不同克氏原螯虾养殖模式下幼虾生长、体成分和血淋巴生化指标的比较
2021-10-08魏开金马宝珊朱祥云
徐 滨,魏开金,马宝珊,徐 进,朱祥云,刘 伟
(中国水产科学研究院长江水产研究所,农业农村部淡水生物多样性保护重点实验室,武汉 430223)
养殖模式指在某一特定条件下,使养殖生产达到一定产量而采用的经济与技术相结合的规范化养殖方式。由于养殖模式的不同,在生长特性和品质特性上会有所差异[1]。养殖模式会影响养殖对象的生存对策,同时也会改变其生命代谢特征[2-3]。生长性能、体成分、血清生化是衡量水产养殖动物健康程度的重要指标,也是其生存状况、生命代谢的重要依据[4-5],对比不同养殖模式生长性能、体成分、血清生化指标,揭示由养殖模式引起的生存对策、生理代谢的变化本质,对推进水产动物的健康高效养殖具有重要意义。
克氏原螯虾(Procambarusclarkii)俗称小龙虾,属甲壳纲十足目螯虾科原螯虾属,原产于墨西哥北部和美国南部,是我国重要的经济水产养殖对象之一,也是目前淡水虾类增养殖的主要品种[6-7]。近年来,国内外学者对克氏原螯虾的关注与研究较多[8-12],同时随着克氏原螯虾价格的提高、养殖规模的扩大,出现了多种多样的养殖模式,养殖模式的发展对克氏原螯虾养殖起着积极的促进作用,而不同养殖模式的养殖效果研究还存在空白。本研究通过对江汉平原池塘单养、虾鱼混养、虾蟹混养、稻虾养殖四种养殖模式的幼虾的生长、体成分、血淋巴生化指标等进行了分析,为比较不同养殖模式的特点,生产优质绿色水产品提供基础资料。
1 材料与方法
1.1 养殖实验
实验用虾均取自湖北潜江水产局后湖稻虾养殖基地,其中虾单养、虾鱼混养、虾蟹混养养殖实验于湖北百瑞生物公司仙桃基地内进行,实验池为水泥池,大小为(6 m×6 m×1 m),小龙虾养殖密度12 尾/m2,虾鱼(小龙虾:泥鳅)和虾蟹(小龙虾:中华绒螯蟹)数量比均为12 ∶1。每组设3个平行,养殖用水为经曝气过滤的室外池塘水,投喂饲料为市场上常见的虾蟹饲料,蛋白含量为30%,日投喂量为虾体重的5%~7%,同时每隔10 d,投放水草一次;稻虾养殖在湖北潜江水产局后湖稻虾养殖基地进行(每口稻田面积33 330 m2,共20口,在稻田四周挖沟,沟宽3~4 m,沟深1~1.5 m,中晚稻,6月份插秧),养殖密度12 尾/m2,同时种植水草,投喂饲料同为蛋白含量为30%的虾蟹饲料,日投喂量为虾体重的5%~7%。定期用YSI EXO2水质分析仪监测水温、溶氧、pH、叶绿素、蓝绿藻、氨氮、硝氮等水质指标,养殖周期为4月5日至6月5日,各实验组水温17.8~27.7 ℃,溶氧5.5~8.9,pH7.3~7.9。
1.2 样品采集及测定
养殖实验共进行60 d,每隔10 d每组取样50尾克氏原螯虾,用吸水纸吸干虾体表水分后,分别用游标卡尺和电子天平测量虾的体长(精确到0.01 mm)和体重(精确到0.01 g)。养殖实验结束后,每组取10尾测定全虾的体成分,另取8尾测定虾的血淋巴生化指标。先测量虾体长、体重,然后用1 mL一次性注射器从头胸甲缝隙插入围心腔内取血淋巴,放入2 mL离心管内,静止2 h后,4 ℃、3 000 r/min离心5 min吸取上层血清,-80 ℃冻存备用。
全虾的粗脂肪含量测定采用索氏抽提法(乙醚);粗蛋白含量测定采用凯氏定氮法(GB/T5009.5-2003);灰分含量测定采用550 ℃马弗炉灰化法(GB/T5009.4-2003);水分采用105 ℃恒温烘干恒重法测定(GB 6435-86);血清生化指标(谷丙转氨酶、谷草转氨酶、葡萄糖、胆固醇、甘油三脂、总蛋白、白蛋白)采用CHEMIX-800全自动生化分析仪测定。
1.3 数据统计与分析
数值采用均用平均值±标准差表示,所得数据均以Excel与SPSS 21.0软件作单因素方差分析、Pesrson相关系数分析和Duncan氏多重比较分析,显著性水平为P=0.05。
体重增长率(WGR)=(W1-W0)/W0×100%
体长增长率(LGR)=(L1-L0)/L0×100%
式中,W0和W1分别为实验虾测量时间段内的起始、结束体重(g),L0和L1分别为实验虾测量时间段内的起始、结束体长(cm)。
2 结果与分析
2.1 四种养殖模式克氏原螯虾生长性能比较
由图1可见,在实验开始至10 d时,4个模式的幼虾体长、体重基本处于同一水平,无显著差异。随着试验的进行,体长和体重均呈现虾单养组>虾鱼组>虾蟹组>稻虾组,且体重比体长差异更大。在20 d,稻虾组体长显著低于其余三组,虾单养组体重显著大于其余三组,其余各组差异不显著;在30 d,体长和体重均表现为虾单养组显著大于另外3组,稻虾组显著低于另外3组,虾鱼组和虾蟹组无显著差异;40 d,稻虾组体长显著低于另外3组,虾单养组体重显著大于另外3组,稻虾组体重显著低于另外3组,虾鱼组和虾蟹组体重无显著差异;至50 d,虾单养组体长显著高于稻虾组,虾单养组和虾鱼组体重显著高于稻虾组,其余各组差异不显著;至60 d体长和体重均表现为虾单养组显著大于另外3组,稻虾组显著低于另外3组,虾鱼组和虾蟹组无显著差异。
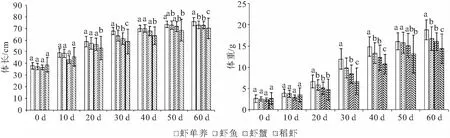
图1 四种养殖模式克氏原螯虾的体长和体重变化
四种养殖模式下,克氏原螯虾LGR随时间大体呈逐渐下降的趋势,WGR率则呈先升高后降低的趋势,见图2。其中LGR在前30 d体长保持较高增长,然后逐渐降低。WGR在前40 d保持较高增长,然后逐渐降低,虾单养组、虾鱼组和虾蟹组WGR在30 d达到最高,稻虾组在40 d达到最高。
2.2 四种养殖模式水环境理化指标变化
在60 d的养殖期间,水温在前30 d一直保持升高,而后随天气变化而变化。溶氧则是先升高后降低,在前30 d增加较快,而后迅速降低。pH则是先降低后升高,开始时虾单养组最高,稻虾组最低,试验结束时稻虾组最高,4个模式组均在40 d时pH最低。叶绿素和蓝绿藻含量在实验开始时4个组均处于较低水平,而后均开始增长,20 d至60 d稻虾组叶绿素和蓝绿藻含量最低且差异显著。硝氮和氨氮随着养殖的持续不断升高。其中稻虾组的氨氮在实验开始时显著大于其余三组,至60 d,稻虾组氨氮含量显著低于另外三组。硝氮无显著差异。
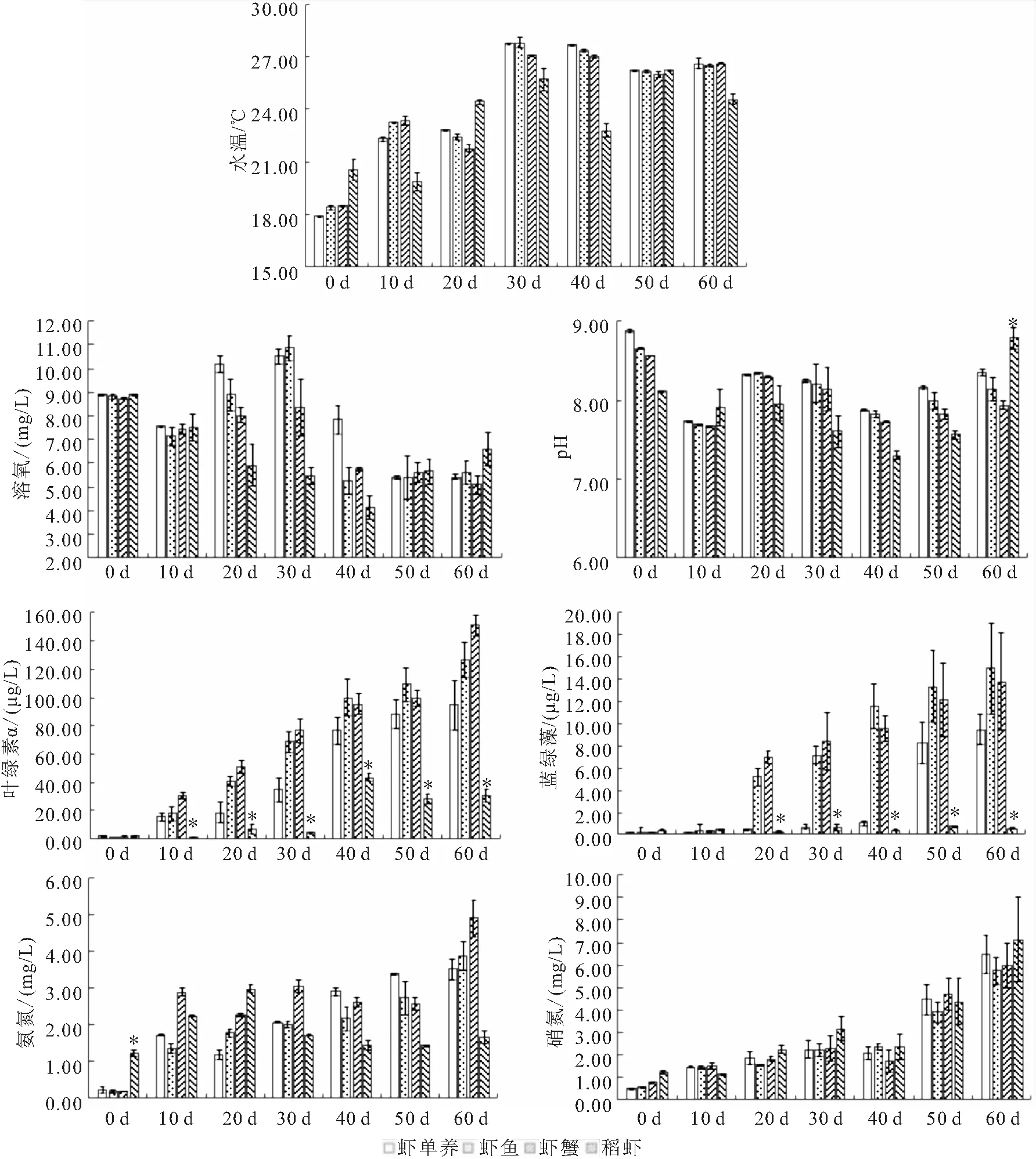
图3 养殖期间水体理化指标变化
2.3 四种养殖模式克氏原螯虾体成分比较
四种养殖模式克氏原螯虾的体成分组成见表1,结果显示,全虾的水分含量稻虾组>虾鱼组>虾蟹组>虾单养组,稻虾组最高,显著高于虾单养组、虾蟹组,与虾鱼组差异不显著。全虾的粗蛋白含量虾蟹组>虾单养组>虾鱼组>稻虾组,虾蟹组最高,显著高于另外3组,其余三组间差异不显著。全虾粗脂肪含量稻虾组>虾鱼组>虾蟹组>虾单养组,虾单养组最低,显著低于另外3组,其余三组间差异不显著。全虾的灰分虾单养组>虾蟹组>虾鱼组>稻虾组,虾稻组最低,显著低于另外3组,虾单养组与虾鱼组也差异显著,其余各组间差异不显著。
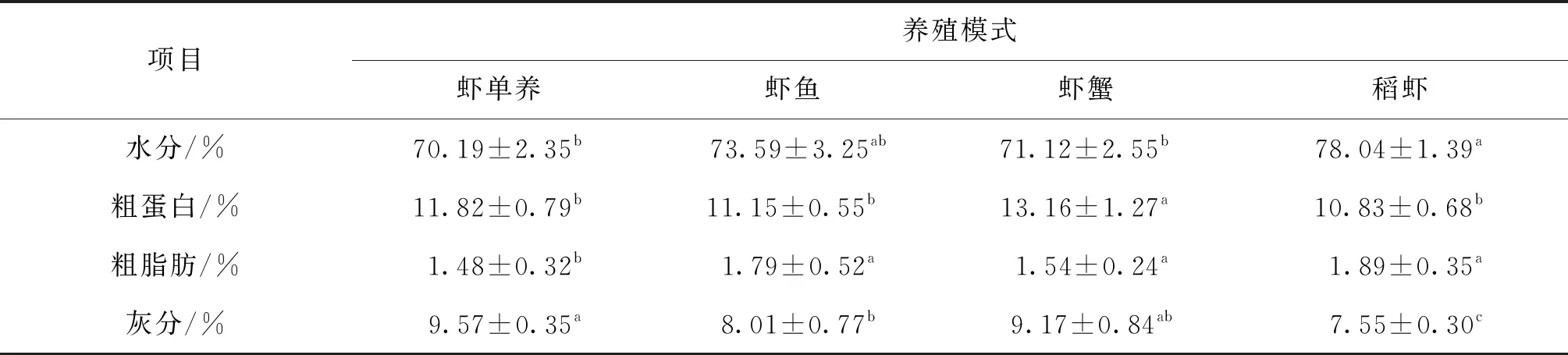
表1 四种养殖模式克氏原螯虾的全虾常规组成
2.4 四种养殖模式克氏原螯虾血淋巴生化指标比较
养殖实验结束后,四种养殖模式克氏原螯虾的血清生化指标见表2。总蛋白为虾蟹组>虾鱼组>虾单养组>稻虾组,其中虾蟹组、虾鱼组与稻虾组差异显著,其余各组差异不显著。白蛋白与总蛋白类似,虾鱼组>虾蟹组>虾单养组>稻虾组,虾鱼组、虾蟹组与虾单养组、稻虾组差异显著,其余各组差异不显著。总胆固醇含量,虾单养组>虾鱼组、虾蟹组>稻虾组,虾单养组显著高于其余3组,其余各组差异不显著。甘油三脂含量稻虾组>虾鱼组>虾单养组>虾蟹组,稻虾组显著高于其余虾单养组和虾蟹组,其余各组差异不显著。葡萄糖含量虾鱼组>虾蟹组>稻虾组>虾单养组,虾鱼组与虾单养组差异显著,其余各组差异不显著。谷丙转氨酶含量,虾单养组>稻虾组>虾鱼组>虾蟹组,虾鱼组和虾蟹组差异显著,其余各组差异不显著。谷草转氨酶含量稻虾组>虾单养组>虾鱼组>虾蟹组,其中稻虾组与虾蟹组差异显著,其余各组差异不显著。

表2 四种养殖模式克氏原螯虾的血液生化指标
3 讨论
3.1 四种养殖模式克氏原螯虾生长性能比较
克氏原螯虾的生长受到其生物习性、食物资源、发育阶段、养殖密度及环境条件等多因素的影响。克氏原螯虾适宜的水温范围为21~30 ℃,当水温高于32.2 ℃时,虾的生长则会受到影响,当水温降至5~6 ℃时,幼虾会停止摄食[13]。同时,水体溶氧也会影响其生长,溶氧降低会引起摄食的减少[14-15],幼虾的适宜pH范围为7.5~8.5[16]。本实验条件下水温、溶氧和pH与建议值均比较接近。本实验期间,水温在10 d略低于20 ℃,其体重增长率略低,但其体长增长率是整个试验期间最快,我们推测跟幼虾所处的发育阶段相关。在实验前10 d,四个模式的幼虾体长、体重无显著差异。随着试验的进行,体长和体重均呈现虾单养组>虾鱼组>虾蟹组>稻虾组,且体重比体长差异更大,克氏原螯虾的体长增长率大体呈逐渐下降的趋势,体重增长率则呈先升高后降低的趋势。克氏原螯虾为蜕壳生长,蜕壳后体长才能增加,结果表明克氏原螯虾在试验前30 d蜕壳较多,体长生长较快。
克氏原螯虾生长指标从高到低依次为虾单养组、虾鱼组、虾蟹组、稻虾组,分析其原因,一是前三组试验池较小,水温相比稻田升高快,水体的叶绿素和蓝绿藻水平较高,能给幼虾提供额外的饵料,有利于幼虾的生长;其次,虾鱼和虾蟹模式的养殖对象之间存在竞争关系,导致这两组生长速度略低于虾单养组。虾单养组、虾鱼组及虾蟹组的体重增长率均在30 d时达到最高,而稻虾组的体重增长率在40 d时达到最高,且稻虾组的标准差比其余三组大,表明稻虾组的幼虾规格不整齐,推测稻虾模式应该有去年打洞的留塘抱卵亲虾,该批虾苗处于不同的生长阶段的原因。
3.2 四种养殖模式克氏原螯虾体成分比较
克氏原螯虾的全虾体成分主要包括粗蛋白、粗脂肪、灰分、水分等,体成分的含量将影响其营养价值与经济价值。体成分是其对特定生境的表型适应,受环境中饵料的丰富程度所驱动,其中蛋白质和脂肪决定了机体的营养水平,已有研究表明蛋白质主要用于动物的生长发育,脂肪主要用于为机体储存能量[17]。对水产动物进行营养成分分析有助于了解其经济应用价值,并为人工饲料的配置提供理论依据。四种养殖模式克氏原螯虾的体成分组成差异较大,全虾的水分含量稻虾组显著高于虾单养组、虾蟹组。全虾的粗蛋白含量虾蟹组显著高于另外3组。全虾粗脂肪含量虾单养组显著低于另外3组。全虾的灰分稻虾组显著低于另外3组。一般养殖品种的个体水分含量低于野生个体,实验结果显示稻虾组的水分含量显著高于其余3组,表明稻虾组更接近野生个体。同时稻虾组虾可以在稻田内打洞穴居,由于竞争少,能量消耗少,导致多余的能量以脂肪形式储存在体内,所以粗脂肪高于其余三组。虾蟹组因其存在较大竞争关系,需要更多的能量用于生存,蛋白含量显著高于另外三组。
3.3 四种养殖模式克氏原螯虾血淋巴生化指标比较
血清生化指标被广泛用于评价动物的营养和健康状况,反映动物体内器官功能状态和物质代谢的重要特征[18]。影响水产动物的血液生化指标的因素很多,包括:养殖模式、温度、盐度、光照、重金属、饥饿、饲料等。
总蛋白可以维持血液渗透压和pH平衡,具有运输、凝血、免疫等功能。白蛋白功能包括营养、保持血液渗透压以及对离子、胆红素、脂肪酸、激素等运输[19]。总蛋白为虾蟹组>虾鱼组>虾单养组>稻虾组,其中虾蟹组、虾鱼组与稻虾组差异显著,白蛋白为虾鱼组>虾蟹组>虾单养组>稻虾组,虾鱼组、虾蟹组与虾单养组和稻虾组差异显著。一般认为,血清中总蛋白含量增加是体内营养成分充足,代谢较快,蛋白质合成旺盛,分泌到血液中的蛋白质增多造成的[4],鱼类在低溶氧条件下总蛋白和白蛋白含量会降低[20],这与稻虾组溶氧较低吻合。结果表明虾蟹组和虾鱼组虾的体内营养充足、新陈代谢较快,患病少。
脂类是甲壳动物重要的能源储备物质[21-22],甘油三脂和胆固醇是血脂的主要成分,主要在肝脏合成,并经血液循环至全身各处[23]。过多摄食脂类食物和肥胖可使血清甘油三酯升高,而运动则可以降低甘油三酯[4,17]。甘油三脂含量稻虾组>虾鱼组>虾单养组>虾蟹组,稻虾组显著高于虾单养组和虾蟹组,表明稻虾组运动低于其余三组,这与体成分结果相印证。饱和脂肪酸的摄入会促进机体胆固醇的合成,进而使血清胆固醇升高,总胆固醇含量,虾单养组>虾鱼组、虾蟹组>稻虾组,表明稻虾组虾的饱和脂肪酸的摄入低于另外三组。
葡萄糖是提供机体活动所需的能量,受栖息环境等许多因素影响,流水环境较静水高,活动较少的生物血糖浓度较低[23]。葡萄糖含量为虾鱼组>虾蟹组>稻虾组>虾单养组,表明虾鱼组和虾蟹组虾的水里活动多于稻虾组和虾单养组,混养种类存在竞争关系。Imsland等[24]认为在正常情况下,鱼类体内有机成分的分解代谢与合成代谢保持动态平衡,血糖含量较高时,鱼类表现为积极摄食,健康状况良好。
转氨酶可以反映肝脏健康状态,在正常状态下转氨酶只有少量释放到血液中,血清转氨酶活性较小;当肝脏发生病变时,细胞的通透性增加,导致转氨酶大量释放到血液内,使谷草转氨酶和谷丙转氨酶升高[25]。一般认为血液中转氨酶活性增加表明肝脏功能出现障碍[5,26]。谷丙转氨酶含量,虾蟹组<虾鱼组<稻虾组<虾单养组,虾单养组和虾蟹组差异显著。谷草转氨酶含量虾蟹组<虾鱼组<虾单养组<稻虾组,其中稻虾组与虾蟹组差异显著。本结果表明虾蟹组虾的转氨酶含量最低,肝脏最健康,这可能与虾蟹组叶绿素和蓝绿藻水平较高,幼虾摄入浮游生物较多有关。本实验是在4-5月份进行,池塘的水草及浮游生物随水温升高快速生长,而稻虾是在5月底开始插秧,稻虾组的叶绿素和蓝绿藻水平偏低。综上所述,从生长角度看虾单养组生长最快,从虾体成分看稻虾组最接近野生群体,从血清生化指标看虾蟹组最健康,我们建议在积极开展稻虾综合种养的同时,在养殖产量较少的地区可以适当增大池塘单养的规模,在养殖规模较大的地区适当开展虾鱼、虾蟹模式。
