基于通径模型对比分析水泊渡水库和倒天河水库后生浮游动物群落结构
2021-08-20孟纯兰李秋华李红梅胡艺李钥
孟纯兰, 李秋华,*, 李红梅, 胡艺, 李钥
基于通径模型对比分析水泊渡水库和倒天河水库后生浮游动物群落结构
孟纯兰1,2, 李秋华1,2,*, 李红梅1,2, 胡艺1,2, 李钥1,2
1. 贵州师范大学贵州省山地环境信息系统和生态环境保护重点实验室, 贵阳 550001 2. 贵州省国际合作研究基地水生态国际联合研究中心, 贵阳 550001
于2015 年的枯水期(1月)和丰水期(8月)进行浮游动物采样,使用通径分析法分析浮游动物群落结构与环境因子的关系。研究结果表明:(1)群落结构特征方面, 轮虫在两座水库的物种种类组成中占比最大, 水泊渡水库和倒天河水库中轮虫分别占各自总物种数的50.00%和72.73%。两个水库的浮游生物的丰度和生物量均存在显著的时空差异, 时间上最高值出现在丰水期, 空间上最高值出现在大坝。相同水库不同水情的优势种有所不同,相同水情期不同水库优势种也不完全一样: 枯水期水泊渡水库优势种为螺形龟甲轮虫()、长额象鼻溞(), 丰水期水泊渡水库优势种为刺盖异尾轮虫()、中剑水蚤()、无节幼体()、长额象鼻溞(); 枯水期倒天河水库优势种为螺形龟甲轮虫()、长额象鼻溞(), 丰水期倒天河水库优势种为螺形龟甲轮虫()、中剑水蚤()、长额象鼻溞()。(2)通径分析表明, 对水泊渡曲腿龟甲轮虫()生物量产生主要影响的关键环境因子为无机氮(DIN)、水柱相对稳定性(RWCS)和电导率(EC); 对水泊渡螺形龟甲轮虫()生物量产生主要影响的环境因子为总氮(TN)、真光层深度(Zue)和电导率(EC); 对水泊渡中剑水蚤()生物量产生主要影响的关键环境因子为总磷(TP)、无机氮(DIN)和深度(H)。对倒天河螺形龟甲轮虫(s)生物量和长额象鼻溞()生物量产生主要影响的环境因子分别为混合层深度(Zmix)和酸碱度(pH)。研究表明, 对不同浮游动物的生物量产生主要的环境因子不完全相同, 不同的水库中对同种浮游动物的生物量产生主要影响的环境因子也不完全相同, 可以根据浮游动物优势种以及现存量(丰度、生物量)大小等进行分析, 判定水库产生主要影响的环境因子, 并且进行控制。
浮游动物; 群落结构; 通径分析; 水泊渡水库; 倒天河水库
0 前言
浮游动物是水域生态系统中重要的次级生产者, 其群落结构的动态变化对着浮游植物的群落结构产生“下行效应”, 对鱼、虾和贝类等生物资源的结构和总量则产生“上行效应”, 对水域生态系统的物质循环和能量流动中起着重要的影响[1]。浮游动物作为反映水环境变化的指示生物, 其种类组成和多样性特征能反映出水体健康程度和状态[2]。因而研究浮游动物群落动态对研究水域健康状况有重要意义[3–4]。然而, 在不同的水域环境中, 导致浮游动物群落结构产生差异的主要因素各不相同。
目前, 浮游动物群落结构与水环境因子间相关关系的分析方法主要为多元线性回归分析、灰色关联分析、主成分分析以及模糊综合评价[5]其中模糊综合评价法和灰色聚类分析等方法本身存在对指标评价的主观性判断和信息丢失的问题[6]而通过主成分分析简化数据信息, 剔除部分不适合因子后再逐步建立多元回归方程的方法所需数据量大, 计算繁琐, 且得到的结果较难一致性地描述因子间的关系和规律[7], 因此寻求一种简便、高效而又科学客观的分析方法显得尤为重要。通径分析是一种可处理较为复杂数据间变量关系的线性回归分析方法, 通径系数可有效分析相关变量之间影响的直接或间接效应, 决定系数可反映出自变量对因变量的作用大小。通径分析比模糊综合分析和灰色关联分析更为全面, 比主成分分析法在数据需求量上更少, 在计算操作上更加便捷, 是一种相对较方便且有效的分析方法, 但该方法多用于分类学, 在浮游动物群落结构研究中的应用还极少见。该研究基于通径模型对贵州两座水库后生浮游动物群落结构信息进行表征, 分析群落结构随季节的变化特征, 探究影响后生浮游动物季节性变化的主要环境因子, 揭示后生浮游动物群落季节演替规律与环境因子的关系, 以期为深入研究水库后生浮游动物的长期变化规律、水库生态系统动态变化机制及生态环境保护措施的制订等提供基础资料和理论依据。
1 材料与方法
1.1 采样点设置与采样时间
水泊渡水库位于贵州省遵义市西南部江河一级支流偏岩河的支流乐民河流域区, 面积共 243 km², 坝高68.8 m,水位高程848 m,库容5510 m³。倒天河水库倒天河发源于贵州省毕节市黄泥大娄山脉南麓, 属乌江水系白甫河(落脚河)支流上游, 位于七星关区大兴桥办事处境内, 距市区约3.5 km, 汇水面积约为120 km², 水库容量1880万m3, 有效库容1800万m3。分别于枯水期(2015 年 1 月) 和丰水期(2015 年 8月) 进行 2 次采样调查分析。根据水泊渡水库水域特点自上游到下游设置为上游(27°53'52″N, 106°77'43″E) 、中游(27°51'75″N, 106°75'65″E) 和大坝 (27°49'41″N, 106°75'16″E) 3个采样点, 由于该水库水位较浅, 故分表层(0.5m) 、中层(10 m) 和底层(20 m) 3 层采样(图1); 根据倒天河水库水域特点自上游到下游设置为上游(27°33'58″N, 105°25' 95″E) 、中游(27°31'82″N, 105°25'86″E) 和大坝 (27° 32'59″N, 105°26'64″E) 3个采样点, 由于该水库水位较浅, 故分表层(0.5m) 、中层(10 m) 和底层(20 m) 3 层采样(图1)。
1.2 样品采集、处理及鉴定方法
定性样品: 用25#浮游生物网于水体表面作“∞”字形缓慢拖网3—5 min, 现场过滤后装入样品瓶中, 并立即加入5%的甲醛溶液保存。定量样品: 用5 L采水器分别在各层采集20 L水样, 每层水样经25#浮游生物网过滤装入样品瓶中, 并立即加入5%甲醛溶液保存。定量和定性样品均在充分摇匀状况下取1 mL至显微镜下镜检. 浮游动物种类按照文献[8–10]进行鉴定,主要鉴定的种类为: 轮虫类、枝角类和桡足类。
1.3 理化因子分析方法
采用YSI便携式多功能水质参数仪测定WT(水温)、pH、ρ(DO)等; 采用塞氏盘现场测定SD(透明度); 参照GB 3838—2002《地表水环境质量标准》分析方法测定ρ(TN)、ρ(TP)、ρ(NO3--N)、ρ(NO2--N)、ρ(NH4+-N)、ρ(PO43--P); 采用反复冻融、丙酮浸提法测定ρ(Chla)[11]。
1.4 数据分析方法
采用优势度值来表示浮游动物优势种(林志, 2018)[12]:

式中:n为第种的个体数;为所有种类总个体数;f为第种在各采样点出现的频率;为优势度值, 优势种为优势度值大于0.02的浮游动物种类.
本文中以透明度的 2.7 倍为真光层深度(Zeu)[13]。已知当采样地点确定以后, 水体中温度改变(≥1℃) 的深度被视作混合层深度(Zmix) , 故可由温度曲线估计。真光层与混合层的比率(Zeu:Zmix) 则成为光可获得性的评判标准[14]水柱相对稳定性(RWCS) 依据 Padisák 等[15]的方法计算:

图1 采样点示意图
Figure 1 Schematic diagram of sampling points

式中,D为底层水体的密度,D为表层水体的密度,4和5分别代表 4 ℃和5 ℃下的水体密度。水体密度由Krambeck 方程计算:
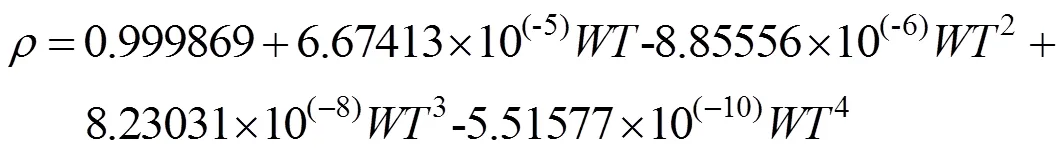
式中, WT 表示水温。在本文中, 0.5 m 为水体表层深度。
通径分析用于分析多个自变量与因变量之间的线性关系, 是回归分析的拓展, 可以处理较为复杂的变量关系。其原理就是将自变量与因变量的单相关系数加以分解, 分解出自变量对因变量的直接影响力和间接影响力。这一分析方法已广泛应用于动植物遗传育种和作物栽培的研究工作中, 也用于其他领域。通径分析的显著性检验包括以下四项: ①回归方程显著性检验: 采用检验法; ②通径系数显著性检验: 采用检验法或检验法; ③通径系数差异显著性检验: 采用检验法或检验法; ④两次通径分析相应通径系数显著性检验: 采用检验法或检验法。一般情况下, 第③种检验和第④种检验在一般的多元线性回归分析中无法实现, 因为不同偏回归系数带有不同量纲, 但是在通径分析中, 这两种检验可以实现。具体通径系数计算方法如下: 根据相关性系数() 矩阵建立正则方程组, 利用高斯消元法得到各环境因子对浮游动物生物量影响的直接通径系数(P) 。根据相关矩阵的组成原理, 获得各环境因子通过其他环境因子对浮游动物生物量产生影响的间接通径系数 (∑rijP) , 依次对各水环境因子对浮游动物生物量的影响展开通径分析, 并计算各因子对浮游动物生物量产生影响的决定系数(d) 以及通过其他因子影响浮游动物生物量的同决定系数(d) 最后通过检验偏回归系数剔除不显著因子建立多元回归方程, 并对多元回归模型进行了假设性检验[16–17]。
“应声虫”现象也被称为“yesman”现象[8],所谓“应声虫”主要是指针对上级的应声。在委托代理模型中,代理人或具有代理人资格的那部分中间阶层,为了得到上一级的认可,不顾实际情况作决策,对委托人不负责任。“地方政府往往会隐蔽行为,在政策的制定、决策的执行、方案的选取等方面不是真正从当地的实际出发,而是看上级政府眼色行事,满足上级主观偏好,表现出一种讨好取巧的代理人机会主义倾向。”[9]
直接通径系数(P) 的计算:
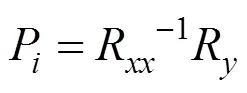
相关系数的组成原理:
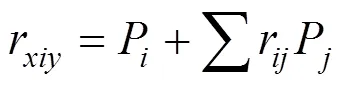
决定系数(d) 的计算:

共同决定系数(d) 的计算:

式中,R-1为水环境因子的相关矩阵的逆矩阵;R为x环境因子对生物量的相关矩阵;为环境因子对环境因子的相关系数。
采用SigmaPlot12.5软件对水库环境因子进行分析; 浮游动物优势种占比、生物量分布图采用Origin 9.0绘制; 采用Arcgis软件对采样图以及后生浮游动物丰度分布进行绘制; 采用SPSS 21软件对群落结构与环境因子的关系进行通径分析。
2 结果与分析
2.1 环境因子
2座水库营养盐有所差别, 但浓度差别不大, 营养盐的变化体现出较强的季节性,具体如下: 两次采样调查期间磷酸根(PO4)、总氮(TN)、叶绿素(Chl.a)、电导率(EC)、酸碱度pH、深度(H)在两座水库之间存在显著性差异(=35,<0.05,=0.73),总磷(TP)、无机氮(DIN)和温度(WT)在两座水库之间无显著性差异(=35,<0.05,=0.82), 真光层深度(Zeu)、真光层与混合层的比率(Zeu:Zmix)和水柱相对稳定性(RWCS) 在两座水库之间无显著性差异(=12,< 0.05,=0.78)。水泊渡水库, 总磷(TP)、温度(WT)、电导率(EC)、酸碱度(pH)、透明度(SD)和水柱相对稳定性(RWCS)在枯水期与丰水期均存在显著性差异(=18,<0.05,=0.78); 总氮(TN)、无机氮(DIN)、真光层深度(Zue)、混合层深度(Zmix)和真光层与混合层的比率(Zeu:Zmix)在枯水期与丰水期均存在无显著性差异(=18,>0.05 ,=0.86)。倒天河水库, 无机氮(DIN)、温度(WT)、混合层深度(Zmix)和真光层与混合层的比率(Zeu:Zmix)和电导率(EC)在枯水期与丰水期均存在显著性差异(=17,<0.05,=0.87); 总磷(TP)、总氮(TN)、透明度(SD)、真光层深度(Zue)、混合层深度(Zmix)和水柱相对稳定性(RWCS)在枯水期与丰水期均存在无显著性差异(=17,>0.05,=0.95)。
2.2 后生浮游动物群落结构
水泊渡水库 3 个采样点 2 次采样调查共鉴定出浮游动物14种, 其中轮虫类7种, 占总数的50.00%; 枝角类3种, 占总数的21.43%; 桡足类4种, 占总数的28.57%。两个水情期种类数中轮虫占比最大, 枝角类和桡足类占比较小。枯水期检出浮游动物7种, 其中轮虫类2种, 占总数的28.57%; 枝角类2种, 占总数的28.57%; 桡足类3种, 占总数的42.86%。丰水期检出浮游动物12种, 其中轮虫类7种, 占总数的58.33%; 枝角类3种, 占总数的25.00%; 桡足类2种, 占总数的16.67%。

图2 水泊渡水库和倒天河水库枯水期和丰水期环境因子对比
Figure 2 comparison of environmental factors between Shuibodu Reservoir and Daotianhe Reservoir in dry and wet periods
倒天河水库 3 个采样点 2 次采样调查共鉴定出浮游动物11种, 其中轮虫类8种, 占总数的72.73%; 枝角类2种, 占总数的18.18%; 桡足类1种, 占总数的9.09%。两个水情期种类数中轮虫占比最大。枯水期检出浮游动物6种, 其中轮虫类4种, 占总数的66.67%; 枝角类1种, 占总数的16.67%; 桡足类1种, 占总数的16.67%。丰水期检出浮游动物10种, 其中轮虫类7种, 占总数的70.00%; 枝角类2种, 占总数的20.00%; 桡足类1种, 占总数的10.00%。

表1 水泊渡水库和倒天河水库后生浮游动物组成对比
2.2.2 优势种
由图3水泊渡水库浮游动物常见优势种曲腿龟甲轮虫()、螺形龟甲轮虫()、刺盖异尾轮虫()、中剑水蚤()、镖水蚤()、无节幼体(s)、长额象鼻溞()、颈沟基合溞(、溞属(sp.), 枯水期主要以螺形龟甲轮虫()、长额象鼻溞()为主, 其丰度分别为28.08、15.50 个·L-1, 其优势种丰度占据整个季节总优势种丰度分别为52.28%、28.85%; 丰水期在各采样点优势种物种数分布较多, 优势种总丰度高于枯水期, 主要以刺盖异尾轮虫(a)、中剑水蚤(、无节幼体()、长额象鼻溞()为主, 其丰度分别为29.25、107.34、36.33、123.33 个·L-1, 其优势种丰度占据整个季节总优势种丰度分别为8.75%、32.10%、10.86%、36.88%。
倒天河水库后生浮游动物常见优势种曲腿龟甲轮虫()、螺形龟甲轮虫()、刺盖异尾轮虫()、中剑水蚤()、无节幼体()、长额象鼻溞(); 枯水期的优势种为螺形龟甲轮虫()、长额象鼻溞(), 其丰度分别为323.35、766.08 个·L-1, 其优势种丰度占据整个季节总优势种丰度分别为29.68%、70.32%; 丰水期在各采样点优势种物种数分布较多, 优势种总丰度高于枯水期, 主要以螺形龟甲轮虫()、中剑水蚤(、长额象鼻溞()为主, 其丰度分别为81.33、50.90 、183.50 个·L-1, 其优势种丰度占据整个季节总优势种丰度分别为21.68%、13.57%、48.93%。
2.2.3 后生浮游动物的丰度和生物量时空分布特征
水泊渡水库浮游动物两个水情期平均丰度为 (67.49±54.31) 个·L-1, 平均生物量为 (1.38± 1.27)mg·L-1。其中, 桡足类平均丰度最高, 为(78.52± 73.10) 个·L-1, 占总平均丰度的 38.78%, 生物量 (2.99± 2.89) mg·L-1最高, 占总平均生物量的72.10%; 枝角类丰度次之, 为(72.63±54.15) 个·L-1, 占总平均丰度的 35.87%, 其生物量(1.15±0.88) mg·L-1最高, 占总平均生物量的 27.67%; 轮虫平均丰度为 (51.31±21.49) 个·L-1, 平均生物量为(0.01±0.01) mg·L-1。调查期间浮游动物丰度变化范围为(53.71—351.20) 个·L-1。根据相关性分析结果, 水泊渡水库后生浮游动物丰度两个水情期之间存在显著差异(=18,<0. 01,=0.10), 其最高值出现在丰水期, 最低值出现在枯水期。生物量两个水情期变化范围为(0.37—7.92) mg·L-1, 相关性分析结果显示, 浮游动物两个水情期生物量也存在显著差异(=18,<0. 01,=0.933) , 其最高值出现在丰水期, 最低值出现在枯水期。水泊渡水库后生浮游动物的丰度和生物量均具有显著的空间差异(=18,<0. 01,=0.96) , 具体表现为大坝(167.10 个·L-1) >上游(146.20 个·L-1) >中游(91.61 个·L-1); 生物量最低值也出现在中游(2.50 mg·L-1) , 最高值出现在大坝 (5.58 mg·L-1) 。整体来说, 丰度与生物量均呈南高北低的变化趋势(图5)。
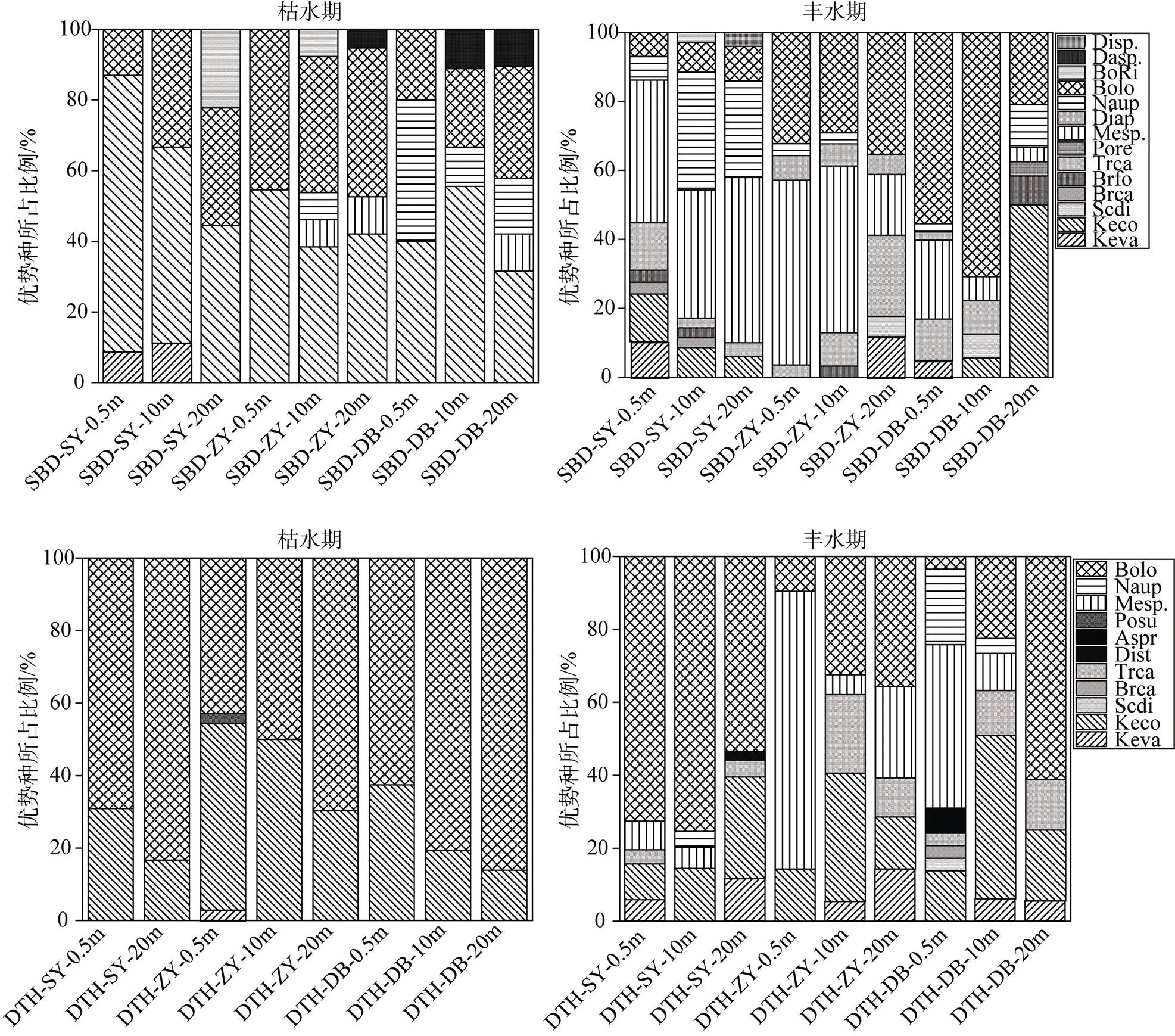
图3 优势种分布图
Figure 3 distribution of dominant species
倒天河水库后生浮游动物两个水情期平均丰度为(247.39±254.69) 个·L-1, 平均生物量为(1.01± 1.25)mg·L-1。其中, 枝角类平均丰度最高, 为(474.79±291.29) 个·L-1, 占总平均丰度的 63.97%, 生物量 (1.68± 1.25) mg·L-1最高, 占总平均生物量的55.51%; 轮虫丰度次之, 为(235.81±102.06) 个·L-1, 占总平均丰度的 31.77%, 其生物量 (0.03±0.02) mg·L-1最低, 占总平均生物量的 0.93%; 桡足类平均丰度为 (31.58±30.33) 个·L-1, 平均生物量为(1.32±1.26) mg·L-1。调查期间后生浮游动物丰度变化范围为(379.15—1105.20) 个·L-1.根据相关性分析结果, 倒天河水库后生浮游动物丰度两个水情期之间存在显著差异(=17,<0. 01,=0.76) , 其最高值出现在丰水期, 最低值出现在枯水期。生物量两个水情期变化范围为(0.50—5.56) mg·L-1, 相关性分析结果显示, 后生浮游动物两个水情期生物量也存在显著差异(=17,<0. 01), 最高值出现在丰水期, 最低值出现在枯水期。倒天河水库后生浮游动物的丰度和生物量均具有显著的空间差异(=17,<0. 01) , 具体表现为大坝(218.95 个·L-1) >上游(186.02 个·L-1) >中游(132.03 个·L-1); 生物量最低值出现在大坝(1.61 mg·L-1) , 最高值出现在上游 (2.51 mg·L-1)。

图4 浮游动物丰度时空分布图
Figure 4 Spatiotemporal distribution of zooplankton abundance
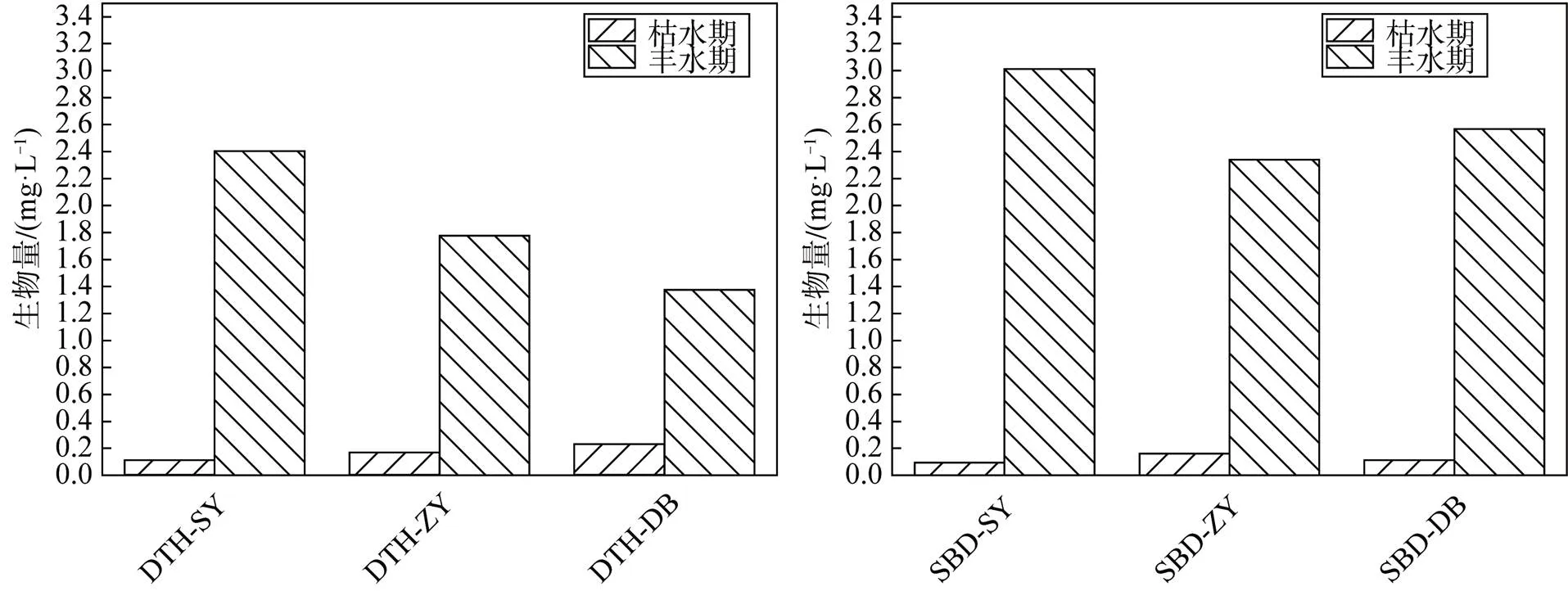
图5 浮游动物生物量时空分布图
Figure 5 Spatial and temporal distribution of zooplankton biomass
2.3 运用通径分析方法探究群落结构与环境因子的关系
对优势种(优势度>0.02)的生物量(分层取平均值)进行选取, 水泊渡水库中选取的优势种为曲腿龟甲轮虫、螺形龟甲轮虫、和中剑水蚤, 倒天河水库中选取的优势种为螺形龟甲轮虫和长额象鼻溞, 即把因变量y设为以上对应物种的生物量。把对应的环境因子TP(mg·L-1)、PO4(mg·L-1)、TN(mg·L-1)、DIN(mg·L-1)、Chl.a(mg·m-3)、WT(℃)、SD(m)、Zue(m)、Zmix(m)、Zue: Zmix、RWCS、EC (µs·cm-1)、pH、H(m)设为自变量1、2、3、4、5、6、7、8、9、10、11、12、13、14。
2.3.1 对进行正态性检验
Kolmogorov-Smirnov Test 和Shapiro-Wilk Test。Kolmogorov-Smirnov Test 检验结果较精确, 适用于大样本的检测, 而 Shapiro-Wilk Test 适用于小样本的检验, 本题属于小样本, 因此对因变量进行正态性检验后利用Shapiro-Wilk Test 的输出结果。 Shapiro-Wilk显著水平 Sig. 均大于0.05, 所以因变量服从正态分布, 即以上优势物种的生物量是正态变量可以进行回归分析。
2.3.2 通径分析
通过逐步回归分析筛选出影响浮游动物生物量变化的关键因子, 计算相关系数, 直接通径系数和间接通径系数。以水泊渡水库两个水情曲腿龟甲轮虫生物量为因变量、关键因子为自变量进行通径分析, 结果见表3, 单项因子及两两因子对生物量的影响程度见决定系数矩阵表4。水库中对曲腿龟甲轮虫生物量有直接正向作用的环境因子为4(DIN)和11(RWCS), 其中11(RWCS)对曲腿龟甲轮虫的直接影响相对较小, 对曲腿龟甲轮虫生物量有直接负向作用的环境因子为12(EC); 在各因子对曲腿龟甲轮虫生物量的间接影响中, 通过11(RWCS)所产生的间接作用均为最大;通过12(EC)产生的间接作用均为正向。其中四关键因子对曲腿龟甲轮虫生物量的直接作用都大于其间接作用; 可见四种环境因子与曲腿龟甲轮虫生物量的变化都存在一定独立的线性关系。决定系数矩阵中对角线以上为单因子对曲腿龟甲轮虫生物量的决定系数, 对角线以下为两两因子对曲腿龟甲轮虫生物量的共同决定系数, 决定系数越大, 影响程度越大。4(DIN)和11(RWCS)的决定系数分别为0.906 和0.020, 其单因子及两因子共同决定系数之和为0.827, 为所有关键因子总决定系数和的 82.69% 。可见4(DIN)和11(RWCS)是与曲腿龟甲轮虫生物量相互影响最主要的两个环境因子。根据两轮通径分析的结果, 利用4(DIN)、11(RWCS)和12(EC)对水泊渡曲腿龟甲轮虫生物量Y建立多元回归方程:

表2 生物量正态性检验
注: *. 这是真实显著水平的下限; Lilliefors 显著水平修正
水泊渡水库曲腿龟甲轮虫生物量=(6.114E-05)4+(-3.164E-07)12+ (1.157E-08)11
以水泊渡水库两个水情螺形龟甲轮虫生物量为因变量、关键因子为自变量进行通径分析, 结果见表4。水库中对曲腿龟甲轮虫生物量有直接正向作用的环境因子为3(TN)、12(EC), 对曲腿龟甲轮虫生物量有直接负向作用的环境因子为8(Zue)、14(H); 在各因子对螺形龟甲轮虫生物量的间接影响中,8(Zue)12(EC)通过3(TN)所产生的间接作用均为负向,14(H)通过3(TN)所产生的间接作用为正向;3(TN)、14(H)通过8(Zue)产生的间接作用均为正向,12(EC)通过8(TN)所产生的间接作用为负向;3(TN)、14(H)通过12(EC)产生的间接作用均为负向,8(Zue)通过12(EC)产生的间接作用为正向;3(TN) 通过14(H)产生的间接作用为负向,8(Zue)、12(EC)通过14(H)产生的间接作用均为正向。其中四关键因子对螺形龟甲轮虫生物量的直接作用都大于其间接作用; 可见四种环境因子与螺形龟甲轮虫生物量的变化都存在一定独立的线性关系。决定系数矩阵中对角线以上为单因子对螺形龟甲轮虫生物量的决定系数, 对角线以下为两两因子对螺形龟甲轮虫生物量的共同决定系数, 决定系数越大, 影响程度越大。3(TN)和12(EC)的决定系数分别为0.30 和-0.03, 其单因子及两因子共同决定系数之和为0.73, 为所有关键因子总决定系数和的73.27%, 可见3(TN)和12(EC)是与螺形龟甲轮虫生物量相互影响最主要的两个环境因子。根据两轮通径分析的结果, 利用3(TN)、8(Zue)、12(EC)和14(H)对水泊渡水库螺形龟甲轮虫生物量建立多元回归方程:

表3 关键环境因子变化对水泊渡水库曲腿龟甲轮虫生物量影响的通径系数及决定系数汇总

表4 关键环境因子变化对水泊渡水库螺形龟甲轮虫生物量影响的通径系数及决定系数汇总
水泊渡水库螺形龟甲轮虫生物量=(3.52E-05)3+ (-3.59E-05)8+(6.38E-07)12+(-1.58E-06)14
以水泊渡水库两个水情中剑水蚤生物量为因变量、关键因子为自变量进行通径分析, 结果见表5。水库中对中剑水蚤生物量有直接正向作用的环境因子为1(TP)和14(H), 对中剑水蚤生物量有直接负向作用的环境因子为4(DIN)和11(RWCS);14(H)通过1(TP)产生的间接作用为正向,4(DIN)和11(RWCS)通过1(TP)所产生的间接作用均为负向;1(TP)通过4(DIN)产生的间接作用为正向,11(RWCS)通过4(DIN)产生的间接作用为负向;1(TP)和4(DIN) 通过14(H)产生的间接作用均为正向,11(RWCS)通过14(H)产生的间接作用为负向;1(TP)和14(H) 通过11(RWCS)产生的间接作用均为正向,4(DIN)通过11(RWCS)产生的间接作用为负向。从总通径系数值的绝对值大小来看, 对水泊渡中剑水蚤生物量影响程度大小排序为14(H)>1(TP)>11(RWCS)>4(DIN), 总通径系数值可以理解为环境因子对水泊渡中剑水蚤生物量大小的影响权重, 由此可以看到,14(H)对水泊渡中剑水蚤生物量大小的影响最大, 而4((DIN)对水泊渡中剑水蚤生物量大小的影响最小。决定系数矩阵中对角线以上为单因子对中剑水蚤生物量的决定系数, 对角线以下为两两因子对中剑水蚤生物量的共同决定系数, 决定系数越大, 影响程度越大。1(TP)和14(H)的决定系数分别为0.742 和1.792 , 其单因子及两因子共同决定系数之和为2.283 , 为所有关键因子总决定系数和的86.08% , 可见1(TP)和14(H)是与中剑水蚤生物量相互影响最主要的两个环境因子。
根据两轮通径分析的结果, 利用1(TP)、4(DIN)、14(H)和11(RWCS)对水泊渡中剑水蚤生物量建立多元回归方程:
水泊渡水库中剑水蚤生物量=0.635+ 12.2281-0.2044+0.00614-0.0211
分别以倒天河水库两个水情螺形龟甲轮虫生物量和长额象鼻溞生物量为因变量、关键因子为自变量进行通径分析, 结果见表6、表7, 通过逐步回归分析筛选出的影响倒天河螺形龟甲生物量和长额象鼻溞生物量变化的关键因子均为一个, 分别为9(Zmix)、13(pH)。
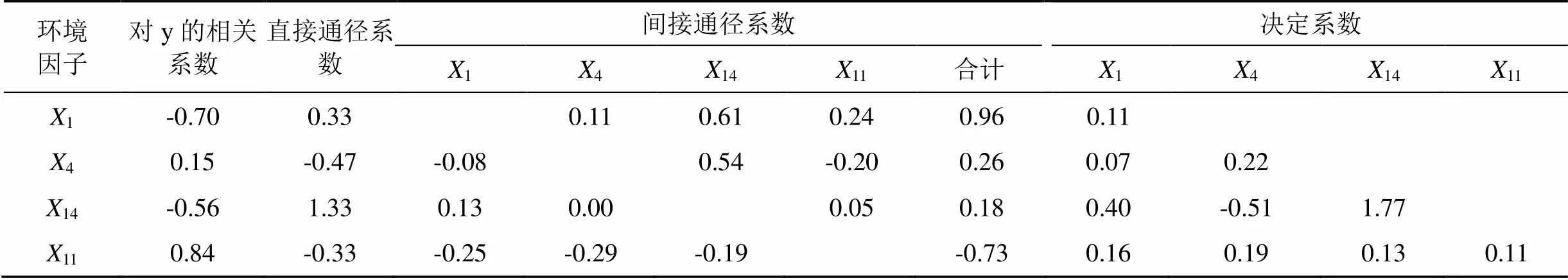
表5 关键环境因子变化对水泊渡水库中剑水蚤生物量影响的通径系数及决定系数汇总
根据通径分析的结果, 分别利用9(Zmix)和13(pH)对倒天河螺形龟甲生物量Y和长额象鼻溞生物量建立多元回归方程:
倒天河螺形龟甲生物量=0.001+(7.07E-05)9
倒天河长额象鼻溞生物量=27.004-3.19713
3 讨论
3.1 水库水环境因子的对比分析
有研究表明: 贵州水库一般比较深, 在不同时期有水体明显分层现象, 在枯水期出现明显的垂直分层现象, 丰水期各采样点垂直断面内温度变化趋势相同; 贵州省水库枯水期透明度明显好于丰水期[18]。对比 2 座水库水环境, 营养盐无显著差异, 流域面积、深度及水动力学参数(WT、Zmix、SD、Zeu、RWCS、光的可获得性)存在显著性差异。已有研究表明湖泊大小会通过影响水体形态和化学性质进而影响到浮游植物功能群的变化从而影响浮游动物群落结构的变化[19]。水泊渡水库和倒天河水库 TN浓度已超出了国际公认富营养化的阈值标准(TN =0.2 mg·L-1)因此氮不再是影响植物生长的限制因子, 进而为浮游动物的生长提供了保障。水泊渡水库光的SD和可获得性及RWCS 均远高于倒天河水库, 故水泊渡水库扰动小。水泊渡水库多样性指数高于倒天河水库, 可知水泊渡水库群落结构复杂, 生态系统内外稳定性较高, 对环境变化适应性较强。

表6 关键环境因子变化对倒天河水库螺形龟甲生物量影响的通径系数

表7 关键环境因子变化对倒天河水库长额象鼻溞生物量影响的通径系数
3.2 水库浮游动物群落结构时空分布特征及与水环境因子的关系
有研究表明: 水体中WT的变化, 能够直接影响轮虫群落的密度, 适宜的水温可以提升轮虫群落的密度, 水温过高或过低, 直接导致轮虫群落密度的减少p H的变化, 即水体中酸碱度发生变化, 对轮虫生活和分布起到一定作用[20]。在浅水湖泊中, 水体中磷含量与水体营养状况密切相关, 是判定水体富营养化程度的一个重要指标; 电导率可以反映水体中离子浓度, 也可以反映出水体的营养状况[21], 二者变化均对水体中浮游植物造成影响[22], 进而导致浮游动物群落结构产生差异[23]。浮游动物种类与丰度的大小与水体酸碱度密切相关,碱性环境有利于浮游植物的光合作用,促使浮游植物大量繁殖,因此碱性环境中浮游植物的生产力较高,为浮游动物生长提供了充足的饵料,促使其丰度增加。螺形龟甲轮虫等对水体中氮磷的相关性极为显著,具有富营养化指示作用,可作为监测水质的指示生物[24]。有研究表明, 大型浮游动物枝角类和桡足类透明度和温度均呈正相关,与pH 值、TP呈负相关[25]。张才学在研究湛江港湾浮游桡足类群落结构的季节变化和影响因素时发现, 桡足类丰度与水温、叶绿素a和浮游植物细胞丰度呈极显著的正相关, 与pH的相关性不明显[26]。研究表明, WT、pH、TP和NO3是影响百花水库后生浮游动物群落结构变化的主要因素[27],轮虫丰度与透明度、总氮、硝态氮呈极显著正相关;枝角类丰度与透明度、硝态氮呈极显著正相关,与温度呈极显著负相关[28]。有研究表明, 大型浮游动物枝角类和桡足类透明度和温度均呈正相关,与pH 值、TP呈负相关[29]。上述研究表明, pH值、电导率、氮磷、电导率等都是影响浮游动物群落结构的重要环境因子, 这与本研究的结果是一致的。上述研究表明, 不同水库湖泊中同种浮游动物受主导影响的环境因子不完全一样, 在本研究中TN、EC、Zue和H是影响水泊渡水库螺形龟甲轮虫生物量的主要环境因子而影响倒天河水库影响螺形龟甲轮虫生物量的主要环境因子是Zmix。造成这一现象的原因可能是在不同的水域环境中水体形态和化学性质不同, 从而使得影响浮游动物群落结构的主导生态因子不同。
[1] 陈雷, 徐兆礼, 姚炜民, 等. 瓯江口春季营养盐、浮游植物和浮游动物的分布[J]. 生态学报,2009, 29(3): 521–527.
[2] PARK K S, SHIN H W. Studies on phyto-and-zooplankton composition and its relation to fish productivity in a west coast fish pond ecosystem[J]. Journal of Environmental Biology, 2007, doi: 10.1016/j.jconhyd.2006.08.012.
[3] TAVEMIN S, MuraMURA G, RossettiROSSETTI G, et al. Factors influencing the seasonal pHenologyphenology and composition of zooplankton communities in mountain temporary pools[J]. International Review of Hydrobiology, 2005, 90(4): 358–375.
[4] EchanizECHANIZ S A, VignattiVIGNATTI A M, De PaggiPAGGI S J, et al. Zooplankton seasonal abundance of south AmericanSaline shallow lakes[J]. International Review of Hydrobiology, 2006, 91(1): 86–100.
[5] 林青, 由文辉, 徐凤洁, 等. 滴水湖浮游动物群落结构及其与环境因子的关系[J]. 生态学报, 2014, 34(23): 6918– 6929.
[6] 邹志红, 孙靖南, 任广平. 模糊评价因子的熵权法赋权及其在水质评价中的应用[J]. 环境科学学报, 2005, 25(4): 552–556.
[7] NOORI R, SABAHI M S, KARBASSI A R, et al. Multivariate statistical analysis of surface water quality based on correlations and variations in the data set[J]. Desalination, 2010, 260(1/3): 129–136.
[8] 王家楫. 中国淡水轮虫志[M]. 北京: 科学出版社, 1961.
[9] 沈嘉瑞. 中国动物志·节肢动物门·甲壳纲·淡水桡足类[M]. 北京: 科学出版社, 1979.
[10] 蒋燮治, 堵南山. 中国动物志·节肢动物门·甲壳纲·淡水枝角类[M]. 北京: 科学出版社, 1979.
[11] 林少君, 贺立静, 黄沛生, 等. 浮游植物中叶绿素 a 提取方法的比较与改进[J].生态科学, 2005, 24(1): 9–11.
[12] 林志, 万阳, 徐梅, 等. 淮南迪沟采煤沉陷区湖泊后生浮游动物群落结构及其影响因子[J]. 湖泊科学, 2018, 30(1): 173–184.
[13] Cole G A. Text book of limnology[M].Llinois: Waveland Pressinc, 1994.
[14] JENSEN J P, JEPPESEN E, OlrikOLRIK K, et al. Impact of nutrients and physical factors on the shift from cyanobacterial to chlorophyte dominance in shallow Danish Lakes[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1994, 51(8): 1692–1699.
[15] PADISAK J, BARBOSA F, KOSCHEL R, et al.Deep layer cyanoprokaryota maxima in temperate and tropical lakes[J]. Advances in Limnology, 2003, 58:175–199..
[16] 杜家菊, 陈志伟. 使用SPSS线性回归实现通径分析的方法[J]. 生物学通报, 2010, 45(2): 4–6.
[17] 刘春雷, 常玉梅, 梁利群, 等. 转大麻哈鱼生长激素基因鲤表型性状与体质量的相关性及通径分析[J]. 应用生态学报, 2011, 22(7): 238–244.
[18] 李秋华. 贵州高原水库富营养化特征及评价[J]. 贵州师范大学学报(自然科学版), 2018, 36(2): 1–8.
[19] GáborGABOR B, BélaBELA T, GáborGABOR V et al. Functional phytoplankton distribution in hypertrophic systems across water body size[J]. Hydrobiologia, 2016, 764(1): 1–10.
[20] 王汨, 杨柏贺, 孟云飞, 等. 北运河水系河流轮虫群落结构与水环境因子的关系[J]. 暨南大学学报(自然科学与医学版), 2017, 38(6): 467–474.
[21] 都雪, 王齐东, 张超文, 等. 洪泽湖轮虫群落结构及其与环境因子的关系[J]. 湖泊科学, 2014, 26(2): 269–276.
[22] 贺筱蓉, 李共国. 杭州西溪湿地首期工程区浮游植物群落结构及与水质关系[J]. 湖泊科学, 2009, 21(6): 795– 800.
[23] 杜彩丽, 杨丽, 赵诣, 等. 淀山湖浮游动物群落时空分布特征及其与环境因子的关系[J]. 环境科学, 2019, 40(10): 4513–4522.
[24] 林海, 王源, 李冰. 北京市妫水河浮游动物群落结构与水质评价[J]. 生态学报, 2019, 39(20): 7583–7591.
[25] 李钥, 李秋华, 陈文生, 等. 贵州三板溪水库后生浮游动物群落结构的动态变化[J]. 湖泊科学, 2016, 28(2): 340–349.
[26] 张才学, 龚玉艳, 王学锋, 等. 湛江港湾浮游桡足类群落结构的季节变化和影响因素[J]. 生态学报, 2016, 31(23): 25–30
[27] 胡艺, 李秋华, 李钥, 等. 基于NMDS和RDA方法分析贵州百花水库后生浮游动物群落结构动态[J]. 环境科学研究, 2019, 32(9): 1510–1518.
[28] 李秋华, 温远志, 杨卫诚, 等. 贵州百花湖麦西河河口后生浮游动物群落结构及与环境因子的关系[J]. 湖泊科学, 2013, 25(4): 531–538.
[29] 汪财生, 李共国. 疏浚后杭州西湖的桡足类[J]. 湖泊科学, 2006, 18(6): 89–94.
Comparsion of community structure of zooplankton in Shuimodu Reservoir and Daotianhe Reservoir based on the path model
MENG Chunlan1,2, LI Qiuhu1,2,*, LI Hongmei1,2, HU Yi1,2, LI Yue1,2
1. Key Laboratory for Information System of Mountainous Area and Protection of Ecological Environment of Guizhou Province, Guizhou Normal University, Guiyang 550001, China 2. Guizhou International Science and Technology Cooperation Research Base Water Ecology International Joint Research Center, Guiyang 550001, China
In order to study the characteristics of the zooplankton community structure in the Shuibodu Reservoir and the Taotianhe Reservoir in Guizhou Plateau, zooplankton were sampled during the dry season (January) and the wet season (August) in 2015 respectively, and the path analysis was used to analyze the relationship between community structure and environmental factors.The results are as follows. (1) As for the community structure, Rotifer accounted for the largest proportion of species in the two reservoirs, with that of the Shuibodu Reservoir and the Taotianhe Reservoir accounting for 50.00% and 72.73% respectively.Thetemporal and spatial differences in the plankton abundance and biomass in the two reservoirs varied considerably, with the peaktemporal value appearing in the wet season, and the highest spatial value indams.The dominant species in different water regimes of the same reservoir were different, while the dominant species in different reservoirs in the same water regime were also different: the dominant species in Shuibodu Reservoir during the dry season wereand, while the dominant species in Shuibodu Reservoir during the wet season were,and;the dominant species in Daotianhe Reservoir during the dry season wereand, while the dominant species in Daotianhe Reservoir during the wet season were,and.(2) Path analysis showed that inorganic nitrogen (DIN), water column relative stability (RWCS) and electrical conductivity (EC) were the key environmental factors that had a major impact on the biomass ofin the Shuibodu Reservoir; total nitrogen (TN), true light depth (Zue) and electrical conductivity (EC) were the main environmental factors affecting the biomass ofin Shuibodu Reservoir; total phosphorus (TP), inorganic nitrogen (DIN) and depth (H) were the key environmental factors influencing the biomass ofin Shuibodu Reservoir.The key environmental factors affecting the biomass ofs andwere the depth of mixed layer (Zmix) and pH (pH)respectively.Studies showed that the main environmental factors that produce the biomass of different zooplankton were different, andthe environmental factors affecting the biomass of the same species of zooplankton in different reservoirs also had differences.Environmental factors affecting the reservoir can be determined and controlled by analyzing the dominant species of zooplankton and the amount, biomass and size of existing ones.
zooplankton; community structure; path analysis; Shuibodu Reservoir;Daotianhe Reservoir
孟纯兰, 李秋华, 李红梅,等. 基于通径模型对比分析水泊渡水库和倒天河水库后生浮游动物群落结构[J]. 生态科学, 2021, 40(4): 1–12.
MENG Chunlan, LI Qiuhu, LI Hongmei, et al. Comparision of community structure of zooplankton in Shuimodu Reservoir and Daotianhe Reservoir based on the path model[J]. Ecological Science, 2021, 40(4): 1–12.
10.14108/j.cnki.1008-8873.2021.04.001
Q939.5
A
1008-8873(2021)04-001-12
2020-07-01;
2020-09-26
国家自然科学基金项目(U1612442); 贵州省科技厅项目(黔科合平台人才[2018]5805和[2020]6009); 贵州省教育厅项目(黔教合KY字[2017]032)
孟纯兰(1995—), 女, 硕士研究生, 主要研究方向为水域生态学, E-mail: 1430315549@qq.com
李秋华, E-mail: qiuhua2002@126.com
