长江上游宜宾-江津与涪陵-丰都江段鱼类早期资源分布研究
2021-10-08张先炳杨胜发李文杰童思陈
张先炳,杨胜发,杨 威,李文杰,肖 毅,胡 江,王 丽,童思陈
(国家内河航道整治工程技术研究中心,重庆市生态航道重点实验室,重庆交通大学,重庆 400074)
长江上游珍稀特有鱼类国家级自然保护区分布的鱼类高达286种,其中上游特有鱼类124种[1],包括濒危的长江鲟(Acipenserdabryanus)和胭脂鱼(Myxocyprinusasiaticus)等,具有不可替代的物种价值[2]。随着长江上游向家坝电站和三峡水库的运行、航道开发、采砂、沿线城镇取水等活动的开展,长江上游的河流形态和水文过程均有一定程度的改变[3],进而影响鱼类栖息、生长、繁殖所需的生境条件[4-5],最终导致鱼类资源的变动。
鱼类早期资源的变化能很好地反映鱼类的繁殖规模和种群的变动趋势[6],其可作为评估长江上游水利工程和航道建设对鱼类资源影响程度的基础资料。已有研究报道了长江上游某些河段如金沙江下游[7-8]、岷江下游[9]、泸州[10]、丰都[11]等江段鱼类早期资源的监测结果。但对于长江上游卵苗过境峰值与水文过程相互关系以及鱼类早期资源的空间分布差异性还不够完善。因此,为探寻鱼类产卵与水文动态过程的相互关系,本实验于2019年6-7月对长江上游宜宾-江津江段和涪陵-丰都江段的鱼类早期资源的动态过程进行了连续监测,分析了卵苗空间分布的差异性、过境峰值与涨水的关系和鱼卵败育的原因。
1 研究方法
1.1 卵苗采样点设置及采集方法
本次采样设置的五个点(见图1),重庆上游为南溪、朱沱和江津,重庆下游为涪陵和丰都,参照三峡库区干流采样点的设置方法[12],于2019年6月22日-7月13日进行采样监测,图1中的公里数表示采样点之间的距离。各采样点的经纬度坐标分别为南溪(北纬28°50′40.275 6″,东经104°59′13.58″)、朱沱(北纬29°2′0.48″,东经105°52′55.41″)、江津(北纬29°13′54.59″、东径106°13′34.99″)、涪陵(北纬29°42′50.99″,东经107°17′25.37″)、丰都(北纬29°51′7.11″、东经107°39′29.99″)。
参照范思林等[13]的方法采用自制的卵苗采集装置(见图1),网口尺寸为1 m×1 m,附带有2 cm×2 cm的格栅网,用于截留江水中的漂浮物,网口的四角采用尼龙绳固定在趸船上,卵苗采集时,尽量保持进水方向与网口平面垂直;网口与鱼卵采集器之间的琼网为圆锥形,宽口端连接进水网口,窄口端连接卵苗收集器,长度为2.5 m,圆形窄口段直径为10 cm;卵苗收集器为边长40 cm正方体框架结构,装有顶部不封口的可拆卸网兜(孔眼直径1 mm)以收集卵苗,卵苗收集器顺水的两侧装有两个直径25 cm的浮筒,使得鱼卵采集器工作时漂浮于水面。

图1 漂流性鱼卵采样点位置及采样装置照片
连续20 d于每日10:00及15:00进行样品采集,每次采样时间为15 min,并采用流速仪(裕泽,ls1206b)同步测定流速,便携式溶氧仪(哈希,HQ30D)测定水温,同步记录临近水文站的流量和水位。
1.2 卵苗的形态观测及种类鉴定
采集的鱼卵和仔鱼用OPTIKA B-10000荧光显微镜观测形态特征并拍照,根据形态特征甄别鱼卵的发育阶段,并比较其卵核是否具有完整结构或者有无卵核离散现象来判断鱼卵是否败育,之后结合形态学的特征对鱼卵和仔鱼进行种类的初步鉴定[14]。同时将未鉴定种类的样品用无水乙醇保存,返回实验室提取DNA,而后对细胞色素b基因[15]进行PCR扩增后测序(上海生工生物工程股份有限公司)。测序结果用Chromas软件比对峰型图并剪辑后,用DNAMAN软件对双向测序结果进行拼接、比对和归类,然后将基因序列在NCBI网站(http://blast.ncbi.nlm.nih.gov/Blast.cgi)中进行比对,依据匹配程度来确定鱼卵和仔鱼种类。
1.3 鱼类早期资源量估算
调查期间,漂流经过采样断面的鱼卵、仔鱼总量估算参照易伯鲁[16]等的计算方法。
2 结果
2.1 卵苗的数量分布与涨水过程
采集的鱼卵和仔鱼的数量见图2,共采集到2 034粒鱼卵和1 086尾仔鱼。其中南溪和朱沱采集的鱼卵数量较少,但采集的仔鱼数量较多。宜宾-丰都江段鱼类早期资源的径流总量沿程增加,采样期间从上至下各采样点的累积过境卵苗总量估算值分别为:0.56亿、0.83亿、2.90亿、5.15亿、7.67亿ind.。

图2 各采样点鱼卵及仔鱼采集数量(a)及资源量估算(b)
鱼卵数量与流量和水位的动态变化过程见图3和表1,在6月24日、6月29日、7月3日和7月10日前后均出现卵苗数量的激增。以江津(6月29日)和涪陵(6月30日)为例,鱼卵数量峰值对应的流量涨幅为20%和54%,水位分别增长了0.74 m和2.76 m。卵苗数量峰值出现在持续涨水2 d以后,重庆上游的南溪、朱沱和江津的流量日涨幅超过10%时且水位日涨幅在0.5 m以上;重庆下游的涪陵和丰都属于三峡水库的变动汇水区,卵苗出现峰值时的流量日涨幅超过40%且水位日涨幅大于2 m。

表1 鱼卵数量与生态水文指标的关系
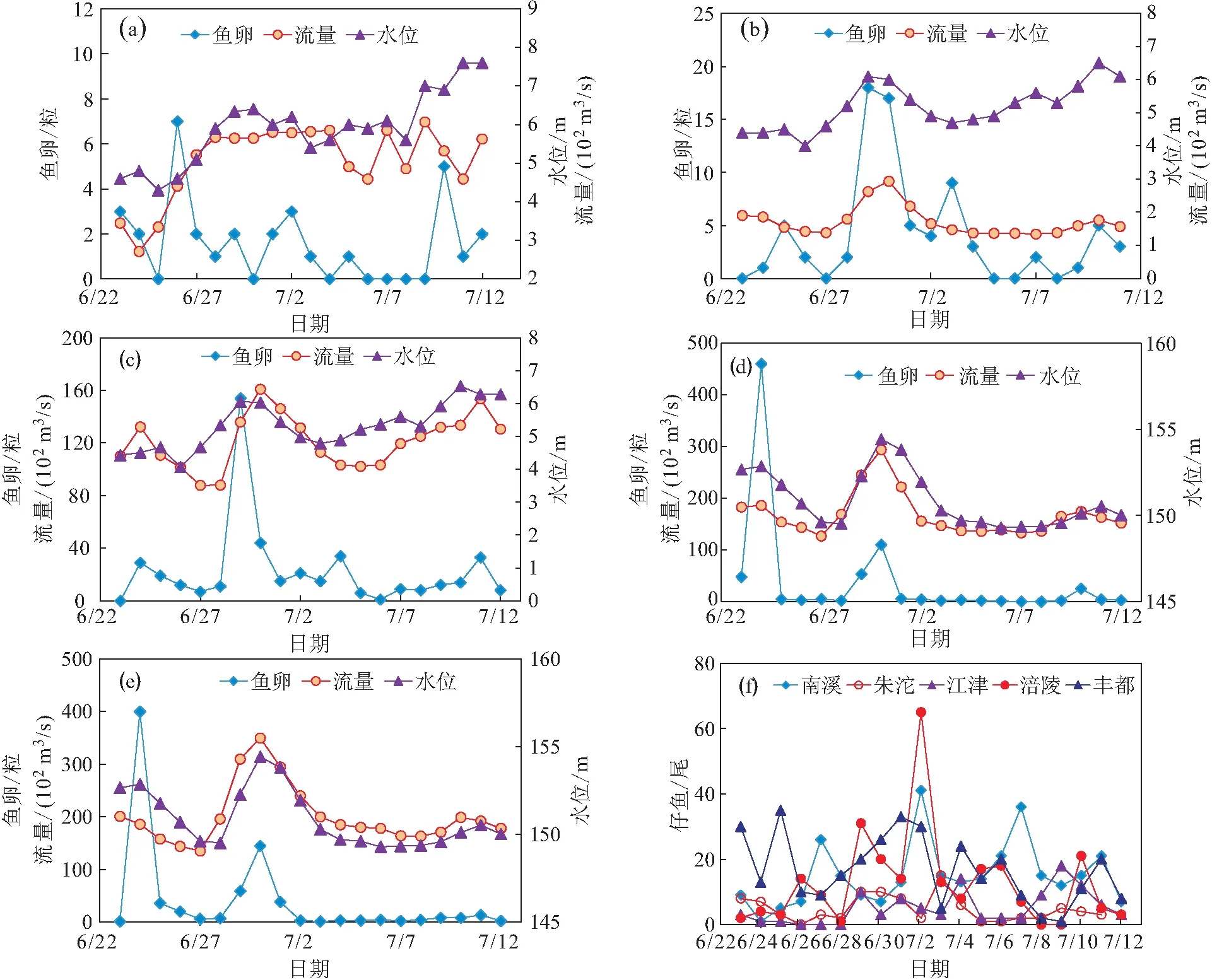
图3 各采样点鱼卵数量、水位和流量的变化趋势及仔鱼的数量波动
2.2 漂流性鱼卵的发育程度
5个采样点正常发育鱼卵509粒所处发育阶段及其占比见图4。其中,各发育时期鱼卵数量分别为:细胞期10粒、囊胚晚期24粒、原肠晚期29粒、胚胎封闭期60粒、眼囊期64粒、尾芽期53粒、晶体出现期92粒、内效应期84粒、孵化期94粒。图4显示上游的南溪和朱沱采集的鱼卵处于发育早期的比例较高,如处于原肠晚期之前发育阶段的鱼卵均发现于重庆上游的采样点,而下游的涪陵和丰都采集的鱼卵大都处于发育中后期。

图4 各采样点处于不同发育阶段鱼卵的数量占比
2.3 卵苗种类的分布
通过形态学和分子生物学的手段对鱼卵和仔鱼进行了种类鉴定,其结果见表2和图5。采集到的漂流性鱼卵共鉴定出1目4科16属18种,其中长江上游特有鱼类9种。5个采样点均采集到草鱼、鲢、鳙、和铜鱼的鱼卵;仅在江津以上河段采集到的鱼卵有10种,分别是青鱼(Mylopharyngodonpiceus)、似鳊(Pseudobramasimony)、寡鳞飘鱼(Pseudolaubucaengraulis)、翘嘴鲌(Culteralburnus)、中华倒刺鲃(Spinibarbussinensis)、犁头鳅(Lepturichthysfimbriata)、中华金沙鳅(Jinshaiasinensis)、中华沙鳅(Botiasuperciliaris)、花斑副沙鳅(Parabotiafasciata)和双斑副沙鳅(P.bimaculata);而蛇(Saurogobiodabryi)、宜昌鳅鮀(Gobiobotiafilifer)和黄颡鱼的鱼卵仅在涪陵和丰都采集到。仔鱼共鉴定到4目6科38属53种,有14种仔鱼在5个采样点均有分布,其中11种属于鲤科;仅在江津以上分布的仔鱼有34种,如属于近红鲌属、鰁属、金沙鳅属和沙鳅属的鱼类;宜昌鳅鮀和壮体沙鳅(B.robusta)的仔鱼仅在涪陵采样点发现。

表2 2019年6月22日-7月13日南溪至丰都断面鱼类早期资源种类组成

图5 各采样点鉴定的鱼卵及仔鱼种类数量
2.4 鱼卵死亡率分布
在对鱼卵样品进行显微结构观测和分析的同时,发现了大量鱼卵败育(图6)。采集的鱼卵中正常发育有509粒,约占总数的25%;败育的鱼卵为1518粒,约占总数的75%。各采样点的败育率依次为:南溪38%、朱沱68%、江津71%、涪陵80%、丰都72%。

续表2

图6 正常发育鱼卵(a)和败育鱼卵(b)微观照片
3 讨论
3.1 卵苗数量与涨水的关系
对比图3中不同采样点的鱼卵数量、流量和水位的数据可知,从南溪和丰都采样点鱼卵峰值过境时间相差约1 d,此趋势同涨水的水文过程一致[9,10,12]。水流刺激(流量、流速和水位涨幅的刺激)是“四大家鱼”等漂流性鱼卵繁殖的必要条件,在产卵期内涨水持续2 d以上,南溪-宜宾江段流量涨幅超过10%且水位日涨幅>0.5 m时即可能出现规模产卵,而涪陵和丰都江段则需要更大的流量(>40%)和水位涨幅(>2 m)刺激才可能出现大规模的产卵(见表1)。此外,长江上游的产卵现象会持续至7月中旬[17],而对于鱼卵的孵化过程而言则会持续到更晚的时间。根据上述产卵量行为同流量和水位涨幅的关系,通过产卵期流量和水位的监测,可预判漂流性鱼卵的产卵时间、产卵量、鱼卵过境时间等,进而合理统筹水运、发电、防洪、城市取水等多个目标,并最大程度保护鱼类资源。
3.2 长江上游卵苗的空间分布特性
卵苗的数量分布显示(图2),南溪、朱沱和江津断面监测到的仔鱼种类要明显高于丰都和涪陵,而在朱沱和江津采样点鉴别出的漂流性鱼卵类种类要多于其它采样点,像中华金沙鳅等某些特定鱼类的鱼卵只在江津以上江段监测到。南溪采样点虽然采集的鱼卵种类较少,但其上游漫长的江段为鱼类的多样性提供了丰富的生境[18-20],本次采集到了除产漂流性卵鱼类的仔鱼外,还采集到了一些产黏性或沉性卵鱼类的仔鱼(表1)。
鱼卵和仔鱼的分布受鱼类生活习性、产卵特性、水温以及河床底质等生境条件影响。如鲤鱼等在流水和静水环境中均能产卵,对环境的适应性强,其鱼卵在宜宾-丰都均有广泛分布;而像中华沙鳅则偏好于在卵石河底的急流浅滩中产卵[21],此类生境在江津以上江段有更为广泛的分布,而在涪陵-丰都的变动回水区范围类则较少有其适宜产卵的生境[22];而像蛇等中下层小型鱼类,喜生活于缓水沙底处,产卵则多发生于微流水的卵石和沙质底质的浅水河滩,因而在涪陵和丰都变动回水区有发现其产卵[23];像黄颡鱼则喜静水或缓流浅滩,产卵期为5月中旬至7月中旬且繁殖水温要求25~30.5 ℃[24],涪陵和丰都江段的水温是整个长江上游水温较高的区域[25],适合黄颡鱼的繁殖产卵。鱼类的习性和生境条件是决定鱼类产卵分布的关键因素,因而对于生境多样性的保护是鱼类资源多样性保护的前提。
3.3 鱼卵致死原因
影响鱼卵正常发育的关键因素有水温、溶解氧、流速等[26-29]。首先,水温是影响鱼卵发育的关键因子,适合家鱼产卵的水温为18~24 ℃,且当水温低于18 ℃时,产卵停止,所以可以把18 ℃作为家鱼产卵的下限温度[26]。六月份,朱沱站的平均水温约为22 ℃,在此温度条件下,鱼卵的孵化成功率(以草鱼为例)为80%,死亡率最高的鱼卵发育阶段为原肠期。产卵期长江上游水体的溶解氧不会成为鱼卵致死的制约因素[30]。因水温对于鱼卵死亡率的贡献程度远没有实测值那么高,由此课题组认为,除了水温和溶解氧之外,被长期忽略的水流对鱼卵的作用亦是导致鱼卵死亡的另一个重要因素,如高流速条件下水流的紊动导致的剪切作用力,鱼卵在流场中的持续旋转、启停等运动过程中能量从水流向鱼卵转移导致的鱼卵死亡[31]。其次,受精卵是漂流性鱼卵正常发育的前提,而漂流性鱼卵通常是体外受精,产卵期长江上游水流高流速和复杂流态的水流特性,亦会造成鱼卵难以完全受精,进而导致鱼卵败育。此外,也不能完全排除水体中的泥砂、有机物、新型污染物[32]等可能导致鱼卵败育的原因。
4 结论和建议
根据2019年6-7月长江上游宜宾-丰都江段卵苗数量、形态特征观测和种类鉴定结果,分析了长江上游卵苗的时空分布以及鱼卵死亡的原因,得到主要结论如下。
(1)各采样点鱼卵与鱼苗的资源量估算处于相同数量级,南溪和朱沱采集的鱼卵数量较少但处于发育早期的鱼卵比例高,而涪陵和丰都采集的鱼卵大多处于发育中后期。江津上游鱼类物种多样性远多于下游的涪陵和丰都,如中华金沙鳅等珍稀特有鱼类主要分布在江津以上江段,但宜昌鳅鮀和壮体沙鳅的仔鱼仅在涪陵采样点发现。
(2)涨水是漂流性卵鱼类产卵的必要条件,南溪-宜宾江段流量涨幅超过10%且水位日涨幅>0.5 m时既会出现大规模产卵,而涪陵和丰都江段则需要更大的流量(>40%)和水位涨幅(>2 m)刺激才可能出现大规模产卵。
(3)采集的鱼卵中约3/4死亡,高流速和高紊动的水流特性可能是导致鱼卵死亡的另一个重要原因,水流对鱼卵孵化过程的影响亟需进一步的研究。
