植物寄生线虫钙网蛋白的研究进展
2021-08-22魏英罗萌戴良英彭德良刘敬
魏英 罗萌 戴良英 彭德良 刘敬
(1. 湖南农业大学植物保护学院 植物病虫害生物学与防控湖南省重点实验室,长沙 410128;2. 中国农业科学院植物保护研究所 植物病虫害生物学国家重点实验室,北京 100193)
植物寄生线虫是农业生产上的重要病原线虫,每年造成全球超过上千亿美元的损失[1-3]。已知的引起作物严重病害的植物寄生线虫主要有根结线虫(Meloidogyne)、孢囊线虫(Heterodera)、滑刃线虫(Aphelenchoides)、穿孔线虫(Radophoius)、短体线虫(Pratylenchus)、茎线虫(Ditylenchus)、伞滑刃线虫(Bursaphelenchus)等[4]。其中最为严重的是孢囊线虫和根结线虫引起的病害,孢囊线虫大多具有寄主专化性,只寄生在特定或有限的寄主植物上,小麦(Triticum aestivum)、马铃薯(Solanum tuberosum)、大 豆(Glycine max)和 甜 菜(Beta vulgaris)是受危害较为严重的作物。而根结线虫的寄主范围较广,可同时寄生多种植物,包括热带、亚热带和温带地区的多种粮食作物、蔬菜、水果、观赏植物均受到根结线虫侵染和危害。与分布范围相似,根结线虫不同种类的寄主范围差异明显[5-6]。香蕉穿孔线虫(R. similis)广泛分布在热带和亚热带的香蕉产区,是香蕉产业中的主要线虫病害,为害香蕉最明显的症状是引起倒伏。松材线虫(B. xylophilus)对日本、韩国、葡萄牙和加拿大等国的林业造成了十分严重的危害,在我国也有发生 分布[7]。
植物寄生线虫侵染寄主时,通过口针刺穿植物细胞壁,向寄主体内分泌物质来改变寄主细胞的结构和功能,通常将这些分泌物称为效应蛋白[8]。随着植物寄生线虫与寄主互作研究体系的建立,已克隆多个效应蛋白基因,并对其功能进行研究[9-10]。马铃薯白线虫(Globodera pallida)效应蛋白RHA1B是一种具有E3泛素连接酶活性的效应蛋白[11]。甜菜胞囊线虫(H.schachtii)效应蛋白Hs10A07利用寄主修饰系统,磷酸化144号和231号的丝氨酸,在细胞质到细胞核的运输途径中发挥作用[12]。拟禾本科根结线虫(M. graminicola)效应蛋白MgGPP通过寄主的翻译后修饰途径在细胞质内进行C端水解,在进入内质网后进行自身N端糖基化,从而抑制寄主的免疫反应,促进线虫寄生[13]。钙网蛋白(calreticulin,CRT)作为效应蛋白首次在南方根结线虫(Meloidogyne incognita)中被发现,随后其他植物寄生线虫钙网蛋白基因也陆续被克隆报道。但是对于更多的植物寄生线虫钙网蛋白的研究还相对较少,大多数的植物寄生线虫钙网蛋白的功能还不明确。因此,有关植物寄生线虫钙网蛋白的研究可能会成为线虫与植物互作机制研究的一个重要方向。
1 钙网蛋白家族
1974年,钙网蛋白首次在兔骨骼肌肌质网中被分离[14]。钙网蛋白是一类高度保守且功能丰富的蛋白家族,广泛存在于高等生物中,属于内质网(endoplasmic reticulum,ER)滞留蛋白家族,在脊椎动物、无脊椎动物和高等植物中均能分离得到CRT,但目前认为在酵母菌和原核生物中不存在CRT基因[15-20]。它通常包含3个主要内部结构域:由球状的N结构域和两个Ca2+结合结构域组成。两个Ca2+结合结构域分别是高亲和力但低容量的P域,以及低亲和力但高容量的C域,且包含了ER保留信号(HDEL/KDEL)。N末端区(a1-180)氨基酸序列高度保守,P区(a181-280)富含脯氨酸,在CRT发挥分子伴侣功能时与底物结合,C末端区(aa291-400)呈强酸性[21-22](图1)。系统发育研究发现,玉米、大麦、拟南芥等高等植物含有两组不同的CRT:CRT1 / CRT2组和CRT3组[23-24]。这3种亚型的CRT均在拟南芥中有发现,拟南芥CRT亚型AtCRT1A与动物CRT相似,可作为分子伴侣和钙稳态的调节剂[25-27]。前人对人、猴、鼠、鸡、爪蟾、河豚、斑马鱼、文昌鱼、海鞘、海胆和线虫的CRT的进化进行了分析,无脊椎动物及尾索动物的CRT基因结构不稳定;而头索动物文昌鱼和脊椎动物的CRT基因结构则相对稳定,从无脊椎动物到脊椎动物CRT基因结构变得复杂和稳定[20,28-29]。小麦白粉菌的CRT在高温处理下表达量高于未经高温处理的,可能在这种不良条件下对小麦白粉菌起到一种保护作用[30]。植物寄生线虫中,水稻干尖线虫(A.besseyi)的AbCRT-1与松材线虫的BxCRT-1高度相似,香蕉穿孔线虫的Rs-CRT与南方根结线虫的Mi-CRT高度相似,在进化树上处于同一分枝。在NCBI进行对比,象耳豆根结线虫和南方根结线虫的钙网蛋白具有高度相似性,同时与来自禾谷孢囊线虫、短体线虫、松材线虫、香蕉穿孔线虫、水稻干尖线虫和马铃薯腐烂茎线虫的钙网蛋白的相似度也较高(图2)。
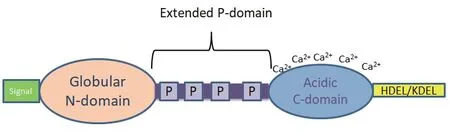
图1 钙网蛋白的结构Fig.1 Structure of calreticulin
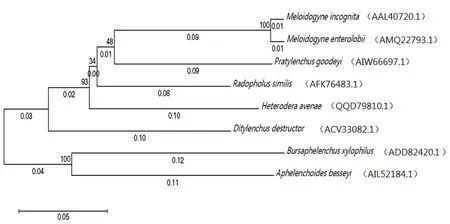
图2 植物寄生线虫钙网蛋白进化关系示意图Fig. 2 Schematic diagram of plant parasitic nematode CRTs evolution relationship
2 植物寄生线虫钙网蛋白的合成及分泌
2.1 钙网蛋白在线虫体内的合成位点
南方根结线虫的钙网蛋白Mi-CRT是第一个从植物寄生线虫鉴定获得的钙网蛋白,它是抑制宿主防御的效应蛋白。原位杂交显示Mi-CRT主要在侵染前二龄(J2)期线虫的亚腹食道腺和寄生时期的背食道腺中表达,表明其主要在食道腺中合成[31]。在之后的研究中也发现效应蛋白HaCRT1,一种禾谷孢囊线虫所分泌的钙网蛋白,也在食道腺中合成[32]。香蕉穿孔线虫钙网蛋白基因Rs-crt是多拷贝基因,在雌性的食道腺和性腺,幼虫的肠和卵中均有表达[33]。类似的研究证实水稻干尖线虫的钙网蛋白AbCRT-1在特异性定位于食道腺和性腺内,且都在食道腺和性腺内表达[34]。除食道腺以外,钙网蛋白在植物寄生线虫的性腺、肠道、卵中合成。这些钙网蛋白在植物线虫不同的部位合成,可能与钙网蛋白在不同线虫中发挥的功能不同相关。
2.2 钙网蛋白在线虫不同龄期的发育表达
固着性内寄生线虫生活史通常分为6个阶段,分别是卵(一龄幼虫)、侵染前二龄(J2)和侵染后二龄(ParJ2)、三龄(J3)、四龄(J4)及成虫(adult)。条件适宜时,卵孵化发育成J2幼虫;J2龄幼虫侵入根组织后,在根内建立永久的取食位点(nematode feeding site,NFS),诱导根部细胞形成合胞体和巨细胞;J2线虫在根内逐渐发育成J3幼虫,J4幼虫,最后再蜕皮形成雌成虫[7]。
明确植物寄生线虫钙网蛋白在不同龄期的表达量有助于了解其在线虫寄生过程中的功能。对水稻干尖线虫AbCRT-1进行qRT-PCR检测,发现AbCRT-1在雌虫中高表达,而在卵,幼虫和雄虫中表达水平低,与AbCRT-1高度同源的BxCRT-1在成虫中表达量高于幼虫,表明AbCRT-1 和BxCRT-1可能与线虫的繁殖有关[34]。而qRT-PCR检测表明,禾谷孢囊线虫的HaCRT1在各龄期均有表达,在侵染后二龄幼虫中最高,说明HaCRT1可能在的早期寄生过程中发挥作用[32]。Mi-CRT在四龄幼虫和雌虫均有表达,原位杂交显示在四龄幼虫背食道腺有很强的信号,因此认为Mi-CRT在南方根结线虫侵入根组织和线虫取食位点的形成过程中起着关键的作用[35-36]。香蕉穿孔线虫的钙网蛋白Rs-crt,在雌虫的食道腺和性腺、雄虫的性腺、幼虫的肠和卵中均表达,并且在雌虫中的表达最高。与雌性相比,雄性、幼虫和卵中Rs-crt表达量较低,雄虫中Rs-crt的表达明显低于幼虫和卵中的,幼虫和卵之间未观察到显著差异[33]。以上表明,不同植物线虫的钙网蛋白不同龄期表达量不同,预示钙网蛋白在植物寄生线虫寄生寄主时发挥着不同的功能。
2.3 植物寄生线虫钙网蛋白在植物细胞中的定位
植物寄生线虫侵染寄主时利用口针向寄主植物分泌效应蛋白来改变植物细胞的结构和功能来促进寄生[8,37]。目前,植物寄生线虫缺乏遗传操作技术,免疫组织定位是研究效应蛋白分泌最直接的手段。在番茄和拟南芥根中对Mi-CRT的分泌进行免疫组织定位,在J3幼虫的口针尖端清楚地检测到Mi-CRT的积累,说明植物寄生线虫在侵染寄主后会将Mi-CRT分泌到植物细胞中。另外,在组成取食部位的巨细胞的细胞壁上也观察到Mi-CRT的积累。同时对南方根结线虫整个寄生过程中的Mi-CRT在植物体内的分泌进行了研究,结果表明J2幼虫会把Mi-CRT分泌到植物组织中,最终分泌到NFS中,表明Mi-CRT对于NFS的形成和维持是必需的[31]。
Mi-CRT在本氏烟(Nicotiana benthamiana)叶片进行亚细胞定位,结果表明无信号肽Mi-CRT定位于烟草叶片细胞的细胞质中,可能由于融合蛋白的被动扩散无信号肽Mi-CRT在细胞核中也检测到信号。有信号肽 Mi-CRT定位于细胞质中。此外,带信号肽 Mi-CRT融合蛋白与ER和高尔基体标记的共定位,表明该蛋白进入了植物的分泌途径。说明Mi-CRT的分泌信号肽在植物细胞中起作用,有信号肽 Mi-CRT通过植物分泌途径被分泌到质体中[31]。而禾谷孢囊线虫的HaCRT1在烟草叶片细胞中的亚细胞定位显示,带有信号肽的和无信号肽的HaCRT1与均与ER标记的信号重叠,表明HaCRT1定位于内质网,信号肽不影响其定位[32]。
3 植物寄生线虫钙网蛋白的功能
3.1 促进线虫寄生
通 过HIGS(host-induced gene silencing)介 导的Mi-CRT沉默显著降低了南方根结线虫对拟南芥侵染,而过表达Mi-CRT的拟南芥对南方根结线虫和及烟草黑胫病菌(Phytophthora parasitica)的易感性增强[31,35]。以上结论表明Mi-CRT在南方根结线虫致病方面发挥重要作用。此外,香蕉穿孔线虫经Rs-crt的dsRNA处理36 h后,线虫的生殖能力和致病性明显降低。HIGS介导的Rs-crt沉默可以有效地传递给后代,而且也显著降低了Rs-crt在线虫体内的表达并提高了转基因番茄对香蕉穿孔线虫的抗性[33]。因此,Rs-crt对拟南芥的繁殖和致病性至关重要。过表达HaCRT1影响拟南芥对丁香假单胞菌(Pseudomonas syringae)的抗性。过表达HaCRT1转基因拟南芥与野生型相比,在接种后第3天,过表达HaCRT1转基因株系的叶片上的病斑更加明显,丁香假单胞菌繁殖量明显增加。水稻干尖线虫钙网蛋白AbCRT-1特异性定位于食道腺和性腺内,并在雌虫中大量表达,瞬时基因沉默后线虫的取食能力降低[34]。此外,在松材线虫中,沉默钙网蛋白基因Bxcrt后,松材线虫的运动异常,并且该线虫的繁殖能力下降,说明BxCRT-1对线虫的繁殖十分重要[38]。以上结论说明在不适宜的环境压力下,植物寄生线虫需要钙网蛋白促进自身寄生与繁殖。
3.2 抑制植物的免疫反应
植物利用两层先天免疫系统抵抗病原体的侵入,第一层是病原相关的分子模式触发的免疫(PAMPtriggered immunity,PTI);第二层是植物进化出的第二道防线,即效应子触发的免疫(effector-triggered immunity,ETI)[39-40]。为了分析Mi-CRT在根结线虫寄生拟南芥中的作用,敲除Mi-CRT基因在线虫中的表达降低了寄生虫侵染拟南芥的能力。相反,过表达Mi-CRT的转基因拟南芥更易受到根结线虫的侵染。在过表达Mi-CRT拟南芥中,防御相关基因PAD4,WRKY33,FRK1和WRKY29的 转 录 水平降低,表明线虫在寄生过程中Mi-CRT抑制寄主PTI[24,31]。同样地,在野生型拟南芥中,flg22强烈诱导了PAD4,WRKY33,FRK1和WRKY29的表达,但是在过表达HaCRT1拟南芥中它们表达量显着低于野生型植物,而且过表达HaCRT1拟南芥flg22诱导产生的ROS也低于野生型。以上结论说明禾谷孢囊线虫HaCRT1抑制了PTI途径。BAX表达可以引起的烟草叶片的细胞死亡,但BAX和HaCRT1的共表达没有诱导细胞死亡,结果表明HaCRT1可以抑制烟草叶片中的BAX诱导的细胞死亡[32]。但是HaCRT1抑制细胞死亡的具体机制还不清楚,是否通过抑制细胞死亡来调控寄主免疫来促进寄生需要进一步研究。
3.3 干扰Ca2+信号传导
钙网蛋白属于内质网滞留蛋白家族,而内质网是Ca2+储存的重要部位。Ca2+通常作为第二信使,在动植物细胞的众多信通路中都起着至关重要的作用[22]。存储在内质网的Ca2+可以与缓冲和释放Ca2+的特定蛋白质结合在一起并控制其局部浓度。在内质网中钙网蛋白具有两个主要功能:伴侣蛋白和调节Ca2+稳态[15,41]。
已经有研究表明植物CRT是主要的Ca2+结合蛋白,并且在细胞内Ca2+稳态和信号传导中发挥作用[42]。蓖麻子CRT含有两个钙结合位点,其CRT也在细胞内Ca2+稳态中起重要作用[43]。过表达ZmCRT的转基因拟南芥对Ca2+消耗低于野生型植物,转基因植物在缺Ca2+的培养基培养后,显示出叶绿素损失延迟。表明ZmCRT的过量表达增加了Ca2+的缓冲能力,并且该Ca2+储备在生物学上可用于植物的生长发育和对胁迫的反应。研究发现植物寄生线虫的钙网蛋白也会影响植物细胞内的Ca2+的浓度变化,在野生型和过表达HaCRT1拟南芥中表达水母发光蛋白(aequorin),利用水母发光蛋白进行 Ca2+浓度标定。用NaCl处理幼苗,植物体内的Ca2+浓度迅速上升形成峰值,发现过表达HaCRT1拟南芥的Ca2+浓度峰值明显高于野生型。尽管研究表明HaCRT1可能通过影响Ca2+信号传导来抑制植物免疫,但是HaCRT1是否具有Ca2+结合的特性,如何调节植物细胞内Ca2+浓度还不清楚。
4 问题与展望
有关钙网蛋白的研究主要在哺乳动物、高等植物、寄生于动物的寄生虫等中研究较多,在植物寄生线虫中的研究还比较少。植物寄生线虫钙网蛋白的研究,目前已知的有水稻干尖线虫钙网蛋白AbCRT-1、松材线虫钙网蛋白BxCRT-1、南方根结线虫钙网蛋白Mi-CRT、禾谷孢囊线虫钙网蛋白HaCRT1、香蕉穿孔线虫的钙网蛋白Rs-CRT、短体线虫钙网蛋白、马铃薯腐烂茎线虫钙网蛋白和象耳豆根结线虫钙网蛋白。AbCRT-1包含多个短的内含子,它参与应激适应,行为模式和生殖。与其高度相似的BxCRT-1只有2个长的内含子,但BxCRT-1是否与松材线虫的寄生有关还不清楚。AbCRT-1和Mi-CRT结构是相似的,虽然RANi介导的AbCRT-1和Mi-CRT都能降低线虫的繁殖能力,但Mi-CRT作为效应蛋白被分泌到寄主体内抑制寄主的防御免疫反应。HaCRT1具有与Mi-CRT相似的功能,可以抑制植物的免疫反应、促进线虫寄生、抑制Bax引起的细胞坏死等。HaCRT1和Mi-CRT在植物内的互作靶标还未找到,所以植物寄生线虫钙网蛋白作为效应蛋白抑制寄主免疫防御反应的分子机制仍不清楚。综述所属钙网蛋白对线虫的寄生、发育及繁殖具有重要作用,并且会降低植物的免疫力。但是植物寄生线虫钙网蛋白的作用机制并不清楚,例如是否具有Ca2+结合特性,作为效应蛋白在寄主体内调控的信号通路。因此,拓展植物寄生线虫钙网蛋白的研究范围和深度,明确钙网蛋白促进线虫寄生的功能和分子机制,对于深入揭示线虫寄生植物的分子机制和绿色防控植物寄生线虫具有重要意义。