鄱阳湖-珠湖浮游植物群落结构时空变化特征与环境因子的关系
2021-08-20周慜何亮黄建美张萌王启沛万禀颢熊名日刘足根
周慜, 何亮, 黄建美, 张萌,*, 王启沛, 万禀颢, 熊名日, 刘足根
鄱阳湖-珠湖浮游植物群落结构时空变化特征与环境因子的关系
周慜1,2, 何亮3, 黄建美1,2, 张萌1,2,*, 王启沛1,2, 万禀颢1,2, 熊名日1,2, 刘足根1,2
1. 江西省生态环境科学研究与规划院, 南昌 330039 2. 江西省环境保护工程技术研究中心, 南昌 330039 3. 南昌大学资源环境与化学工程学院, 鄱阳湖环境与资源利用教育部重点实验室, 南昌 330031
珠湖是鄱阳湖唯一具有重要饮用水水源供水功能的大型子湖和国家水质良好湖泊, 但长期以来对其浮游植物群落结构及其时空变化情况缺乏了解。为研究浮游植物在珠湖的空间分布特征及其与环境因子的关系, 2018年1月、7月、12月在珠湖设置13个采样点开展浮游植物调查。三期调查共鉴定浮游植物79种, 隶属于7门58属, 绿藻门和蓝藻门种类最丰富, 浮游植物平均生物量为4.80 mg·L-1, 平均细胞丰度为2.56×106个·L-1。珠湖藻类在空间上分布存在差异, 在7月、1月内珠湖团林—四十里街湖区蓝藻较其他湖区优势明显, 在12月团林—四十里街湖区隐藻的优势度明显高于其他湖区, 双港湖区及县水源地湖区的硅藻较其他湖区优势明显。珠湖水质生物学评价结果表明, 珠湖目前处于中污型向重型过渡的状态。RDA分析显示, TDN、TN、CODMn、TP、WT等环境因子解释了物种与环境之间关系的93.96%, TDN是影响藻类生物量的主要环境因子。
珠湖; 鄱阳湖子湖; 浮游植物; 水质评价; 多样性指数; 冗余分析(RDA)
0 前言
浮游植物群落结构、种类组成及季节变化与水质有极为密切的关系, 影响湖泊生态系统的健康与生态平衡的维系, 因此浮游植物的种类组成、数量及多样性也常常作为评估水质和水生态状况的重要指示生物指标[1-4]。在污染较轻的水环境中往往表现出较高的多样性, 且硅藻占优势, 反之, 在受污染较深的水环境中, 蓝藻、绿藻大量繁殖, 成为优势种, 而硅藻不再占优, 在此类浮游植物群落结构发生明显变化过程中, 不同的指示藻类也参与了不同阶段的演替。因此, 研究湖泊浮游植物组成及演替过程有利于对湖泊的水环境状况的认知、健康状况的评价, 对湖泊保护工作起到一定的指引作用。鄱阳湖周边分布着较密集的子湖群, 水域面积大于40 km2的子湖有蚌湖、珠湖、军山湖、大明湖、大湖池等, 这类子湖多由鄱阳湖湖汊围堤形成, 枯平期基本隔断与鄱阳湖的水力联系, 汛期通过闸口对鄱阳湖湖区进行纳洪分流。多属浅平宽广型湖泊, 平均水深3—8 m, 是鄱阳湖湿地生态的重要组成部分, 研究鄱阳湖大型子湖对鄱阳湖流域有着重要意义。近年来鄱阳湖流域水体污染与富营养化风险与压力日益增加, 尤其是环湖区的部分子湖水环境质量不断下降, 氮磷含量增加、富营养化现象发生愈发频繁, 导致水体功能严重受损, 给人类健康与社会可持续发展带来了潜在风险[5-6]。浮游植物是湖泊水生态系统中的初级生产者, 也是水环境监测中的重要指示生物, 在湖泊生态系统的食物链中扮演重要角色, 揭示湖泊生态系统功能演变趋势[7-8]。目前对鄱阳湖的浮游植物演变趋势研究报道较多, 已有研究显示鄱阳湖浮游植物的演变趋势明显, 其中清洁型种类逐渐减少, 喜营养型种类增加, 藻类生物量和丰度呈现增加趋势[9], 但目前单独对大型子湖开展的浮游植物的演变趋势研究相对较少。本文以鄱阳湖唯一具有集中饮用水水源地功能的大型子湖和国家水质良好湖泊—珠湖为研究对象, 结合珠湖同期水生态环境状况数据, 调查分析其浮游植物种类组成、多样性在季节上的变化特征与分布规律, 评价珠湖水环境污染状况, 以期为鄱阳湖重要子湖水生态环境保护和治理提供科学依据。
1 材料与方法
1.1 样品采集
珠湖位于江西省上饶市鄱阳县西部, 鄱阳湖东缘, 处于东经116°33′06″E—116°48′54″E, 北纬29°02′ 16″N—29°15′11″N, 原系鄱阳湖东部一大湖汊, 于20世纪60年代起修筑珠湖联圩18.87 km, 隔断了与鄱阳湖的水力联系, 因此由天然湖汊演变为受人工调控的湖泊。王家渡—瓢里山—渡口村一线以西湖区水域, 称外珠湖, 以东水域, 称内珠湖。内、外珠湖彼此相互连通, 全湖呈枝汊形。湖泊水域总面积80 km2, 平均水深5.72 m, 蓄水量4.62亿m3。流域范围包括鄱阳县高家岭镇、双港镇、珠湖乡、四十里街镇、团林乡、白沙洲乡6个乡镇, 流域面积373.81 km2。珠湖多年最高水位多出现在7至8月, 多年最低水位出现在12月或1月份。水位年变幅, 一般为6—8 m。修筑珠湖联圩后, 提高了珠湖的饮用、调蓄、灌溉等综合利用效益, 在珠湖启用为集中式饮用水水源地以前, 珠湖水产养殖业发展较集中, 主要养殖草、青、鲢、鲤、银鱼、大闸蟹等水产品种。
2018年1月、7月、12月对珠湖开展湖泊浮游植物调查, 在内珠湖、外珠湖、通道区共布设样点13个, 以反映珠湖不同功能湖区浮游植物状况。其中, 外珠湖DS01为鄱阳县饮用水水源取水口, 外珠湖DS02为饮用水水源二级保护区, DS03点位属外珠湖白沙洲湖区, DS05点位属外珠湖大塘村湖区, DS08点位属外珠湖闸口区, 通常情况下湖水由此单向注入鄱阳湖, 在特大洪水期, 鄱阳湖圩堤防汛压力时, 闸口区为珠湖蓄洪区主要入口, SW13点位属外珠湖双港湖区, DS04点位属外珠湖与内珠湖交换通道区域, DS6点位属内珠湖腰里湖区, DS12点位属内珠湖团林湖区、SW11属于四十里街湖区、DS07、SW11均属于内珠湖湖区。2018年1月、7月、12月波阳站的月累计降水量分别为107.3mm、133mm及163mm。湖泊采样点位示意图如下图1所示。
用25号浮游生物网在采样点的水面和水面下0.5 m处以20—30 cm·s-1的速度作“∞”形循回缓慢拖动约1—3 min, 将采得的水样倾入标本瓶中, 加入鲁哥氏液固定。样品预先经沉淀、浓缩至约30 mL, 每个样品取5滴水样做成5张临时装片, 采用全水量观察法, 用显微镜观察, 参照《中国淡水藻类-系统、分类及生态》[10]采用的分类系统进行种类鉴定。
1.2 样品处理
定量样品用2500 mL有机玻璃采水器取上、中、下层水样, 混合后取1000 mL用鲁哥氏液固定, 室内沉淀36小时, 静置后用虹吸管缓慢在水面以下吸去多余水样, 浓缩至30 mL, 摇匀后吸取0.1 mL样品置于浮游植物计数框内, 在显微镜下按视野法计数, 数量特别少时全片计数, 每个样品计数2次, 取其平均值, 每次计数结果与平均值之差应在15%以内, 否则增加计数次数。每升水样中浮游植物数量的计算公式:

式中:为一升水样中浮游植物的数量(ind.·L-1);为计数框的面积(mm2);c为计数面积(mm2);w为一升水样经浓缩后的体积(mL);为计数框的容积(mL);为计数所得个数(ind.)。
选择10-30个浮游植物个体测定体积平均值, 用体积估算法计算浮游植物生物量(), 公式为:
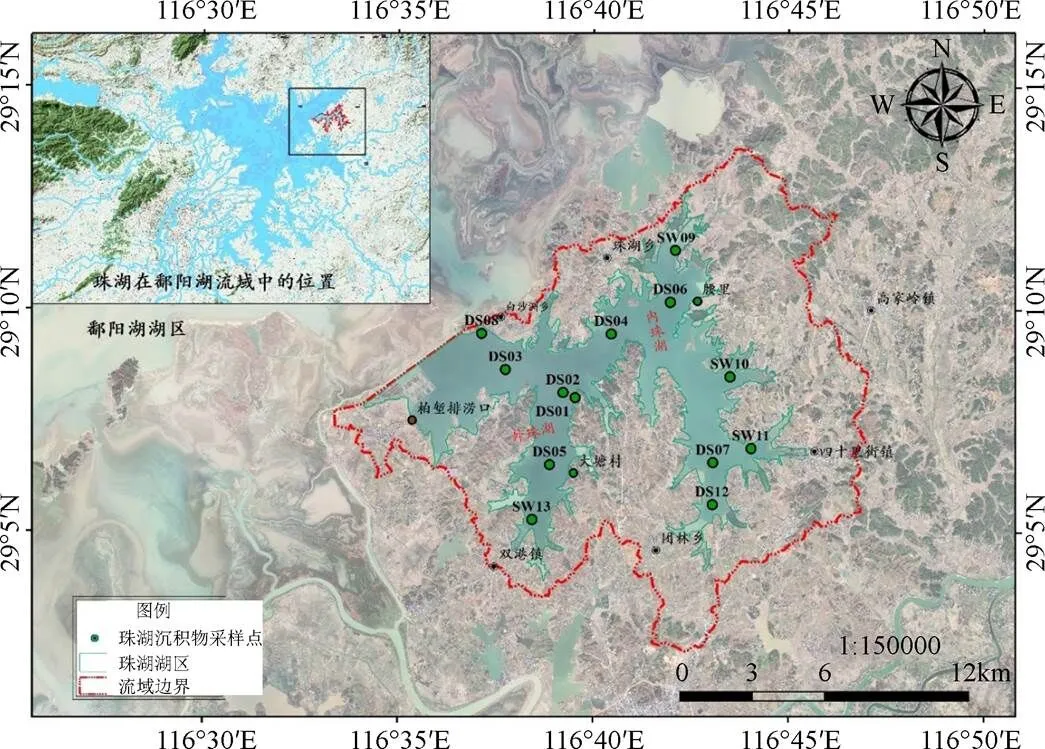
图1 江西鄱阳湖-珠湖采样点位示意图
Figure 1 Location of sampling sites in Zhuhu Lake of Poyang Lake, Jiangxi
=··10-9
利用Shannon-Wiener多样性指数(’)[11,12]、Margalef丰富度指数()[13,14]与Pielou均匀度指数()[15]对珠湖的水质状况进行评价, 3个指数的评价标准见表1。
1.3 理化参数测定
采用YSI-556型快速多参数水质分析仪测定水温(WT)、pH值、溶解氧(DO)、电导率(EC), 采用哈希2100Q型便携式浊度分析仪测定浊度(Turb)。水体化学指标总磷(TP)、可溶性总磷(TDP)采用钼酸铵分光光度法测定, 总氮(TN)、可溶性总氮(TDN)采用碱性过硫酸钾消解紫外分光光度法测定, 氨氮(NH4+-N)采用纳氏试剂分光光度法测定, 高锰酸盐指数(CODMn)采用高锰酸钾法测定, 硝态氮(NO3--N)采用离子色谱法测定, 正磷酸盐(PO43--P)采用钼锑比色法测定, 叶绿素a(Chl a)、悬浮物(SS)浓度等测定参照《湖泊富营养化调查规范》[16], 用塞氏盘法现场测定透明度(SD)。

表1 Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(M)与Pielou均匀度指数(J)的评价标准
1.4 数理统计分析
采用ArcGIS和EXCEL对珠湖藻类群落结构以及水环境因子进行作图, 利用SPSS对不同月份珠湖藻类群落结构通过检验进行单因素方差分析。显著性水平设置为0.05。
选取本次调查的15种优势浮游植物物种, 小环藻、颗粒直链藻极狭变种、针杆藻、浮鞘丝藻、隐球藻、尖头藻、伪鱼腥藻、微囊藻、卷曲鱼腥藻、丛球韦斯(丝)藻、顶锥十字藻、华丽四星藻、锥囊藻、卵形隐藻、尖尾蓝隐藻与16个环境变量(WT、SD、pH、DO、TN、TDN、TP、TDP、CODMn、Chl_a、PO43--P、TDN、NO3--N、EC、Turb和SS)进行排序分析。首先运用Canoco 5.0软件对浮游植物生物量数据先进行DCA(detrended corres pondence analysis, 除趋势对应分析), 计算结果发现物种的单峰响应值SD(梯度)最大值小于3, 因此藻类与水环境因子关联的分析方法选择冗余分析(Redundancy Analysis, RDA)。利用方差膨胀因子分析, 对所有纳入RDA分析的环境因子进行共线性分析, 并删除掉共线性的环境因子pH、Chl a。对剩下的主要环境因子再采用蒙特卡洛置换检验(Monte Carlo permutation test)进行筛选, 选出具有显著性影响的因子。经过蒙特卡洛置换检验后, 采用RDA进一步对本次调查的藻类生物量与环境因子相关性进行研究。为了减少藻类数据的偏锋影响, 对水体理化指标数据除pH外全部进行lg(+1)转换。
2 结果与分析
2.1 浮游植物种类组成及优势种
此次调查三个水情期共采集到浮游植物79种, 隶属于7门58属。其中绿藻门24属41种, 蓝藻门12属14种, 硅藻门11属13种, 甲藻门4属4种, 裸藻门4属4 种, 隐藻门2属3种, 金藻门1属1种。硅藻门生物量占所有门类总生物量的31.75%, 蓝藻门生物量占总生物量的23.89%, 绿藻门生物量占总生物量的17.30%。此次调查中硅藻门、绿藻门、蓝藻门生物量共占总生物量的72.93%, 各水情浮游植物物种以硅藻、蓝藻、绿藻为主, 珠湖属“硅藻—蓝藻”型湖泊。调查结果表明, 珠湖的浮游植物生物量为0.29—8.55 mg·L-1, 平均值为4.80 mg·L-1, 细胞丰度为33.1×104—5327.5×104个·L-1, 平均值为255.5×104个·L-1, 与江西省其他水质良好湖泊相比较, 柘林湖湖区夏季丰度为9960×104个·L-1 [17], 仙女湖湖区夏季为2.19×109个·L-1[18], 而珠湖夏季最高丰度为5327.5×104个·L-1, 略优于柘林湖, 各门浮游植物的生物量和密度见表2。
根据各月份浮游植物出现的频率和丰度, 按照优势度大于0.02来确定优势种, 2018年1月珠湖藻类的优势种有2种, 分别是硅藻门的小环藻(sp.), 其相对生物量为12.8%; 蓝藻门的浮鞘丝藻(sp.), 其相对生物量为9.2%。2018年7月珠湖藻类的优势种有7种, 分别是蓝藻门的浮鞘丝藻(sp.)、隐球藻(sp.)、尖头藻(sp.)、伪鱼腥藻(sp.)、微囊藻(sp.)、卷曲鱼腥藻(), 其相对生物量分别为15.7%、0.2%、3.6%、4.3%、1.2%、1.9%; 硅藻门的颗粒直链藻极狭变种(var.angustissima f.spiralis), 其相对生物量为2.9%。2018年12月珠湖藻类的优势种有10种, 分别是硅藻门的小环藻(sp.)、针杆藻(sp.)、颗粒直链藻极狭变种(f. spiralis), 其相对生物量分别为16.0%、4.2%、1.7%; 绿藻门的丛球韦斯(丝)藻()、顶锥十字藻()、华丽四星藻(), 其相对生物量分别为0.3%、1.1%、0.9%; 金藻门的锥囊藻(sp.), 其相对生物量为9.9%; 蓝藻门的微囊藻(sp.), 其相对生物量为1.6%; 隐藻门的卵形隐藻()和尖尾蓝隐藻(), 其相对生物量为16.6%、1.6%。

表2 珠湖浮游植物生物量和细胞丰度
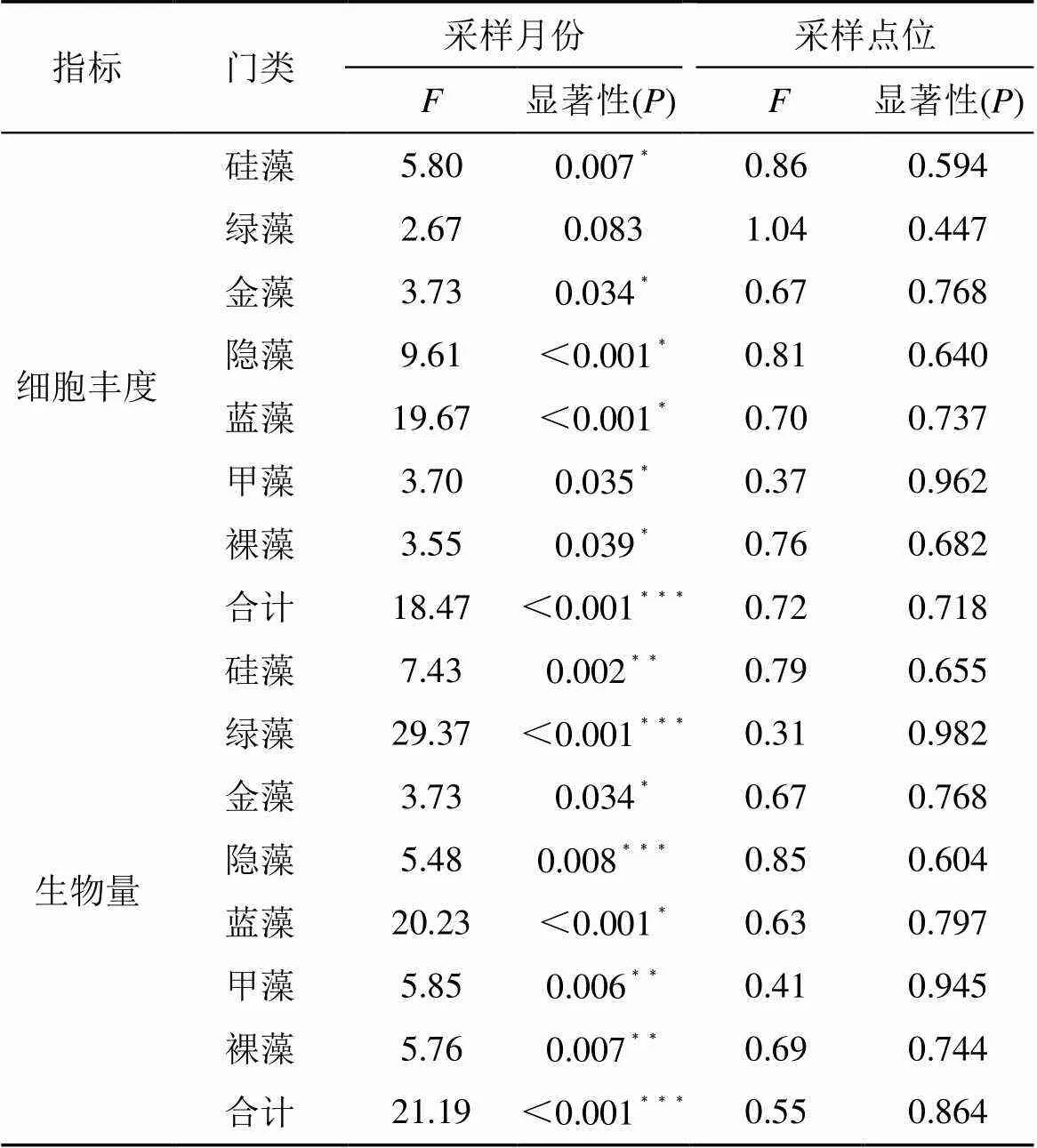
表3 不同因素对珠湖藻类群落结构影响的单因素方差分析
注:<0.05表示具有显著性差异。其中, “﹡”表示<0.05, “﹡﹡”表示<0.01, “﹡﹡﹡”表示<0.001。

表4 珠湖浮游植物优势种和优势度指数
2.2 浮游植物群落的季节变化
2018年7月, 浮游植物生物量最大, 平均生物量高达到3.43 mg·L-1, 而2018年12月的生物量为全年最低, 平均生物量仅为0.55 mg·L-1。珠湖浮游植物不仅总体生物量随季节有变化, 各门藻类的生物量也随季节产生差异, 不同月份间珠湖藻类群落结构差异显著(见表3)。本次调查结果表明, 2018年12月、1月硅藻门生物量最大, 约占38.5%、36.6%, 甲藻门次之, 约占16.8%、20.4%, 硅藻门明显高于其他门类。而在2018年7月蓝藻为优势门类, 约占29.5%, 其次为硅藻门, 约占29.3%, 再次之为绿藻, 约占19.4%。
浮游植物细胞丰度变化与生物量变化略有不同, 2018年12月份的细胞丰度主要由绿藻、蓝藻、硅藻贡献, 分别占39.2%、24.1%、22.9%, 在2018年1月份的细胞丰度主要由蓝藻、绿藻贡献, 分别占74.5%、13.2%, 2018年7月份的细胞丰度主要由蓝藻贡献, 高达92.2%。从细胞丰度所占比例来看, 不同季度下, 蓝藻占优势均较明显, 占比为24.1%—92.2%, 而绿藻在12月份占比也较高, 当月平均占比为39.2%。
2.3 浮游植物的空间分布特征
珠湖不同月份间的藻类群落结构差异显著, 但不同采样点位间藻类群落结构差异不显著(见表3), 在空间上内珠湖的藻类生物量、细胞丰度最高, 外珠湖次之, 通道区域最小。在2018年1月份, 浮游植物的生物量在内珠湖SW11四十里街湖区最高, 生物量为2.18 mg·L-1, 其次为外珠湖DS03白沙洲湖区, 生物量为1.23 mg·L-1, 外珠湖DS01湖区也相对较高, 生物量为1.04 mg·L-1, 而在外珠湖SW13双港湖区、内珠湖DS12团林湖区相对较低, 生物量分别为0.19 mg·L-1、0.31 mg·L-1。从群落结构来看, 除内珠湖SW11四十里街湖区外, 其余各采样点均为硅藻占优势, 内珠湖SW11四十里街湖区蓝藻生物量占优势, 金藻其次, 甲藻再次之, 其余藻类生物量较低。从细胞丰度来看, 内珠湖各采样点, 均由蓝藻贡献占优势, 其次为绿藻或硅藻, 双港湖区细胞丰度贡献以硅藻或绿藻占优, 其余藻类丰度较低。

图2 珠湖浮游植物各门类平均生物量的季节变化
Figure 2 Seasonal variations of biomass in different phyla of phytoplankton in Zhuhu Lake

图3 珠湖浮游植物各门类丰度的季节变化
Figure 3 Seasonal variations of abundance in different phyla of phytoplankton in Zhuhu Lake
在2018年7月份, 浮游植物的生物量在内珠湖SW11四十里街湖区最高, 生物量为8.55 mg·L-1, 其次为内珠湖DS12团林湖区, 生物量为5.21 mg·L-1, 内珠湖SW10湖区、DS07湖区也相对较高, 生物量分别为5.19 mg·L-1、4.78 mg·L-1, 而在外珠湖各湖区生物量要明显低于内珠湖湖区, 外珠湖湖区生物量为0.93 mg·L-1—3.34 mg·L-1。从群落结构来看, SW11四十里街湖区、DS12团林湖区、DS07湖区均由硅藻生物量占优势, 蓝藻次之, 内珠湖SW09湖区蓝藻生物量占优势, 裸藻次之; 外珠湖SW13双港湖区、外珠湖DS08湖区、DS02湖区均为蓝藻占优势, 硅藻或绿藻次之, 外珠湖DS03、通道区DS04均为绿藻占优; 外珠湖DS01湖区生物量硅藻明显占优势, 绿藻次之, 其余藻类生物量较低。从细胞丰度来看, 7月份各点位均由蓝藻细胞丰度贡献占绝对优势, 占比88.0%—96.6%。

图5 珠湖浮游植物生物量的空间分布图
Figure 5 Spatial distribution of phytoplankton biomass in Zhuhu Lake
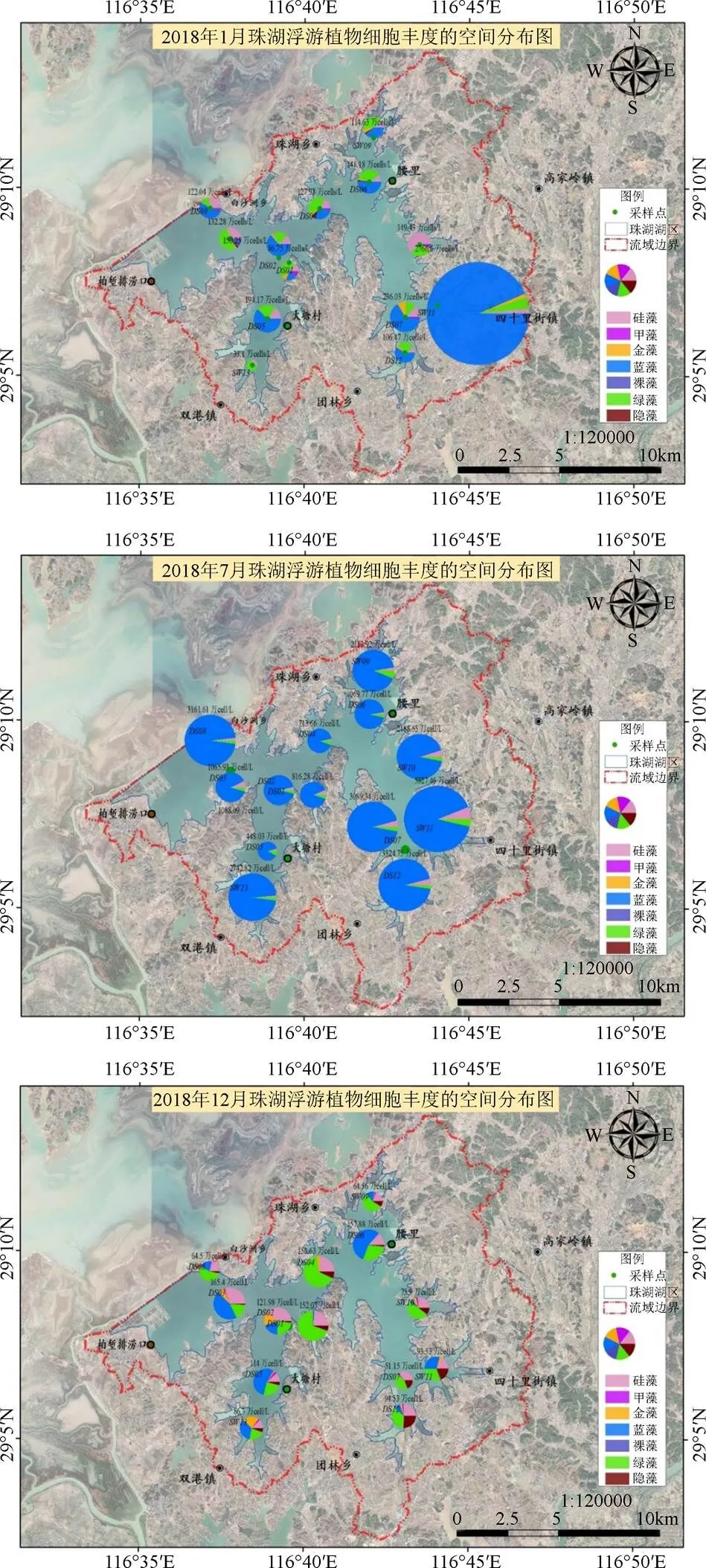
图6 珠湖浮游植物丰度的空间分布图
Figure 6 Spatial distribution of phytoplankton abundance in Zhuhu Lake
在2018年12月份, 浮游植物的生物量在外珠湖DS01最高, 生物量为1.19 mg·L-1, 其次为外珠湖DS03, 生物量为0.69 mg·L-1, 外珠湖DS02湖区也相对较高, 生物量为0.62 mg·L-1。从群落结构来看, 外珠湖DS01、DS02、DS03均由硅藻占绝对优势, 外珠湖SW13双港湖区由金藻占优势, 而内珠湖DS12、SW11、DS07均由隐藻占优。从细胞丰度来看, 外珠湖DS03白沙洲湖区、DS05大塘湖区、SW13双港湖区由蓝藻细胞丰度贡献占优, 外珠湖DS01、通道区DS04、内珠湖团林—四十里街湖区(DS12、SW11、DS07)均由绿藻细胞丰度贡献占优。
2.4 珠湖水质生物学评价
采用浮游植物群落的Shannon-Wiener多样性指数、Pielou均匀度指数以及Margalef丰富度指数对珠湖进行水质生物评价, 结果显示, 珠湖目前处于中污型向重型过渡的状态, 但是不同季节, 不同点位仍有较大差异。
其中Shannon-Wiener多样性指数全年大多介于1-3之间, 1月、7月、12月与全年均值分别为2.03、1.73、1.72与1.83, 1月属于α-中污型水质, 7月和12月属于β-中污型水质, 全年珠湖属于β-中污型水质。说明1月份物种群落结构相对较复杂, 对环境压力的承受能力较强, 群落结构相对较稳定。Shannon-Wiener多样性指数最小值出现在12月内珠湖腰里湖区DS06, 最高值出现在1月份的通道区DS04,′指数在珠湖整体上呈现出: 闸口区<内珠湖<外珠湖<通道区。
Margalef丰富度指数差异较大, 在0.57-2.06范围间波动, 平均值为1.26, 大多介于1—2之间, 占采样总数的82.1%, 小部分介于2—3之间, 占采样总数的2.6%, 部分介于0—1之间, 占采样总数的15.4%, 表明珠湖存在一定的从中度污染型至重污染型过度的风险。Margalef丰富度指数最小值出现在1月份闸口区DS08, 最高值出现在7月份四十里街湖区SW11。指数在珠湖整体上呈现出: 闸口区<内珠湖<通道区<外珠湖。
而Pielou均匀度指数在0.31—0.96范围间波动, 平均值为0.63, 大多介于0.5—0.8之间, 占采样总数的53.8%, 部分介于0.3—0.5之间, 占采样总数的30.8%, 少部分高于0.8, 占采样总数的15.4%, 说明珠湖存在一定的寡污型至β-中污型过度的风险。Pielou均匀度指数最小值出现在7月份外珠湖双港湖区SW13, 最高值出现在1月份外珠湖DS03。指数在珠湖整体上呈现出: 内珠湖<闸口区<外珠湖<通道区。
2.5 环境因子对浮游植物的影响
经蒙特卡洛置换检验对主要环境因子进行筛选后, 保留<0.05的理化因子, 经检验TDN(= 0.002)、TN(=0.020)、CODMn(=0.002)、TP(= 0.002)、WT(=0.002)、TD(=0.004)、NO3--N(= 0.004)、TDP(=0.002)、NH4+-N(=0.004)、DO(= 0.004)、SS(=0.014)与Turb(=0.006)共12个环境因子被保留, 保留的环境因子与本次调查的浮游植物群落时空动态变化关系密切。浮游植物与RDA的分析结果显示, 第1排序轴和第2排序轴特征值分别是0.5161和0.0511, 两轴累计变异百分率解释了物种数据的66.7%, 解释了物种与环境之间关系的93.96%, 表明15种浮游植物与12个环境因子之间存在显著相关性, 其中最重要的因子是TDN值, 解释了总变异率的43.8%, 其后四位分别是TN (43.2%)、CODMn(23.3%)、TP(20.5%)、WT(20.4%)。环境因子对浮游植物生物量解释了66.7%, 表明还有其他的环境因子对浮游植物生物量变化产生重要影响, 浮游植物不仅受到氮、磷、碳源等影响, 还受到珠湖流域的土地利用性质、渔业生产、鱼类群落组成、水生植物和水文等因素影响。

图7 珠湖浮游植物群落Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(M)与Pielou均匀度指数(J)分布
Figure 7 Distribution of Shannon-Wiener index (′) Mangalef index ()and Pielou index () of phytoplankton communities in Zhuhu Lake
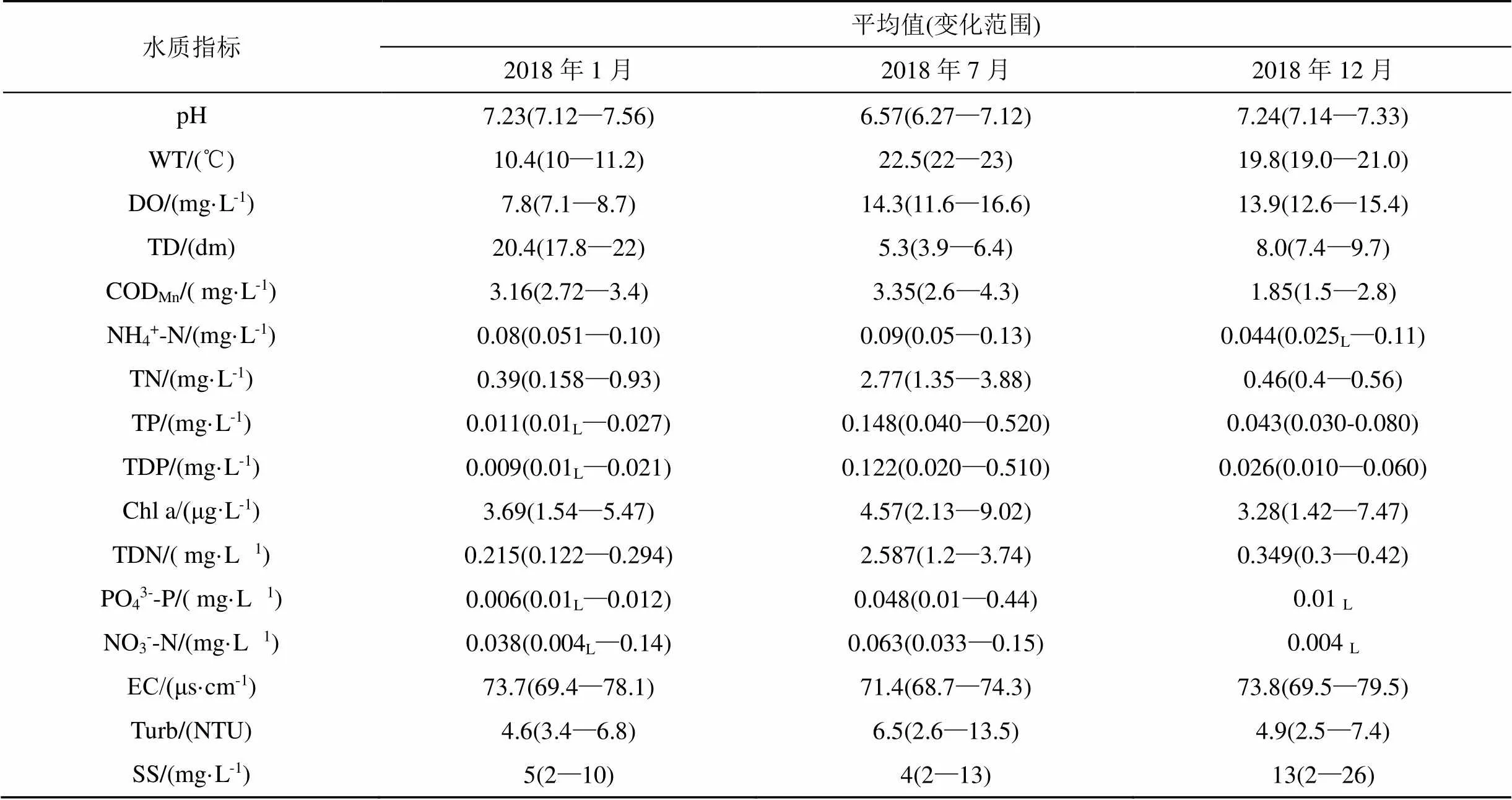
表5 珠湖水体主要环境因子
注: 数字后加“L”表示小于检测方法检出限, 在计算平均值浓度时取二分之一检出限值代入计算。

表6 浮游植物与环境因子的相关关系

注: S1为小环藻, S2为颗粒直链藻极狭变种, S3为针杆藻, S4为浮鞘丝藻, S5为隐球藻, S6为尖头藻, S7为伪鱼腥藻, S8为微囊藻, S9为卷曲鱼腥藻, S10为丛球韦斯(丝)藻, S11为顶锥十字藻, S12为华丽四星藻, S13为锥囊藻, S14为卵形隐藻, S15为尖尾蓝隐藻。
Figure 8 RDA of representative functional groups and environmental variables in Zhuhu Lake
3 讨论
3.1 浮游植物的分布特征
夏季珠湖蓝藻的生物量较1月份出现一定程度上升, 平均生物量占比上升了26.9%, 硅藻平均生物量占比下降了7.3%, 夏季珠湖蓝藻的优势得到进一步巩固。需特别注意的是, 在取水口DS01处7月份蓝藻的平均生物量占比较1月份提高了6.1%, 但微囊藻在蓝藻中生物量占比提高了54.5%, 其可能产生微囊藻毒素, 微囊藻毒素是一种单环七肽化合物, 对水生生物和饮用水水源安全带来潜在的隐患。冬季珠湖蓝藻的生物量较夏季迅速下降, 平均生物量占比下降了68.1%, 硅藻又上升为主要的优势种, 平均生物量占比上升了18.9%。当夏季水温升高, 同时伴有喜温藻类大量繁殖, 表现为蓝藻门的浮鞘丝藻、隐球藻、尖头藻、伪鱼腥藻、微囊藻等喜高温的种类大量出现, 成为优势类群。1月、12月珠湖水温相对较低, 光照强度较弱, 喜低温的硅藻门种类小环藻、针杆藻、颗粒直链藻极狭变种等成为冬季的主要优势类群。12月和1月珠湖均为硅藻占优势, 硅藻比较适合在低温的环境下生长繁殖, 到7月, 转变为蓝藻占优势, 生物量超过硅藻。在7月, 绿藻也出现较明显的增加, 生物量仅次于蓝藻和硅藻, 这也表明夏季水温最高、光照最强, 蓝藻、绿藻具有竞争优势, 生物量达到峰值, 浮游植物是通过叶绿素生产力和细胞分裂速度的改变来适应温度的变化, 温度改变的范围有利于浮游植物生长, 细胞分裂速度也增加。此外, 珠湖较普遍的水产养殖活动等带来的外源污染物输入, 加剧了水体污染, 也加速了富营养化进程。
珠湖存在加剧转向“硅藻—蓝藻”型湖泊的风险, 在7月、1月内珠湖团林—四十里街湖区蓝藻的优势明显高于其他湖区, 在12月团林—四十里街湖区隐藻的优势明显高于其他湖区, 双港湖区及DS01湖区的硅藻的优势明显于其他湖区。内珠湖团林—四十里街湖区为珠湖入湖河流的主要汇入区, 临近团林、四十里街集镇, 地表径流带来的外源性营养面广, 造成隐藻等喜营养型的藻类增加, 硅藻中清洁型的藻类减少, 导致了该湖区与其他湖区在浮游植物种类和生物量上的空间差异。
3.2 珠湖水质多样性指数评价
采用Shannon-Wiener 多样性指数′、Margalef 物种丰富度指数与Pielou 均匀度指数的水质生物学评价结果往往有一定的差异[19], 本次珠湖多样性指数的水质评价结果也存在一定差异, 但本次评价结果整体上呈现出珠湖目前处于中污型向重型过渡的状态, 且外珠湖和通道区水质优于内珠湖和闸口区水质。珠湖为人工调控的围堰湖泊, 呈“π”型, 作为鄱阳湖东岸重要的蓄洪区, 珠湖大汛期开闸蓄洪, 平枯期闭闸蓄水, 以满足环湖的农业与部分生活需水, 正常年份全年较长时间闭闸, 珠湖与鄱阳湖水力联系少, 其水体交换能力明显弱于鄱阳湖的天然湖叉。在珠湖内部, 内外珠湖之间通过狭长的通道区联系, 内珠湖的水系交换能力较外珠湖、通道区差, 内珠湖与闸口区系传统的水产养殖区域, 内珠湖东南侧临近团林、四十里街2个集镇, 系集镇生活污水主要受纳区域, 内珠湖处于不利的水利交换条件, 面临上游来水的富营养化压力, 以及水产养殖外源污染, 致使内珠湖的水质较差于其他湖区。
3.3 浮游植物与环境因子关系
从RAD排序图可看出, 2018年7月采样点主要分布在第一主轴的右侧, S2、S3、S4、S5、S6、S7、S8、S9、S11对7月采样点贡献较大, 而2018年1月、12月采样点主要分布在第一主轴的左侧, S1、S10、S12、S13、S14、S15对1月、12月采样点贡献较大, 相同采样点在不同季节有明显的区别, 浮游植物群落结构、生物量体现了较大的差别。在7月份珠湖, 隐球藻、伪鱼腥藻、微囊藻、卷曲鱼腥生物量与TND、TN、TP、TDP、N03--N、NH4+-N、CODMn、Turb、DO、WT箭头方向一致, 与TD、SS箭头方向相反, 其生物量与TND、TN、WT、NH4+-N、CODMn呈显著正相关, 与Turb、DO、TP、TDP、NO3--N呈正相关, 与TD、SS呈显著负相关。氮磷等营养元素是影响浮游植物的主要因素[20-21], 鄱阳湖南部的子湖军山湖相关研究表明[22-23], 近年来TN、TP浓度的上升为蓝藻的大量繁殖提供了充足的营养盐, 硝态氮含量对绿藻等浮游植物群落分布有明显影响。在其他水质较好湖泊仙女湖的相关研究中发现[18], TN、TP均是驱动浮鞘丝藻等在浑浊的富营养化水体中具生存优势的藻类生物量变动的重要因素, 该类型藻类在夏季和秋季在仙女湖具有明显优势。同时, 较高水温可诱发蓝藻大量繁殖, 蓝藻较其他藻类更能耐住25℃高水温, 在高水温环境下体现出更强大的竞争优势[24]。在7月份的珠湖, TN、TP、TDP、TDN、CODMn、PO43--P、NO3--N等主要营养盐物种的浓度均在2018年7月份最大(见表5), 较高含量的氮、磷、化学需氧量等营养物为隐球藻、伪鱼腥藻、微囊藻、卷曲鱼腥藻等优势种提供了丰富的便于利用的营养物质, 利于生长繁殖, 藻类生物量增加明显。因此, 在夏季珠湖这类阻隔形成的, 水体较为稳定的湖泊, TND、TN、WT、NH4+-N、CODMn等环境因子为隐球藻、伪鱼腥藻、微囊藻、卷曲鱼腥藻等优势种生长的主要环境影响因子。现场调研也发现珠湖周边农业与渔业生产与城镇生活污染源为珠湖水体提供了大量营养物质, 外珠湖DS03白沙洲湖区, 内珠湖DS06、DS09湖区为水产养殖的主要区域, SW11湖区为四十里街集镇污水的主要汇入湖区, 受到外源输入直接影响, 以上湖区富营养化程度也较高于其他湖区。戴国飞等人在鄱阳湖南矶湿地的有关研究也提出了类似结论, 当湖区温度较高时, 湖区开发方式将直接在短期内加剧碟形湖内营养盐升高, 部分湖区营养盐浓度局部发生浓缩升高, 形成大量浅水高营养盐碟形湖面, 为蓝藻类浮游植物生长提供了优良的特殊生境[25]。在1月份和12月份, 珠湖小环藻、丛球韦斯(丝)藻、华丽四星藻、锥囊藻、卵形隐藻、尖尾蓝隐藻生物量与SS箭头方向一致, 其生物量与SS呈显著正相关, 与TND、TN、WT、NH4+-N、CODMn呈显著负相关。SS成为珠湖冬季小环藻藻类生长的主要影响因素, 可能主要是因为1月和12月珠湖悬浮物含量较高, 悬浮颗粒对藻类絮凝沉降作用得到增强[26]。此外, 水温的变化更多地影响了藻类季节演替, 1月和12月珠湖较低的水温比夏季更不适宜浮游植物大量生长, 温度影响浮游植物生长率, 进而影响其生物量、群落结构变化[27]。
致谢:感谢南昌大学张欢博士在野外采样部分工作中的指导, 特别感谢国家钨与稀土产品质量监督检验中心在水质样品分析上提供的协助。
[1] 黄成, 岳强, 史元康, 等. 广东省韶关市3 座大中型饮用水水库浮游植物功能类群特征[J]. 生态科学, 2014, 33(2): 282–287.
[2] REYNOLDS C S. The ecology of planktonic blue-green algae in the North Shropshire Meres[J]. Field Studies, 1971, 3(2): 409–431.
[3] JUDIT PADISÁK, CROSSETTI L O, NASELLI-FLORES L. Use and misuse in the application of the phytoplankton functional classification: a critical review with updates[J]. Hydrobiologia, 2009, 621(1): 1–19.
[4] TOLOTTI M, MANCA M, ANGELI N et al. Phytoplankton and zooplankton associations in a set of alpine high altitude lakes: Geographic distribution and ecology[J]. Hydrobiologia, 2006, 562(1): 99–122.
[5] ZHANG L, YAO X L, TANG C J et al. Influence of long-term inundation and nutrient addition on denitrification in sandy wetland sediments from Poyang Lake, a large shallow subtropical lake in China[J].Environmental Pollution, 2016, 219: 440–449.
[6] JIANG Y H, XIE H Q, ZHANG H et al. Dissolved heavy metals distribution and risk assessment in the Le'an River subjected to violent mining activities[J]. Polish Journal of Environmental Studies, 2018, 27(4): 1559–1572.
[7] XIAO L J, ZHU Y, YANG Y et al. Species-based classification reveals spatial processes of phytoplankton meta-communities better than functional group approaches: A case study from three freshwater lake regions in China[J]. Hydrobiologia, 2018, 811(1): 313–324.
[8] SANTANA L M, CROSSETTI L O, FERRAGUT C. Ecological status assessment of tropical reservoirs through the assemblage index of phytoplankton functional groups[J]. Brazilian Journal of Botany, 2017, 40(3): 695–704.
[9] 钱奎梅, 刘宝贵, 陈宇炜. 鄱阳湖浮游植物功能群的长期变化特征(2009—2016年)[J]. 湖泊科学, 2019, 31(4): 1035–1044.
[10] 胡鸿钧, 魏印心. 中国淡水藻类-系统、分类及生态[M]. 北京: 科学出版社, 2006.
[11] HUGHES B D. The influence of factors other than pollution on the value of Shannon diversity index for benthic macro-invertebrates in streams[J]. Water Research, 1978, 12: 359–364.
[12] LAN F S, PETER J F. A tribute to Claude Shannon (1916-2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon-Wiener’ Index[J]. Globle Ecology and Biogegraphy, 2003, 12: 177– 179.
[13] MARGALEF D R. Inpersproctivesi marine biology (A Buzzati-Traversoed)[M]. California: University California Press, 1958: 323–347.
[14] WASHINGTON H G. Diversity and community comparison in dices[J]. Water Research, 1984, 18(6): 653– 694.
[15] 高远, 慈海鑫, 亓树财, 等. 沂河4条支流浮游植物多样性季节动态与水质评价[J]. 环境科学研究, 2009, 22(2): 176–180.
[16] 金相灿, 屠清瑛. 湖泊富营养化调查规范(第二版)[M]. 北京:中国环境科学出版社,1990.
[17] 戴国飞, 刘慧丽, 张伟, 等. 江西柘林湖富营养化现状与藻类时空分布特征[J]. 湖泊科学, 2015, 27(2): 275–281.
[18] 于晨, 张萌, 陈宏文, 等. 仙女湖及入湖河流浮游植物功能类群与环境因子的相互关系[J]. 水生生物学报, 2018, 42(3): 622–634.
[19] 赵秀侠, 卢文轩, 李静, 等. 安徽城东湖浮游植物群落结构与水质评价[J]. 生态科学, 2020, 39(5): 187–196.
[20] JACQUEMIN C, BERTRAND C, OURSEL B et al. Growth rate of alpine phytoplankton assemblages from contrasting watersheds and N-deposition regimes exposed to nitrogen and phosphorus enrichments[J].Freshwater Biology, 2018, 63(10): 1326–1339.
[21] WINDER M, SOMMER U. Phytoplankton response to a changing climate[J]. Hydrobiologia, 2012, 698(1): 5–16.
[22] 陶志英, 余智杰, 陈文静, 等. 军山湖浮游植物群落结构及其与环境因子的关系[J]. 江苏农业科学, 2018, 46(14): 274–279.
[23] 刘霞, 钱奎梅, 谭国良, 等. 鄱阳湖阻隔湖泊浮游植物群落结构演化特征:以军山湖为例[J]. 环境科学, 2014, 35(7): 2557–2564.
[24] 史小丽, 王凤平, 蒋丽娟, 等. 扰动对外源性磷在模拟水生态系统中迁移的影响[J]. 中国环境科学, 2002, 22(6): 537–541.
[25] 戴国飞, 张萌, 冯明雷, 等. 鄱阳湖南矶湿地自然保护区蓝藻水华状况与成因分析[J]. 生态科学, 2015, 34(4): 26–30.
[26] 王艺兵, 侯泽英, 叶碧碧, 等. 鄱阳湖浮游植物时空变化特征及影响因素分析[J]. 环境科学学报, 2015, 35(5) : 1310–1317.
[27] 李季东, 肖利娟, 胡韧, 等. 沙河水库冬季浮游植物群落结构与水华分析[J]. 生态科学, 2011, 30(5): 500–506.
Spatiotemporal variation of phytoplankton communities and their relationship with environmental factors in Zhuhu Lake of Poyang Lake
ZHOU Min1,2, HE Liang3, HUANG Jianmei1,2, ZHANG Meng1,2,*, WANG Qipei1,2, WAN Binhao1,2, XIONG Mingri1,2, LIU Zugen1,2*
1. Jiangxi Academy of Eco-environmental Sciences And Planning, Nanchang 330039, China 2. Environmental Pollution Engineering Center of Jiangxi, Nanchang 330039, China 3. School of Resources, Environmental & Chemical Engineering and Key Laboratory of Poyang Lake Environment and Resource Utilization of Ministry of Education, Nanchang University, Nanchang 330031, China
Zhuhu Lake is not only the sole large sublake that has the function of important drinking water supplyment in Poyang Lake, but also one of national good-quality lakes. However, it is not well-documented on the structure and spatiotemporal variation of algea communities in Zhuhu Lake. Our study examined the variation patterns and the spatial distribution characteristics of phytoplankton. Field sampling of 13 sites was conducted and the phytoplankton samples were collected in Zhuhu Lake in January, July and December of 2018. The 79 taxa of phytoplankton belonging to 58 genera 7 phyla were identified, of which the most abundant phyla were Chlorophyta, followed by Bacillariophyta. The average phytoplankton biomass and cell abundance were 4.80 mg·L-1and 2.56×106cells·L-1. The distribution of phytoplankton was spatial-dependent in the lake. The dominance of cyanobacteria in the Tuanlin-Sishilijie lake area was higher than in other areas of Zhuhu Lake in January and July, and the dominance of diatoms in both Shuanggang and county water-source area and cryptophyta dominance in Tuanlin-Sishilijie area were higher than in other areas of Zhuhu Lake in December. The results of water quality biological evaluation showed that it was in the state of moderate pollution at present. RDA analysis showed that TND, TN, CODMn, TP and WT explained 93.96% of the relationship between species and environmental factors, and TND was the main environmental factor affecting phytoplankton biomass.
Zhuhu Lake; sublake of Poyang Lake; phytoplankton; water quality evaluation; diversity index; redundancy analysis
周慜, 何亮, 黄建美, 等. 鄱阳湖-珠湖浮游植物群落结构时空变化特征与环境因子的关系[J]. 生态科学, 2021, 40(4): 36–46.
ZHOU Min, HE Liang, HUANG Jianmei, et al. Spatiotemporal variation of phytoplankton communities and their relationship with environmental factors in Zhuhu Lake of Poyang Lake[J]. Ecological Science, 2021, 40(4): 36–46.
10.14108/j.cnki.1008-8873.2021.04.005
Q178.51+3
A
1008-8873(2021)04-036-11
2020-11-18;
2020-12-10
中国博士后科学基金资助(2020T130268, 2020M671968); 江西省主要学科学术和技术带头人资助项目(20182BCB22016); 江西省科技计划项目(20161BBG70098); 江西省重点研发计划项目(20203BBG72W007)
周慜(1987—), 男, 江西资溪人, 硕士, 工程师, 主要从事地表水与土壤污染联防联控研究, E-mail: zmtt5156@163.com
张萌, 男, 湖北京山人, 博士, 研究员, 主要从事湖沼与流域环境科学, E-mail: tomdeshiye@126.com
