哈蟆油与青蛙油蛋白质组学差异分析
2021-09-04董颖张鹤张玉尧潘黛安魏星华赵大庆鲁西南医院有限公司山东聊城5300长春中医院大学附属医院长春300吉林省科普服务中心长春300
董颖,张鹤,张玉尧,潘黛安,魏星华,赵大庆(.鲁西南医院有限公司,山东 聊城 5300;.长春中医院大学附属医院,长春 300;3.吉林省科普服务中心,长春 300)
哈蟆油为蛙科动物中国林蛙Rana temporaria chensinensisDavid 雌蛙的输卵管,为滋补药材,具有补肾益精、健脾益胃、滋阴补肾、润肺生津等功效。中国林蛙的输卵管呈不规则弯曲,位于腹腔中下位两侧,全长呈现粗细不均匀,近肺根段较细,中段渐渐加粗,近子宫段输卵管较粗且略扁,其长度一般为体长的4 ~8 倍,质量为体质量的20%~26%[1];呈现乳白色或黄白色,干品偶带有灰白色薄膜状的干皮,呈脂肪样光泽,手摸有滑腻感,质硬且易断;有特殊的油脂气味和腥味,味微甘,嚼之有黏滑感;膨胀度大于55;其主要成分为蛋白质,含量约占50%以上[2]。常见的伪品黑斑蛙的输卵管(青蛙油)外观性状为不规则弯曲,中间灰黑色瓣膜,管径均一,透明;白色或黄白色,有脂肪样光泽;质脆易碎,手摸之稍有油脂状润滑感;味辛辣,嚼之麻嘴、黏牙;膨胀度大于55;其主要成分为蛋白质,含量约为47.5%[3]。在目前的研究中,伪品青蛙油与正品哈蟆油性状上相似,不易区别,但通过显微、薄层色谱等方法可以进行鉴别[4]。至今研究显示哈蟆油中特异性的成分仅有1-甲基海因,但目前在青蛙油中也发现含有该成分[5]。本研究采用iTRAQ技术和质谱分别建立和比较哈蟆油和青蛙油的蛋白质组数据库,筛选两个种属蛙在冬眠前输卵管中蛋白组成的差异。
1 材料
1.1 样品采集
哈蟆油的基源动物为中国林蛙长白山亚种,保护地域为吉林省长白山区域。林蛙采于吉林省蛟河市前进乡(东经127°40′18.4′,北纬43°54′20.9′)、永吉县北大湖镇(东经126°30′14′,北纬43°27′53′)、舒兰市上营镇(东经127°15′43.9′,北纬44°8′25.2′)、桦甸市红石砬子镇(东经127°6′26.5′,北纬42°57′17.7′)、集安市清河村(东经125°55′58.2′,北纬41°25′59.5′)和安图县三道湾镇(东经129°10′19.5′,北纬43°9′29.2′),海拔在500 ~1000 米。黑斑蛙(青蛙)采自于吉林省永吉县北大湖镇(东经126°30′14′,北纬43°27′53′)和吉林市(东经126°25′17′,北纬44°4′40′)两个地区,海拔在100 ~300 米。中国林蛙和黑斑蛙均经长春中医药大学药学院肖井雷副教授鉴定为正品。
1.2 仪器与试药
Q Exactive 质谱仪(Thermo 公司);AKTA Purifier 100 蛋白纯化仪、数控600V 电泳仪(通用GE Healthcare 公司);5430R 型低温高速离心机、真空离心浓缩仪(德国Eppendorf 公司);胰蛋白酶(Promega 公司);iTRAQ Reagent-8plex Multiplex Kit(美国AB SCIEX 公司);丁香酚(Sigma,纯度:98%,批号:E2069)。
2 方法[6-7]
2.1 林蛙和黑斑蛙的输卵管(蛙油)摘取
分别取活蛙,沿腹部正中线解剖,取出输卵管,去除筋膜、肉、卵等杂物,用蒸馏水反复清洗至呈乳白色或黄白色,置于阴凉干燥处晾干,研钵研磨成粉末。哈蟆油样品:5 个产地各取10 只混合后准确称量100 mg 后混匀;青蛙油样品:两个产地分别取10 只混合,准确称量100 mg,混匀。
2.2 蛙油中蛋白质的提取和含量测定
两个蛙油样品各准确称取10.0 mg,分别加入1 mL SDT 裂解液,匀浆后沸水浴5 min,超声破碎10 min,14 000 g 室温离心45 min,取上清液,-80℃保存。考马斯亮蓝法测定两个蛙油样品中蛋白质的含量,哈蟆油蛋白质的质量浓度分别为8.83 μg·μL-1和7.09 μg·μL-1,青蛙油蛋白质的质量浓度为6.59 μg·μL-1。
2.3 iTRAQ 定量分析
每份样品各取250 μg,胰酶消化,酶解后的肽段用8 标iTRAQ 试剂中的113、114 和115 进行标记。采用二维液相(纳升流速HPLC 液相系统Easy nLC 和强阳离子交换液相色谱)进行分离,每份样品经分离后用Q-Exactive 质谱仪进行分析[8]。
2.4 哈蟆油与青蛙油差异蛋白质筛选
对哈蟆油蛋白质组数据库中1220 个蛋白质组在样本哈蟆油113(A)/REF 和114(B)/REF与青蛙油115(C)/REF 中的定量信息进行差异分析,筛选条件为比值>1.5(或比值<0.67),且P<0.05。
2.5 生物信息学分析
2.5.1 Gene Ontology(GO)功能注释分析 在GO 注释过程中,Blast 2 GO 通过比较目标序列和比对序列的相似性、GO 条目来源的可靠性和GO有向无环图的结构,将map 过程中提取的GO 功能条目中符合条件的条目注释给目标蛋白质。
2.5.2 WEGO 注释分析 利用WEGO 软件对鉴定到的差异蛋白质的细胞学组件、生物学途径和分子功能完成GO 注释,比较青蛙油(C/REF)与哈蟆油(/REF)差异蛋白质进行WEGO 分析。对GO 2 层次进行汇总。
2.5.3 KEGG 通路注释 利用KAAS(KEGG automatic annotation server)数据库将目标蛋白质序列与KEGG GENES 数据库中的两栖纲蛋白质序列进行比对,通过同源/相似蛋白质的KO 号注释到相关KEGG 通路上。
3 结果
3.1 青蛙油与哈蟆油差异蛋质白质筛选结果
对青蛙油与哈蟆油中蛋白质定量的数据进行差异比较,筛选出差异表达蛋白质总数为55 个,其中上调的蛋白质数为40 个(见表1),下调的蛋白质数为15 个(见表2)。
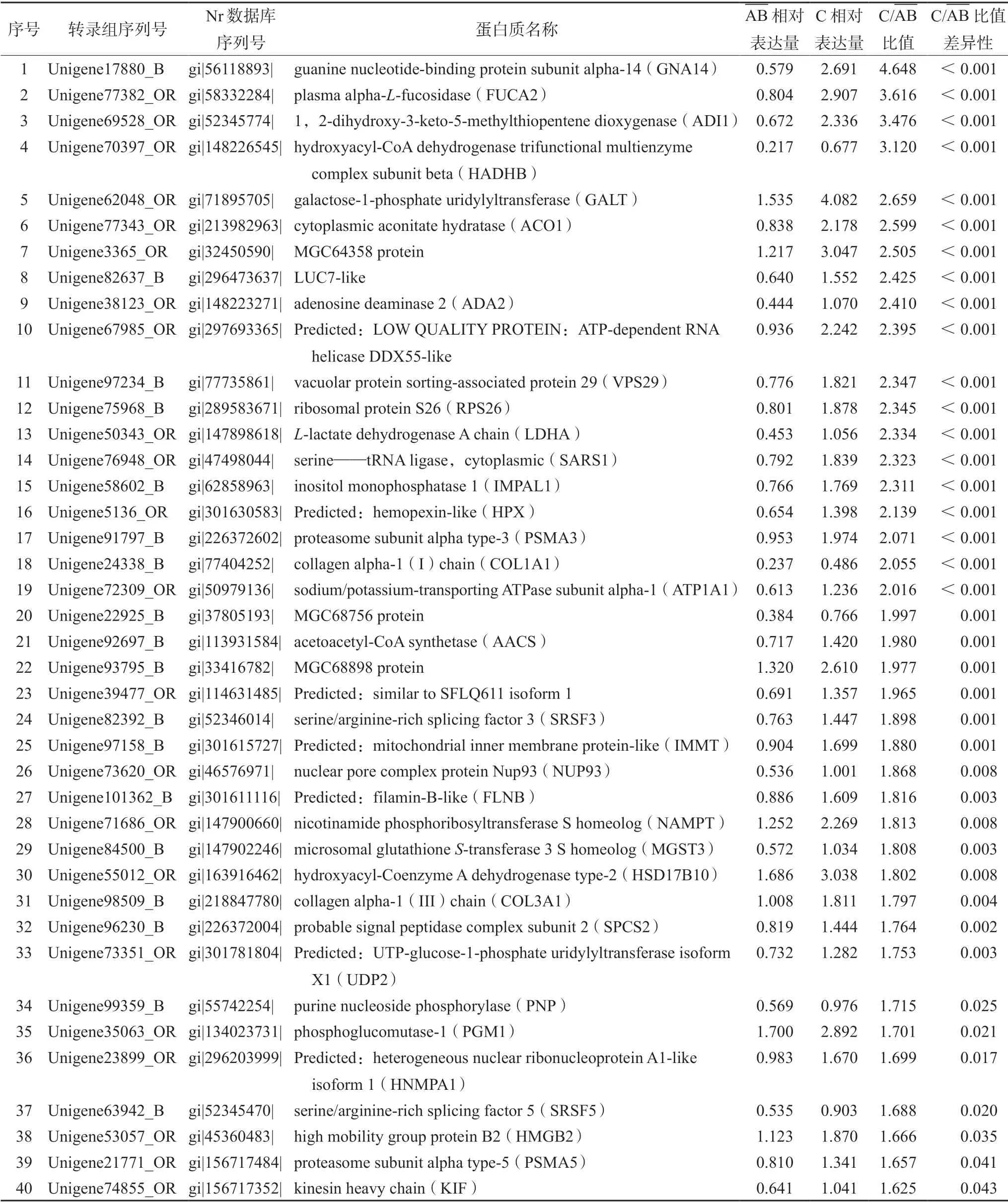
表1 青蛙油与哈蟆油相比上调的蛋白质Tab 1 Up-regulated proteins in the proteomics database of OCB compared with that of ORN
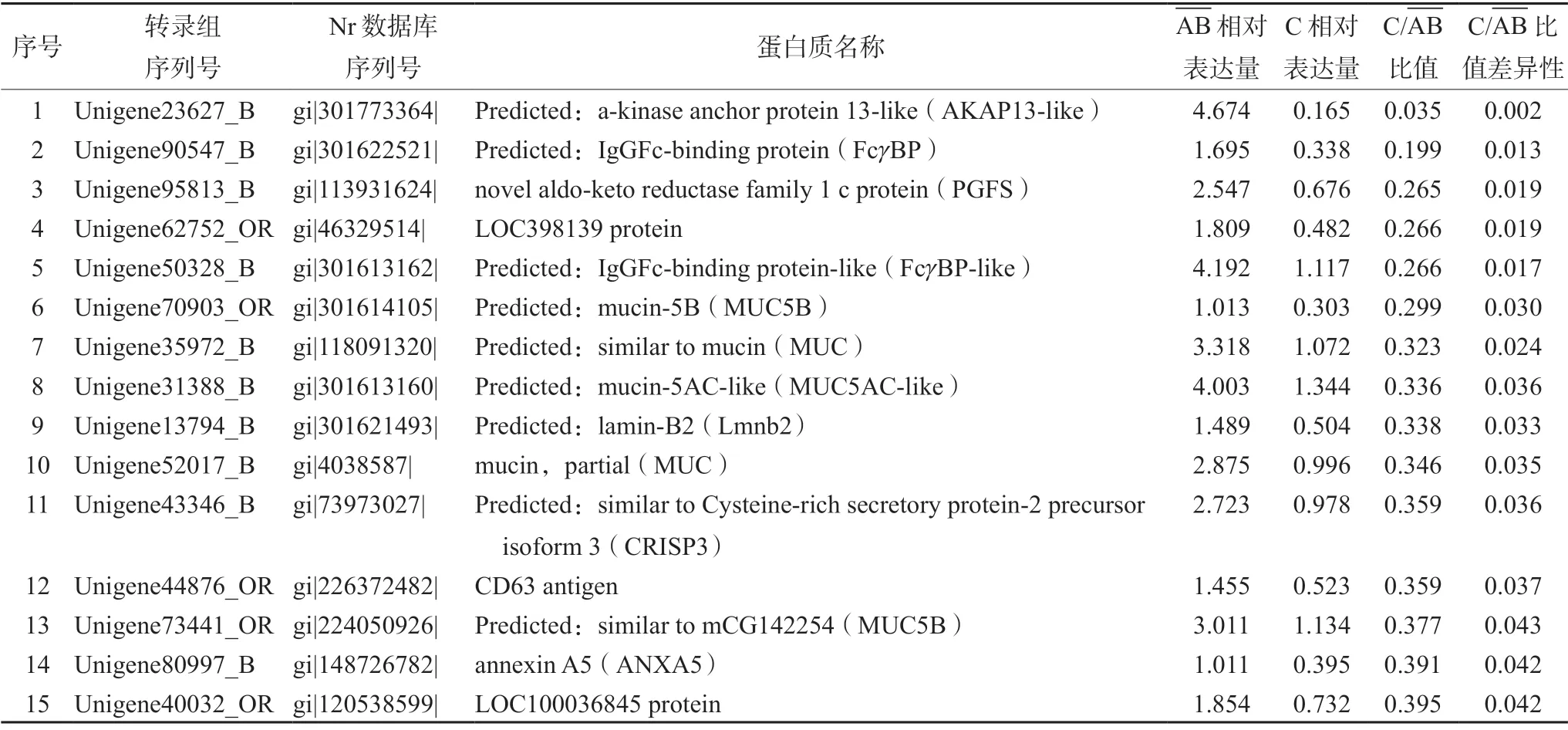
表2 青蛙油与哈蟆油相比下调的蛋白质Tab 2 Down-regulated proteins in the proteomics database of OCB compared with that of ORN
3.2 青蛙油中含量较高的差异蛋白质(上调蛋白)
参与能量代谢过程的蛋白质包括羟酰基辅酶A脱氢酶三官能多酶复合体亚基β(HADHB)、乙酰辅酶A 合成酶(AACS)、乌头酸水合酶(ACO1)、烟酰胺磷酸核糖转移酶s 同源体(NAMPT)和羟酰辅酶A 脱氢酶Ⅱ型(HSD17B10)等。参与糖代谢的蛋白质包括半乳糖-1-磷酸尿苷酰转移酶(GALT)、血浆a-L-岩藻糖苷酶(FUCA2)和磷酸葡萄糖变位酶(PGM1)等。参与应激反应的蛋白质包括鸟嘌呤核苷酸结合蛋白a 亚基14(GNA14)、L-乳酸脱氢酶A 链(LDHA)、血红素结合蛋白(HPX)和蛋白酶体亚基α-3 型(PSMA3,PSMA5)等。参与生长发育相关的蛋白质包括LUC7-like、核孔复合物蛋白(NUP93)和高速泳动族蛋白B2(HMGB2)等。参与生物合成的蛋白质包括核糖体蛋白(RPS26)、不均一核核糖核蛋白质A1 样亚型1(HNMPA1)和丝氨酸转运RNA 连接酶(SARS1)等。参与细胞骨架形成的蛋白质包括细丝蛋白(FLNB)和驱动蛋白重链(KIF)等。参与氧化还原的蛋白质包括1,2-二羟基-3-酮-5-甲基硫戊烯双加氧酶(ADI1)和微粒体谷胱甘肽s-转移酶3 S 同源体(MGST3)等。参与免疫调节相关的蛋白质包括腺苷脱氨酶2(ADA2)。
3.3 哈蟆油中含量较高的差异蛋白质(下调蛋白)
3.4 WEGO 注释分析结果
3.4.1 细胞组分分析 从图1细胞组分中可以看出青蛙油与哈蟆油比较差异蛋白质表达体现在细胞、细胞器、膜、大分子复合体、膜封闭腔和胞外区,分别占所有差异蛋白质的34%、23%、16%、16%、8%和3%。
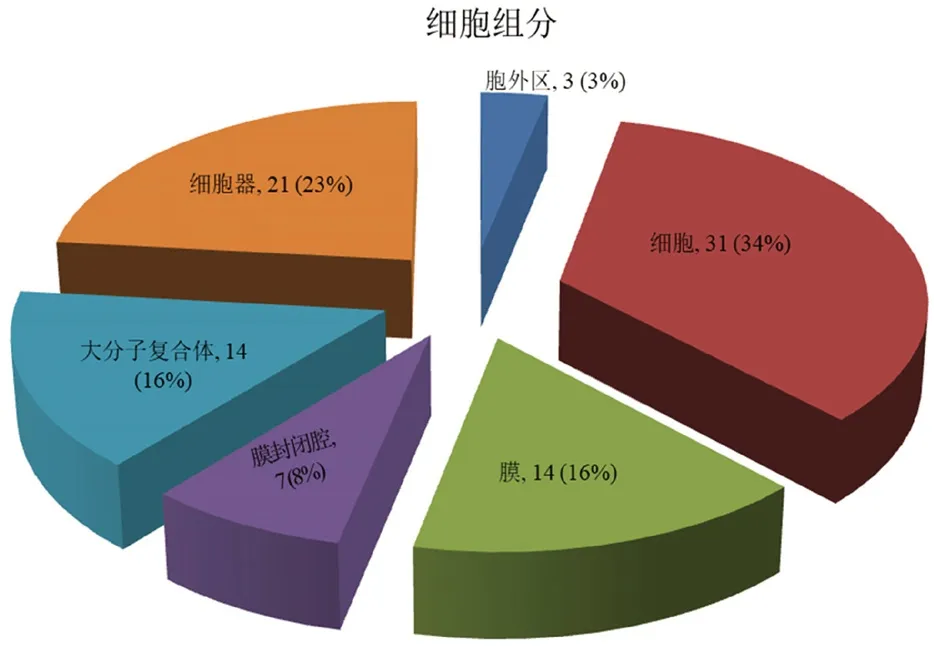
图1 差异蛋白质细胞组分分析Fig 1 Cellular components of differentially expressed proteins
3.4.2 分子功能分析 从图2分子功能中可以看出催化活性、结合功能、结构分子活性、酶调节活性、化学引诱物活性、载体活性、核酸结合转录因子活性、抗氧化活性和分子转导活性是青蛙油与哈蟆油差异蛋白质比较集中的9 类功能,其中催化活性和结合功能类蛋白质最多,分别占所有差异蛋白质的43%和42%。
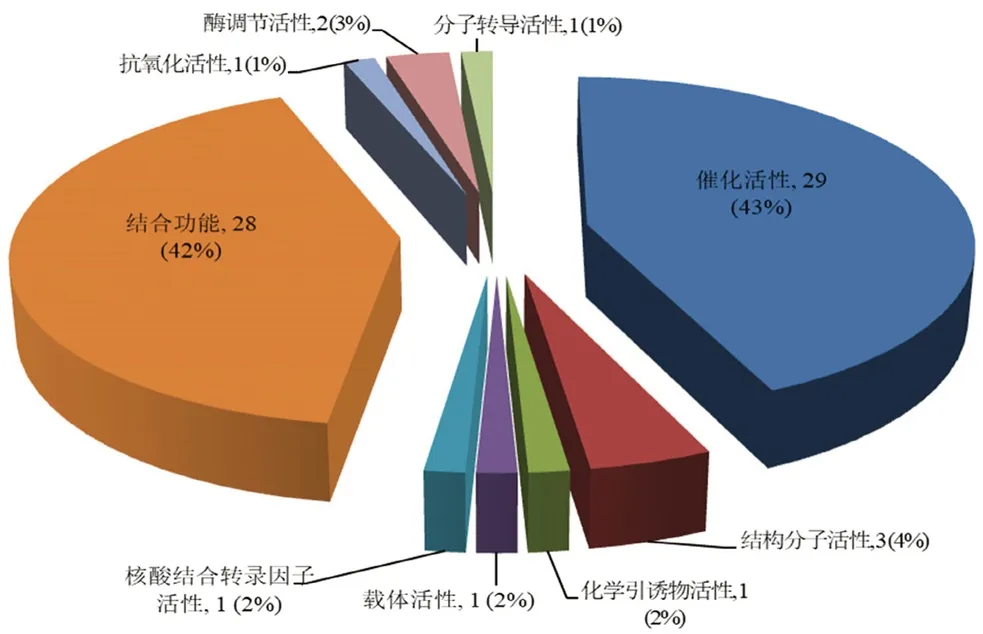
图2 差异蛋白质分子功能分析Fig 2 Molecular function of differentially expressed proteins
3.4.3 生物学途径分析 从图3生物学途径中可以看出繁殖、免疫系统过程、代谢过程、细胞过程、信号、多组织过程、发育过程、生长过程、位移运动、单组织过程、刺激反应、定位、多生物过程、生物调节和细胞成分组织/生物合成是青蛙油与哈蟆油差异蛋白质比较集中的15 个生物学途径,其中参与代谢过程、细胞过程、单组织过程和生物调节的蛋白质数量较多,分别占所有差异蛋白质的21%、19%、12%和10%。
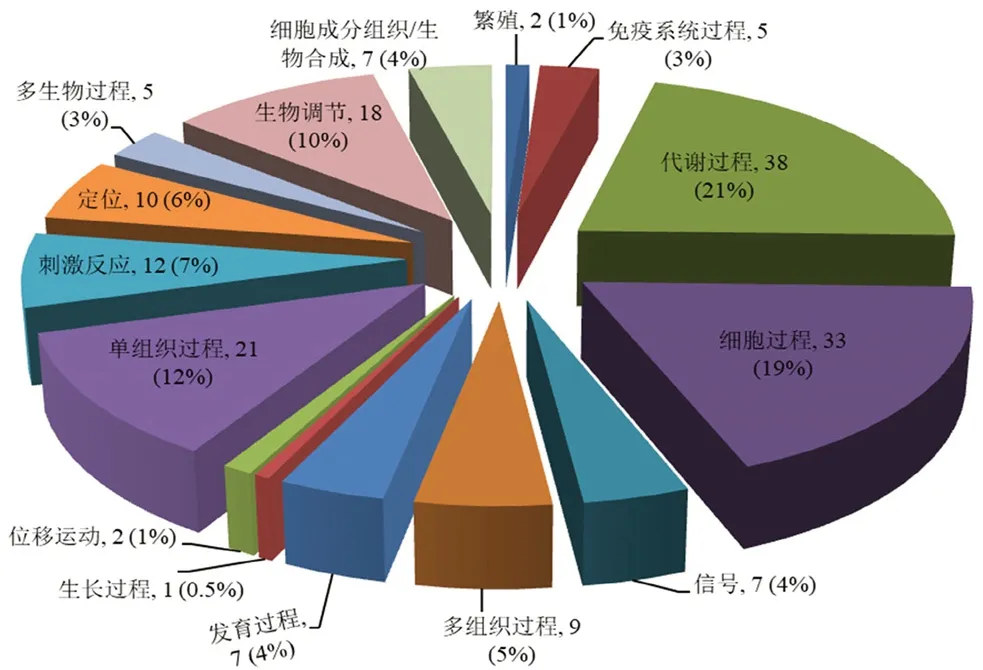
图3 差异蛋白质生物学途径分析Fig 3 Biological process of differentially expressed proteins
3.5 青蛙油与哈蟆油差异蛋白质KEGG 代谢通路分析
青蛙油与哈蟆油的差异蛋白质总数为55 个,其中29 个差异蛋白质map 到77 条KEGG 信号/代谢通路中;在这29 个差异蛋白质中,27 个上调蛋白map 到76 条KEGG 信号通路中,两个下调蛋白map 到8 条KEGG 信号通路中。
与哈蟆油蛋白质数据比,青蛙油中含量较高(即上调)的27 个差异蛋白质中钠/钾离子转运ATP 酶亚基α1 肽(ATP1A1)参与cGMP-PKG 信号通路(cGMP-PKG signaling pathway)、唾液分泌(salivary secretion)、心肌细胞肾上腺素能信号转导(adrenergic signaling in cardiomyocytes)、胰岛素分泌(insulin secretion)、甲状腺激素合成(thyroid hormone synthesis)等16 个代谢过程;PGM1 参与糖酵解/糖异生途径(glycolysis/gluconeogenesis)、磷酸戊糖途径(pentose phosphate pathway)、半乳糖代谢(galactose metabolism)、嘌呤代谢(purine metabolism)等7 个代谢过程;Ⅰ型胶原蛋白a1 链(COL1A1)参与焦点黏连(focal adhesion)、ECM 受体相互作用(ECM-receptor interaction)、血小板活化(platelet activation)等6 个代谢过程;ACO1 参与三羧酸循环(citrate cycle)、乙醛酸和二羧酸代谢(glyoxylate and dicarboxylate metabolism)、原核生物固碳过程(carbon fixation pathways in prokaryotes)、碳代谢(carbon metabolism)等6 个代谢过程;HADHB 参与脂肪酸降解(fatty acid degradation),脂肪酸伸长(fatty acid elongation),缬氨酸、亮氨酸和异亮氨酸降解(valine,leucine and isoleucine degradation),苯甲酸降解(benzoate degradation)和脂肪酸代谢(fatty acid metabolism)等5 个代谢过程;LDHA 参与糖酵解/糖异生途径(glycolysis/gluconeogenesis)、半胱氨酸和甲硫氨酸代谢(cysteine and methionine metabolism)、丙酮酸代谢(pyruvate metabolism)、丙酸代谢(propanoate metabolism)等4 个代谢过程。
在差异蛋白质富集分析中发现,半乳糖代谢、氨基糖和核苷酸糖代谢(amino sugar and nucleotide sugar metabolism)均富集到3 个差异蛋白质(GALT、PGM1、UDP2)。糖酵解/糖异生途径富集到两个差异蛋白质LDHA 和PGM1。嘌呤代谢(purine metabolism)富集到两个差异蛋白质PNP 和PGM1。半胱氨酸和甲硫氨酸代谢富集到两个差异蛋白质ADI1 和LDHA。缬氨酸、亮氨酸和异亮氨酸降解富集到两个差异蛋白质HADHB 和HSD17B10。
哈蟆油中含量较高(即下调)的两个差异蛋白质包括LOC398139 protein,CD63 antigen。其中LOC398139 protein 参与三羧酸循环(citrate cycle)、乙醛酸和二羧酸代谢(glyoxylate and dicarboxylate metabolism)、原核生物固碳过程(carbon fixation pathways in prokaryotes)、氧代羧酸代谢(2-oxocarboxylic acid metabolism)和氨基酸生物合成(biosynthesis of amino acids)等6 个代谢过程。CD63 antigen 参与溶酶体(lysosome)和癌症过程蛋白聚糖(proteoglycans in cancer)等两个代谢过程。
其他差异蛋白质在KEGG 代谢通路中未被注释到。
4 讨论与结论
4.1 青蛙油中含量较高的差异蛋白质与环境、生长发育的关系
成年黑斑蛙常栖息于稻田、池溏、湖泽、河滨、水沟内或水域附近的草丛中。一般10月开始冬眠,钻入向阳的坡地或离水域不远的田地、沟埂、塘埂土层30 ~40 cm 深处穴居冬眠,次年4月后出蛰[9]。冬眠前青蛙油中含量较高的差异蛋白质(即上调成分)与黑斑蛙平时的栖息环境、冬眠环境可能存在密切关系。
鸟嘌呤核苷酸结合蛋白(GNA14)是在膜结合受体和细胞内效应器之间起信号传递作用,可水解GTP 生成GDP,具有GTP 酶活性,在哺乳动物细胞中G 蛋白和其受体形成一个最普遍的信号系统,在哺乳动物的各种组织中均有表达,如脑、心脏、胚胎等组织。参与调节感官感知[10]、细胞生长、激素的分泌和释放[11]、防御物质的信号传导[12]及脂肪体中能源物质的使用[13]。有研究表明,在低氧或缺氧的胚胎中GNA14 高表达,可调节高血压和内皮细胞的功能[14-15]。因此认为GNA14 在蛙冬眠过程中调节机体机能,适应环境方面起到重要作用。
血红素结合蛋白(HPX)是肝脏合成的一种急性时相反应蛋白,在血浆中与游离的血红素结合能力最强,广泛参与机体多种生理和病理过程。机体内游离的血红素是有害的,可以破坏脂质双分子层,促进低密度脂蛋白转化成具有细胞毒性的氧化产物[16],还可以插入红细胞膜,缩短红细胞的寿命,加速溶血的发生[17],HPX 可以结合和转运有毒的游离血红素至肝脏而发挥解毒作用。Grinberg 等[18]研究发现HPX 与血红素结合后,血红素诱导的氧自由基明显减少,过氧化活性下降80%~90%,脂质过氧化活性也显著降低,进而达到抗氧化的作用。目前研究结果显示HPX 还具有维持体内铁的稳态[19]、保护神经[20]等功能。在黑斑蛙冬眠过程中,高含量的HPX降解机体产生的毒素-血红素,同时起到抗氧化的作用。
羟酰基辅酶A 脱氢酶三官能多酶复合体亚基β(HADHB)是一种具有多功能的膜结合型蛋白酶,可以催化3 种酶活性(长链烯酰辅酶A 水合酶、长链3-羟烷基辅酶A 脱氢酶和长链3-酰基辅酶A 硫解酶),参与脂肪酸和脂质代谢过程,在线粒体中参与长链脂肪酸的β氧化过程产生能量[21-22]。ACO1 也称乌头酸酶,广泛分布于动植物和微生物中,在肝脏、肾脏、肌肉、前列腺中含量特别高。在三羧酸循环中,ACO1 为催化柠檬酸和异柠檬酸之间相互转换的辅酶[23]。GALT具有催化、连接活性,参与半乳糖代谢过程[24]。在KEGG 代谢通路中, 笔者发现ATP1A1、PGM1、ACO1、HADHB、GALT 和LDHA 是青蛙油中重要的差异蛋白质,主要参与体内的各种糖代谢、糖异生、氨基酸代谢、脂肪代谢等代谢过程。
综合以上青蛙油中差异的蛋白结构、活性特点可以看出,部分蛋白质是参与胚胎细胞生长、发育的重要物质基础,部分蛋白质在蛙冬眠过程中提供能量,是维持生命活动的重要成分,为冬眠期和翌年的排卵期做准备。
4.2 哈蟆油中含量较高的差异蛋白质的特点与环境、生长发育的关系
中国林蛙冬眠期主要在水下,这个特点是中国林蛙所特有的,其他种类的蛙一般都在泥土中冬眠。同时在东北中国林蛙最早结束冬眠,比黑斑蛙、蟾蜍等蛙类早20 ~30 d。本文通过比较青蛙油与哈蟆油蛋白质组学的差异,发现哈蟆油中蛋白成分差异主要为a-激酶锚定蛋白13 类似蛋白、免疫球蛋白Fc 段连接蛋白和黏蛋白,且此3 类成分在哈蟆油中表达量较高,对哈蟆油的功能活性起到至关重要的作用。
a-激酶锚定蛋白13 类似蛋白(AKAP13-like),AKAP13 包含两个重要的结构域(PKA 结构域和GEF 结构域),在许多信号传导中发挥着重要的作用,能激活GTP 酶Rho,水解GTP 为GDP。AKAP13 可将PKA、PKD、PKC 多蛋白复合物锚定于细胞骨架肌动蛋白[25-26],在PKD 和Rho 通路中起着整合cAMP 信号的作用;AKAP13将PKD 与其上游激活因子PKC 连接,PKC 磷酸化PKD,使其激活,而PKA 磷酸化AKAP13,促进激活的PKD 从AKAP 复合物平台上游离出来。AKAP13 在多个器官和组织中高表达,如脾、胸腺、外周血白细胞、骨路肌和睾丸;胎盘和肺中适度表达;而在肝、脑、小肠、结肠和前列腺癌中没有或者少量表达[27]。目前研究显示AKAP13 通过GEF 作用,参与心肌肥大的形成[28]。邢修业[29]研究AKAP13 在小鼠卵泡发育中的作用,发现AKAP13 基因敲除杂合型小鼠的生殖能力降低,表明AKAP13 可能通过作用于FSHR 信号通路下游效应分子参与并影响FSHR信号通路,进而在小鼠卵泡发育中起至关重要的作用。免疫球蛋白Fc 段连接蛋白(IgGFc binding protein,FcγBP)主要在胎盘和结肠的上皮细胞表达,作为黏膜的组成部分,维持黏膜的结构。FcγBP 类似黏蛋白,具有黏蛋白的结构特点,同时具有免疫球蛋白连接的活性,在黏膜免疫系统和炎症中具有重要的作用[30]。Kobayashi 等[31]报道FcγBP 存在于血清中,在各种自身免疫性疾病的患者中高水平表达。黏蛋白(mucin)是分布在各种管腔内表面的高分子量糖蛋白,对上皮细胞起到重要的保护和润滑的作用,黏蛋白中的糖链在维持蛋白质的稳定、抵抗蛋白酶水解、防止抗体识别等方面发挥重要的作用[32]。近年来,发现黏蛋白还参与细胞增殖的调控、受精,上皮细胞的分化、更新,调节细胞黏附、细胞信号转导等过程[33]。Mucin2、Mucin-5AC 和Mucin-5B 属于凝胶形成型的黏蛋白,具有形成凝胶的能力,结构中包含半胱氨酸富含区域,经二硫键连接形成二聚体,因此其分子量较高不溶于水[34]。人类Mucin2(包含5179aa)有超过100 个不同寡糖修饰后形成的糖蛋白[35],糖分子产生一个高度异构的黏蛋白凝胶层,形成一个天然的防御机制,在肠道内,带电的糖链能有效地控制水分子,形成一个稳定的环境保护半透膜[36]。Mucin-5B 主要在呼吸道黏膜腺体的黏液细胞中表达,在舌下腺高度表达,在颌下腺、子宫内膜、胆囊和胰腺也有表达,有助于唾液和宫颈黏液的润滑和弹性的作用。在胃和呼吸道上皮细胞中Mucin-5AC 通过结合吸入的微生物和微粒,随后被黏液系统清除,来保护黏膜,防止感染和化学损伤[36]。当卵子通过输卵管时即被腺体所分泌的胶状物所包裹,为卵提供丰富的营养物质,此类胶状物多为黏蛋白[37],因林蛙刚出蛰后,温度较低、缺乏食物等为其提供营养物质,输卵管作为重要的生殖器官,分泌大量的胶状物质,达到保护卵子的作用。以上结果提示在哈蟆油中高表达的蛋白质主要参与免疫调节、细胞骨架形成、生殖发育等过程。
哈蟆油本身具有高的膨胀度与其含有大量的、多种类型的凝胶型黏蛋白密切相关。黏蛋白凝胶的程度会受到各种因素的影响,包括黏蛋白自身分子量大小、糖基侧链长短、聚合度和浓度以及外界因素如pH、离子强度的变化等。由于黏蛋白和免疫球蛋白Fc 段连接蛋白都包含大量的糖链结构,对于哈蟆油膨胀的能力具有重要的影响。
4.3 结论
中国林蛙与黑斑蛙的生活习性不同导致哈蟆油与青蛙油中蛋白质含量的差异,本研究通过比较青蛙油与哈蟆油蛋白质组数据库的差异,筛选到差异表达蛋白质55 个,其中上调的蛋白质数为40 个,下调的蛋白质数为15 个。哈蟆油中含量较高的15 差异蛋白质主要参与免疫调节、细胞骨架形成和生殖发育等过程;而青蛙油中含量较高的40 个差异蛋白质主要参与能量代谢、糖代谢、应激反应、生长发育、生物合成、细胞骨架形成、氧化还原和免疫调节等过程;其中AKAP13-like、FcγBP、MUC5AC-like、MUC、MUC5B 等15 个高表达的蛋白质可作为哈蟆油区别于青蛙油的重要蛋白质,同时哈蟆油的特异性膨胀度可能与含量较高的黏蛋白类密切相关,本文的研究结果为进一步研究哈蟆油的功效成分奠定基础。
