干扰乙酰辅酶A合成酶2对三阴性乳腺癌MDA-MB-468细胞增殖及凋亡的影响
2021-09-18傅聪周月鹏尹超云凌锐浦希汤翔陈德玉
傅聪,周月鹏,尹超云,凌锐,浦希,汤翔,陈德玉
(江苏大学附属医院 1. 肿瘤治疗中心; 2. 肿瘤学实验室; 3. 血管外科, 江苏 镇江 212001)
三阴性乳腺癌即雌激素受体、人表皮生长因子受体-2和孕激素受体均阴性,是一种具有高侵袭性、高耐药性、进展快、预后差且缺乏明确治疗靶点的亚型[1],占所有病例的10%~15%,患者死亡率较其他类型高2.6倍[2-3]。研究表明,乙酰辅酶A在恶性实体肿瘤细胞代谢中发挥重要调控作用。异常高表达的乙酰辅酶A合成酶2(acetyl-CoA synthetase 2,ACSS2)作为关键调控酶,参与食管癌细胞缺氧诱导的放疗抵抗,抑制肿瘤细胞凋亡[4];宫颈癌细胞中ACSS2可促进营养缺乏(1%血清培养条件)诱导的癌细胞迁移和侵袭[5];同样,肾细胞癌患者肾脏病理组织中ACSS2高表达,且与肿瘤进展、侵袭行为及不良预后密切相关[6]。然而ACSS2在三阴性乳腺癌中具体作用尚不清楚。因此,本实验以MDA-MB-468细胞为研究对象,基于siRNA技术研究ACSS2在三阴性乳腺癌细胞增殖以及凋亡发生中的作用,探讨磷酸肌醇3-激酶(phosphatidylinositide 3-kinase,PI3K)/蛋白激酶B(protein kinase B,PKB,又称 AKT)信号通路调控乳腺癌细胞凋亡的作用。
1 材料和方法
1.1 细胞、主要试剂及仪器
人三阴性乳腺癌MDA-MB-468细胞株购自上海吉凯基因化学技术有限公司;人正常乳腺上皮MCF-10A细胞株购自中国科学院上海生命科学研究院细胞资源中心。
L15基础培养基和胎牛血清(美国Gibco公司);MCF-10A细胞专用培养基(瑞士Lonza公司);RNA提取试剂盒(上海奕杉生物科技有限公司);实时荧光定量PCR(qRT-PCR)试剂盒和逆转录试剂盒(南京诺唯赞生物科技股份有限公司);鼠抗人ACSS2抗体(美国Santa Cruz公司);兔抗人PI3K、p-PI3K、AKT、p-AKT、Ki-67、cleaved-caspase-3、Bcl-2、Bax抗体(美国Cell Signaling Technology公司);HRP标记的抗小鼠或抗兔二抗(北京索莱宝科技有限公司);脂质体Lipofectamine 2000(美国Invitrogen公司);CCK-8以及Annexin V-FITC/7-AAD凋亡试剂盒(杭州联科生物技术有限公司);针对ACSS2的特异性干扰RNA(siRNA-ACSS2)及无序序列的阴性对照(siRNA-NC)由上海吉凯基因技术公司设计并合成,siRNA-ACSS2序列: 5′-UAUGCUUGGUGACAGGCU CAUCUCC-3′,siRNA-NC序列:5′-GCGACGAUCUGC CUAAGAUdTd -3′。
细胞恒温培养箱、超净工作台(德国Heraeus公司);化学发光凝胶成像机(上海勤翔科学仪器有限公司);Mx3000p 荧光定量 PCR扩增仪(美国 Stratagene公司);紫外分光光度仪(美国Thermo公司);多功能酶标仪(美国Bio Tek公司);流式细胞分析仪(美国BD公司)。
1.2 细胞培养
三阴性乳腺癌MDA-MB-468细胞株用含10%胎牛血清、100 U/mL青霉素及 0.1 mg/mL链霉素的L15基础培养基,正常乳腺上皮MCF-10A细胞株用专用培养基,均于 37 ℃、5% CO2的培养箱中培养。
1.3 qRT-PCR检测ACSS2 mRNA表达
按照RNA提取试剂盒说明书提取细胞RNA,并使用紫外分光光度仪分析其纯度及浓度。按照试剂盒说明书操作方法将RNA逆转录为cDNA。ACSS2上游引物:5′-GGATTCCAGCTGCAGTCTTC-3′,下游引物:5′-CAGCCAGCTCCTTCAGGTT-3′;β-肌动蛋白上游引物:5′-TCACCCACACTGTGCCCATCTACGA-3′,下游引物:5′-CAGCGGAACCGCTCATTGCCAATGG-3′。按照反应试剂盒说明书在冰上配制反应体系 20 μL:10 μL TB Green PremixExTaq(Tli RNaseH Plus)、上下游引物各0.4 μL、0.4 μL ROX Reference Dye、2 μL DNA模板和6.8 μL灭菌水。95 ℃预变性 30 s;95 ℃ 10 s,60 ℃ 30 s,共循环 40 次;退火:95 ℃ 15 s,60 ℃ 60 s。循环结束后记录各样品Ct值结果,用2-ΔΔCt法计算相对表达水平,以β-肌动蛋白为内参照基因。
1.4 细胞分组与转染
细胞分组如下:空白对照组(正常培养)、阴性对照组(转染siRNA-NC)和实验组(转染siRNA-ACSS2)。选取生长状态良好且处于对数生长期MDA-MB-468细胞;转染前一天,将2×105的细胞种于6孔板,待密度达到40%~50%时,加入5 μL siRNA(阴性对照组加入siRNA-NC;实验组加入siRNA-ACSS2)和 5 μL Lipo2000TM混合液,补齐培养基至最终体积为 2 mL/孔,孵育8 h;更换为完全培养基继续培养24 h,进行后续实验。
1.5 蛋白质印迹法检测ACSS2、PI3K/AKT通路相关蛋白、增殖相关蛋白Ki-67和凋亡相关蛋白表达
取“1.4”分组转染细胞,分别以2×105个/孔密度种于6孔板,待24 h细胞贴壁,弃上清液,PBS 清洗2次;每孔加入50~100 μL蛋白裂解液(含蛋白酶及磷酸酶抑制剂),置于冰上孵育5 min,每15 min将EP管混匀,重复3次;4 ℃,12 000×g离心30 min,收集上清液;加入4×上样缓冲液,混匀后 100 ℃煮10 min。取5~10 μL上样,60 mA恒流电泳30 min;利用湿转法100 V恒压转移30~100 min,将蛋白转移至PVDF膜;用5%牛血清白蛋白封闭1 h;TBST洗3次;加入一抗ACSS2、PI3K、p-PI3K、AKT、p-AKT、Ki-67、cleaved-caspase-3、Bcl-2、Bax(TBST稀释,均为1 ∶1 000),4 ℃孵育过夜,TBST洗膜;加入对应HRP标记的相应二抗(TBST稀释,均为1 ∶1 000) ;室温孵育1 h;TBST洗膜3次;曝光显影,Image J软件用于灰度分析。以β-肌动蛋白作为内参,计算目的蛋白相对表达量。
1.6 CCK-8法检测细胞增殖能力
取“1.4”分组转染细胞,分别以2×103个/孔密度种于96孔板。继续培养24、48和72 h;加入5 mg/mL的CCK-8试剂10 μL并混匀,继续培养 2 h;波长450 nm处测量光密度(D)值,并计算细胞增殖活力。细胞增殖活力(%)=(实验组D值-空白组D值)/(对照孔D值-空白组D值)×100%。实验重复3次。
1.7 流式细胞术检测细胞凋亡水平
取“1.4”分组转染细胞,分别以2×105个/孔种于6孔板,孵育至细胞贴壁后进行由Lipo2000TM介导的siRNA转染。转染后24 h重新计数,以每管收集3×105个细胞置入流式管中,按照Annexin V-FITC/7-AAD凋亡试剂盒说明书,将5×缓冲液稀释成1×工作液,取300 μL重悬细胞,每管加入 5 μL Annexin V-FITC和10 μL 7-AAD,室温避光孵育10 min;选择最低的上样速度于流式细胞分析仪检测。每组重复3次。
1.8 统计分析
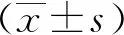
2 结果
2.1 ACSS2在三阴性乳腺癌MDA-MB-468细胞中呈高表达
结果显示,三阴性乳腺癌MDA-MB-468细胞中ACSS2 mRNA和蛋白相对表达量均明显高于正常乳腺上皮MCF-10A细胞(t=15.14,21.2,P均<0.01)。见图1。
图1 qRT-PCR和蛋白印迹法分别检测MDA-MB-468和MCF-10A细胞中ACSS2 mRNA和蛋白表达
2.2 鉴定siRNA干扰ACSS2效果
qRT-PCR和蛋白印迹法结果显示,与阴性对照组相比,实验组中ACSS2mRNA和蛋白表达量明显下降(t= 31.73,26.88,P均<0.01);空白对照组与阴性对照组间差异无统计学意义(P均>0.05)。见图2。

图2 qRT-PCR和蛋白印迹法鉴定ACSS2干扰效果
2.3 siRNA-ACSS2抑制MDA-MB-468细胞增殖并促进其凋亡
CCK-8法结果显示,干扰ACSS2处理24、48、72 h后,实验组细胞增殖能力较阴性对照组明显下降(t=5.056、5.324、13.240,P均<0.01);空白对照组与阴性对照组间差异无统计学意义(P均>0.05)。见图3。流式细胞术结果显示,实验组细胞凋亡率较阴性对照组明显上升(t= 9.215,P<0.01);空白对照组与阴性对照组间差异无统计学意义(P均>0.05)。见图4。
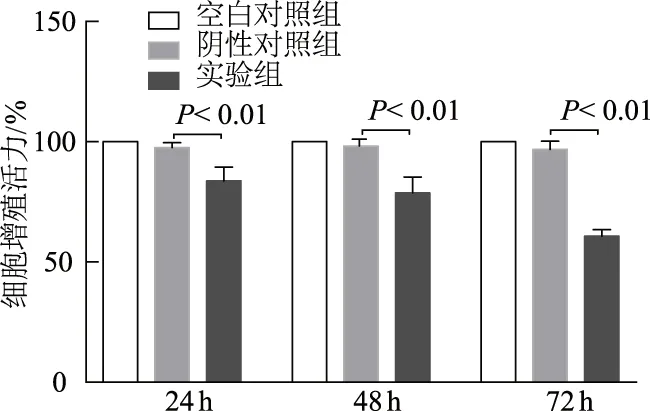
图3 CCK-8法检测各组MDA-MB-468细胞增殖能力变化

图4 流式细胞术检测各组MDA-MB-468细胞凋亡率变化
2.4 siRNA-ACSS2降低细胞增殖相关蛋白表达并增加PI3K/AKT信号通路及凋亡相关蛋白表达
蛋白印迹法结果显示干扰ACSS2表达后,PI3K/AKT信号通路相关蛋白p-PI3K、p-AKT表达量较阴性对照组均明显下降(P均<0.01);Ki-67和Bcl-2蛋白表达量也较阴性对照组均明显下降(P均<0.01),而cleaved-caspase-3和Bax蛋白表达量明显上升(P均<0.01);空白对照组与阴性对照组间无明显差异(P均>0.05)。见图5。

a: P<0.01,与阴性对照组比较图5 蛋白印迹法检测各组MDA-MB-468细胞相关蛋白表达
3 讨论
现有研究发现,肿瘤微环境中的乙酸盐可经肿瘤细胞中异常高表达的ACSS2利用,催化形成乙酰辅酶A,通过调节组蛋白乙酰化修饰,进一步激活调控细胞增殖、凋亡、自噬等特定基因表达或激活相关信号通路[7]。前期研究表明,ACSS2蛋白在胃癌[8]、宫颈癌[5]、食管癌[4]、肾癌[6]中呈不同程度高表达。本研究表明,无论是转录还是翻译水平,三阴性乳腺癌MDA-MB-468细胞中ACSS2表达明显高于正常乳腺上皮MCF-10A细胞,与上述研究相一致。进一步证实ACSS2利用糖酵解途径代谢产生的乙酸盐合成乙酰辅酶A,并在一系列酶促反应下合成脂肪酸作为细胞能量来源及合成原料[9],而正常细胞的能量代谢方式并不依赖ACSS2。
Zhang等[6]研究表明,下调ACSS2蛋白表达可抑制肾细胞癌细胞增殖、迁移和侵袭。Wang等[10]研究表明,在肝癌细胞中,细胞核内ACSS2第一个转录起始位点表达增加,使组蛋白释放的醋酸酯去乙酰化,增加核糖体产生,进而促进肿瘤细胞增殖、侵袭和转移。本研究表明,靶向干扰ACSS2表达可抑制三阴性乳腺癌MDA-MB-468细胞增殖,并下调增殖相关Ki-67蛋白的表达水平,从而可能进一步延缓肿瘤发展,可能与细胞核内组蛋白去乙酰化有关。由此提示,ACSS2沉默可能与抑制乳腺癌细胞生长相关。
凋亡是细胞程序性停止生长和分裂的过程,其调控失败,可致体内受损细胞累积,导致癌症发生[11]。细胞凋亡的启动依赖于一系列的半胱氨酸-天冬氨酸蛋白酶的激活,其中cleaved-caspase-3是具有活性的执行蛋白酶,其可激活内切酶导致DNA片段化和细胞骨架破坏等一系列引起细胞凋亡事件发生[12]。此外,细胞凋亡过程还涉及Bcl-2家族成员Bax激活,而Bcl-2可在线粒体水平对抗凋亡刺激,与Bax形成异源二聚体,进而抑制凋亡,对细胞起保护作用。Yao等[13]研究显示,植物提取物Wikstromol可在乳腺癌MDA-MB-231细胞中通过抑制Bcl-2、促进Bax蛋白表达以及激活Caspase-3诱导三阴性乳腺癌细胞凋亡。同样,Du等[14]研究发现,敲减长链非编码RNA DLX6的反义RNA(DLX6-AS1)可通过抑制Bcl-2蛋白表达、促进Caspase-3蛋白活化诱导肿瘤细胞凋亡,而过表达DLX6-AS1则相反。本研究发现,靶向下调ACSS2表达后乳腺癌细胞凋亡率明显上升,并使Bax和cleaved-caspase-3蛋白表达增加,Bcl-2蛋白表达减少。由此说明,ACSS2缺失可诱导三阴性乳腺癌细胞凋亡。
研究发现PI3K/AKT信号转导通路在多种恶性肿瘤中异常激活,如肝癌[15]、胰腺癌[16]及胃肠道肿瘤[17-18]等,其与肿瘤发生、发展密切相关,调控细胞增殖、凋亡、转化、黏附和胞外基质降解等功能,参与肿瘤的全程。Yao等[13]研究显示,Wikstromol可通过降低NF-κB的转录水平从而抑制PI3K/AKT通路诱导三阴性乳腺癌细胞凋亡。Niu等[19]研究发现,在三阴性乳腺癌MDA-MB-468细胞中,蚕茧中的大分子蛋白Sericin通过PI3K/AKT途径抑制细胞增殖,诱导细胞周期停滞和促进细胞凋亡。本研究结果显示,在MDA-MB-468细胞中,靶向干扰ACSS2可通过磷酸化PI3K和AKT蛋白,活化PI3K/AKT信号通路促进乳腺癌细胞凋亡。目前研究显示,抑制ACSS2表达可降低乙酰辅酶A活性和组蛋白乙酰化,从而抑制癌细胞侵袭转移[20]。然而在三阴性乳腺癌MDA-MB-468细胞中,ACSS2、组蛋白乙酰化与细胞增殖凋亡间的关系尚未明确,有待后续研究。本研究通过体外靶向下调ACSS2探究其生物学特性,但由于体外实验无法完全模拟体内环境,ACSS2生物学特性有待在体内以及组织样本中进一步验证。
综上,ACSS2在三阴性乳腺癌MDA-MB-468细胞中高表达;干扰ACSS2表达可抑制肿瘤细胞增殖,其可能通过PI3K/AKT信号通路促进乳腺癌细胞凋亡。
