA,90 min)、加入相应Iba-1(1∶1 000)、GFAP(1∶2 000)、NF-κB p65(1∶1 000)、IκB-α(1∶1 000)、p-IκB-α(1∶1 000)及β-actin(1∶1 000)一抗稀释液孵育,4 ℃孵育过夜,次日室温下孵育相应二抗(1∶2 000),ELC 显色液显色后,凝胶成像分析系统扫描并拍照,利用Image J软件进行光密度分析。
2 结果
2.1 MB 降低NSS 评分
分别于损伤后3、7、14 及21 d后对小鼠神经功能进行评价,实验结果显示,与Sham 组相比,SWI 组小鼠NSS 评分明显升高,差异有统计学意义(P
<0.05);而给予MB 治疗后能够明显降低NSS评分,差异有统计学意义(P
<0.05),说明MB 能够促进SWI 小鼠神经功能恢复;给予PDTC 之后,PDTC、MB+PDTC 组与SWI 组相比NSS 评分降低不明显,差异无统计学意义(P
>0.05),且PDTC 与MB+PDTC 组相比,差异无统计学意义(P
>0.05),说明PDTC 抑制了MB 改善SWI 小鼠神经功能的作用(表1)。
表1 各组小鼠神经功能缺损评分比较/(分,± s)
2.2 MB 抑制神经细胞凋亡
于损伤后7 d 利用TUNEL 试剂盒检测神经细胞凋亡情况,观察MB 对SWI 后大脑皮层神经细胞凋亡的影响。结果见图1和表2,SWI组凋亡细胞数量显著增加,与Sham组相比,差异有统计学意义(图中蓝色为细胞核DAPI,红色为凋亡细胞,P
<0.01),MB 组与SWI 组相比,能够显著降低凋亡细胞数量(P
<0.05),说明MB 能够减少脑损伤小鼠神经细胞凋亡。PDTC 及MB+PDTC组与SWI 组相比,T 凋亡细胞数量明显减少,且MB组与PDTC 及MB+PDTC 组相比,差异无统计学意义(P
>0.05),提示PDTC可抑制MB的抗凋亡作用。
表2 各组小鼠脑组织神经细胞凋亡数量比较

2.3 MB 减轻脑组织病理损伤
利用HE 染色检测各组小鼠大脑皮层病理损伤情况,结果见图2,Sham组皮层神经元清晰致密、染色形态正常、排列均匀、胞体饱满、胞核呈蓝色;而SWI 组伤口周围可见大量的红细胞,炎性细胞浸润明显,神经元皱缩、胞质深染、核不清晰、坏死明显;而MB、PDTC 及MB+PDTC 组伤口周围神经元形态明显改善、炎症细胞浸润减轻、坏死细胞数量减少,红细胞溢出情况减轻,且三组之间无明显差别。2.4 MB 减轻SWI所致炎症反应
为了探究MB对SWI 引起的炎症反应的影响,我们检测了小鼠脑组织中炎性因子IL-1β、IL-6 及TNF-α 的表达水平;并检测了脑组织中Iba-1、GFAP 蛋白表达情况。ELI‑SA 结果显示,SWI 组小鼠损伤皮层IL-1β、IL-6 及TNF-α 水平明显升高,与Sham 组相比,差异有统计学意义(P
<0.01);MB 组、PDTC 组小鼠脑组织中IL-1β、IL-6 及TNF-α 水平明显降低,与SWI 组相比,差异有统计学意义(P
<0.05,P
<0.01),而MB 组与PDTC 组、MB+PDTC 组相比,差异无统计学意义(P>
0.05)。见表3。
表3 各组小鼠脑组织中白细胞介素(IL)、肿瘤坏死因子α(TNF-α)水平比较/(ng/mL,± s)
Western blotting 结果显示,小鼠脑损伤后Iba-1、GFAP 蛋白表达明显增加,与Sham 组相比,差异有统计学意义(P
<0.01),MB 能够明显减少脑内Iba-1、GFAP蛋白表达,差异有统计学意义(P
<0.01);PDTC及MB+PDTC 组与SWI 组相比,Iba-1、GFAP 蛋白表达明显减少,且MB、PDTC 及MB+PDTC 组之间相比,差异无统计学意义(P
>0.05),提示PDTC 可减弱MB对炎症反应的抑制作用。见图3与表4。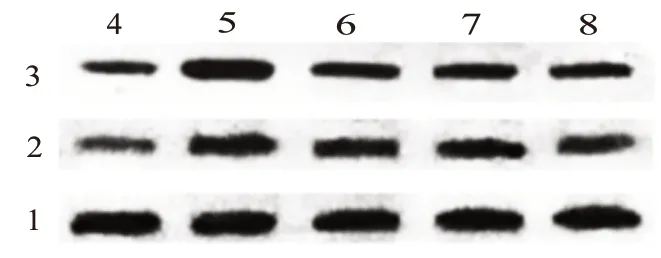
图3 各组小鼠脑组织中Iba-1及GFAP蛋白表达(Western blotting检测图)
表4 各组小鼠脑组织中小胶质细胞(Iba-1)、星形胶质细胞(GFAP)蛋白水平比较/± s
2.5 亚甲蓝抑制NF-κB 信号通路相关蛋白表达
为了研究MB 治疗对SWI 小鼠脑组织NF-κB 信号通路活性的影响,我们检测了NF-κB 信号通路相关蛋白的表达。结果显示,与Sham 组相比,SWI 组NFκB p-65、p-IκB-α 水平显著升高,差异有统计学意(P
<0.05),而IκB-α 蛋白水平明显降低,差异有统计学意义(P
<0.05);MB 组小鼠脑组织中NF-κB p-65、p-IκB-α 蛋白表达明显降低,而IκB-α 蛋白水平明显升高,与SWI 组相比,差异有统计学意义(P
<0.05);而MB 组与PDTC 组、MB+PDTC 组相比,NF-κB 信号通路相关蛋白表达变化不明显,差异无统计学意义(P>
0.05,图4 与表5),提示给予PDTC 后MB 抑制NFκB 信号通路活性的作用被阻断,说明MB 对SWI 小鼠的治疗作用可能与抑制了NF- κB 信号通路有关。
表5 各组小鼠脑组织中NF-κB p-65、IκB-α及p-IκB-α蛋白水平比较/± s

图4 各组小鼠脑组织中NF-κB p-65、IκB-α及p-IκB-α蛋白表达(Western blotting检测图)
3 讨论
继发性脑损伤在损伤后数小时内发生,继发损伤的基本机制主要包括氧自由基、神经炎症、脑水肿和神经元凋亡。脑损伤诱发的神经炎症主要以小胶质细胞和星形胶质细胞活化为主要特征,损伤发生后脑内的小胶质细胞和星形胶质细胞活化并迁移至损伤周围,并且能分泌大量的炎性因子,比如,IL-6,IL-1β 和TNF-α,破坏血脑屏障,从而加剧了脑水肿、炎症级联反应及神经元凋亡。因此,如何抑制脑损伤后炎症反应,减少神经细胞凋亡,恢复患者受损神经功能一直是本研究领域的关键。
MB 是在临床上广泛使用并且在治疗剂量范围内无明显不良反应的药物。MB 从被发现后,曾先后用于治疗高铁血红蛋白血症、疟疾、氰化物中毒、脓毒血症及外科手术的示踪剂等。有研究表明,MB 的LD腹腔注射给药为150 mg/kg,而静脉给药LD为77 mg/kg。而且一般认为,静脉给药剂量在10 mg/kg 以下都是安全的,并且其起治疗作用的剂量通常也是低剂量,一般在4 mg/kg 以内。近年来研究表明MB 在治疗中枢神经系统疾病方面具有明显的药理作用。MB 能够显著降低APP/PS1 转基因小鼠脑实质和脑血管β-淀粉样蛋白沉积物以及各种Aβ 物种(包括寡聚物)的水平,从而改善其认知功能障碍。MB 能够通过上调脑源性神经营养因子来保护多巴胺能神经元,减轻MPTP 对神经元诱导的神经毒性。在脑损伤小鼠模型中,在损伤1 d 后,MB 显著改善脑组织水肿并降低了炎性基因在巨噬细胞及中性粒细胞中表达。在我们的实验中,我们也发现,应用MB 腹腔注射1 mg/kg 的剂量能够明显改善小鼠神经功能缺损评分,阻止SWI导致的神经细胞凋亡并能改善病理损伤。
脑损伤后会引发继发性炎症反应,表现为星形胶质细胞及小胶质细胞的异常活化及IL-1β、IL-6、TNF-α 等炎性因子的过度释放。IL-1β 和TNF-α 是SWI 中最早被释放的炎性因子,两者还能够发挥协同作用,共同促进其他炎性因子的生成与释放;IL-6 也是炎症反应过程中一个重要的炎性因子。本实验发现MB 能显著抑制SWI 小鼠模型脑组织中星形胶质细胞及小胶质细胞的活化,并降低损伤脑组织周围IL-1β、IL-6和TNF-α 因子的表达,提示MB能够通过抑制炎性因子的生成与释放及胶质细胞活化,减轻SWI引起的炎症反应。
NF-κB信号通路在调节细胞的生长、凋亡、分化及炎症等方面具有重要作用,NF-κB 通路的激活可诱导IL-6,IL-1β和TNF-α等炎性因子的表达,且IL-6,IL-1β 和TNF-α 等炎性因子释放增加也会激活NF-κB信号通路,形成反馈环路,从而诱发机体的炎症反应。抑制NF-κB 信号通路使IκB-α 的磷酸化被抑制,并能阻止因NF-κB 从IκB-α 中释放并转移到细胞核后所引发的炎症级联反应。并且NFκB 信号通路的活化能够参与皮层及海马区的神经细胞凋亡,脑损伤后NF-κB 在神经细胞中广泛表达,诱导神经细胞产生促凋亡基因及蛋白,经过级联反应,最终导致神经元的凋亡。的本研究发现,MB 能够显著降低脑损伤小鼠NF-κB p-65、p-IκB-α蛋白表达,而升高IκB-α蛋白水平,说明MB对脑损伤小鼠NF-κB 信号通路相关蛋白具有调控作用。
为了探讨MB 对脑损伤小鼠模型的治疗作用与调控NF-κB信号通路有关,在制备SWI小鼠模型前,利用100 mg/kg 的PDTC 封闭NF-κB 信号通路,抑制其活性。结果显示,NF-κB 信号通路被封闭后,MB 对小鼠神经功能恢复、神经细胞凋亡的影响、对炎症反应的影响及对NF-κB 信号通路相关蛋白的调控作用被抑制,进一步说明MB 减轻继发性脑损伤的作用机制可能是通过抑制NF-κB 信号通路活性实现的。
综上所述,本研究表明,MB 能改善脑损伤后小鼠的神经功能、抑制细胞凋亡、减轻损伤后的炎症反应,这可能在一定程度上与抑制了NF-κB 信号通路活性有关,但具体的分子机制还需要进一步研究。虽然目前MB 距离成为脑损伤的临床治疗药物还有很远的路要走,但相信在今后有一天通过剂型优化、结构修饰等手段能够降低甚至消除亚甲蓝的毒副作用,其能够成为安全有效的脑损伤临床治疗药物。
