气候变化背景下南美蟛蜞菊在中国的潜在适生区预测
2021-08-20肖雨沙郑洁宁李红春宋莉英
肖雨沙, 郑洁宁, 李红春, 宋莉英
气候变化背景下南美蟛蜞菊在中国的潜在适生区预测
肖雨沙, 郑洁宁, 李红春, 宋莉英*
广州大学生命科学学院, 广州 510006
采用最大熵物种分布模型(MaxEnt)对南美蟛蜞菊当前、未来2050、2070年在我国适生区分布情况进行预测, 探究影响南美蟛蜞菊分布的主要环境因子。模型AUC值达到0.9以上, 表明预测结果可信度较高。模型预测结果显示, 19个环境因子中贡献率较高的前四位依次是最暖季度降水量(bio 18)、最湿月降水量(bio 13)、最干月降水量(bio 14)、最湿季度降水量(bio 16), 可以看出降水量是影响南美蟛蜞菊分布的主导因子。模型预测南美蟛蜞菊在我国总适生区面积为21.85万km2, 占国土面积的22.77%, 表明其入侵风险较高; 其中, 最佳适生区面积为6.60万km2, 与其当前分布区基本吻合。在未来气候变化下, 南美蟛蜞菊总适生区面积变化不大, 但分布格局发生变化, 最佳适生区有向西南地区转移的趋势。研究结果为该入侵种的生态风险评估及防控工作提供了重要依据。
南美蟛蜞菊; MaxEnt; 入侵植物; 潜在适生区
0 前言
随着对外交流和进出口贸易的增加, 我国已经成为世界上遭受生物入侵现象最为严重的国家之一。2017年第三届国际入侵生物学大会报告, 目前入侵我国的外来生物有六百余种, 每年的经济损失超过2000亿元人民币。因此, 对入侵生物的潜在分布区进行预测, 阻止其进入适合生存的地区, 对于有效预防和控制外来入侵种扩张显得尤为重要。
南美蟛蜞菊(), 又称三裂叶蟛蜞菊, 菊科(Asteraceae)多年生草本植物, 原产热带美洲。作为园林地被植物被许多国家频繁引进, 现已广泛分布于东南亚和太平洋许多国家和地区[1]。由于其适应性强、繁殖速度快、且化感作用强烈, 能够抑制其他物种的生长从而形成单优群落, 严重威胁当地的物种多样性。在一些热带、亚热带地区南美蟛蜞菊侵入种植园, 与作物竞争水分和营养, 导致作物减产, 被IUCN列为“世界最有害的100种外来入侵种”之一[2]。20世纪70年代南美蟛蜞菊被引入我国, 很快逃逸为野生[3–4]。目前已经成为华南地区最常见的入侵植物之一。然而, 有关该入侵种在中国的潜在危害范围尚不明确。此外, 有研究报道南美蟛蜞菊与本地蟛蜞菊()相比对高温环境表现出更强的适应性[5], 未来气候变化可能会进一步影响南美蟛蜞菊的适生区分布。刘勇涛等[6]预测在未来气候变暖背景下南美蟛蜞菊在我国的适生区会向北发生迁移, 但仅基于对美洲和我国海南省的分布数据, 没有考虑不同气候变化情境下可能存在的差异。Qin等[7]的研究则着重比较了GARP和MaxEnt两种模型对当前南美蟛蜞菊适生区的预测效果。因此, 已有的对南美蟛蜞菊适生区的研究存在一定的局限性。
对生物适生区预测的物种分布模型有多种, 常用的有BIOCLIM[8]、CLIMEX[9]、GRAP[10]、DOMAIN[11]、MaxEnt[12]等。最大熵模型(Maximum Entropy, MaxEnt)是以最大熵理论为基础的物种地理尺度空间分布模型, 由于其预测精度高、效果较好[13], 在五爪金龙()[14]、豚草()[15]、银毛龙葵()[16]、黄顶菊()[17]等外来植物适生区预测中得到广泛应用。
本研究以南美蟛蜞菊为研究对象, 结合课题组对华南多个省区的野外实地调查, 采用MaxEnt模型对南美蟛蜞菊当前及未来不同气候变化情景下在我国的潜在适生区进行预测, 探讨影响其分布的主导环境因子, 为有效预防和控制该入侵种的扩散提供理论依据。
1 材料与方法
1.1 地理分布数据收集
南美蟛蜞菊地理分布数据主要来源于(1)课题组在广东、广西、福建、云南、海南等省区野外实地调查记录; (2)中国植物标本馆(http://www.cvh.ac.cn/)和全球生物多样性信息网络(GBIF)(https://www. gbif.org/)记录; (3)公开发表论文中有关南美蟛蜞菊的分布记录。利用Google Earth核实坐标信息, 去除重复和采集地点不准确的分布数据, 共获得1997至今南美蟛蜞菊的有效分布数据703条。
1.2 气象数据收集
本研究所用气候数据均下载自全球气候数据库(http://worldclim.org/)。通过该网站获得当前(1970—2000年)、未来2050s、2070s的气候数据, 包含19个生物气候因子(见表1), 数据的精度为2.5′。未来气候采用全球大气环流谱模式CCSM4模型。根据IPCC第五次评估报告, 未来气候情景包括4种典型浓度路径(representative concentration pathway, RCP)。本研究参照张殷波等[18]的方法选择3种温室气体排放情景, 分别是低排放情景(RCP 2.6)、中排放情景(RCP 4.5)和高排放情景(RCP 8.5)。
1.3 物种分布模型预测
将南美蟛蜞菊地理分布数据和气候数据分别导入MaxEnt 3.4.1软件中。随机选取75%的分布点作为训练数据集来建立模型, 将剩余25%的分布点作为测试数据集来验证模型[19]。设置数据输出包括创建响应曲线(create response curves)、预测图输出(make pictures of predictions), 特征值设定使用默认自动特征(auto features), 其余参数设为模型默认值。
模型建立后, 利用受试者工作特征曲线(receiver operator characteristic curves, ROC curves)分析法对模型精确度进行检验[20]。以特异性为横坐标, 灵敏度为纵坐标绘制ROC曲线, 曲线下面积(area under curve, AUC)可用于评价模型模拟的准确性。AUC值范围为0—1, 数值越大表明预测效果越好。0.50—0.60时预测结果不可接受; 0.60—0.70时预测结果勉强可以接受; 0.70—0.80时预测结果为一般, 可以接受; 0.80—0.90时预测结果良好; 0.90—1.00时预测结果极佳[21]。利用模型提供的刀切法(Jackknife) 检验, 可检测各环境因子对南美蟛蜞菊分布的相对贡献率。
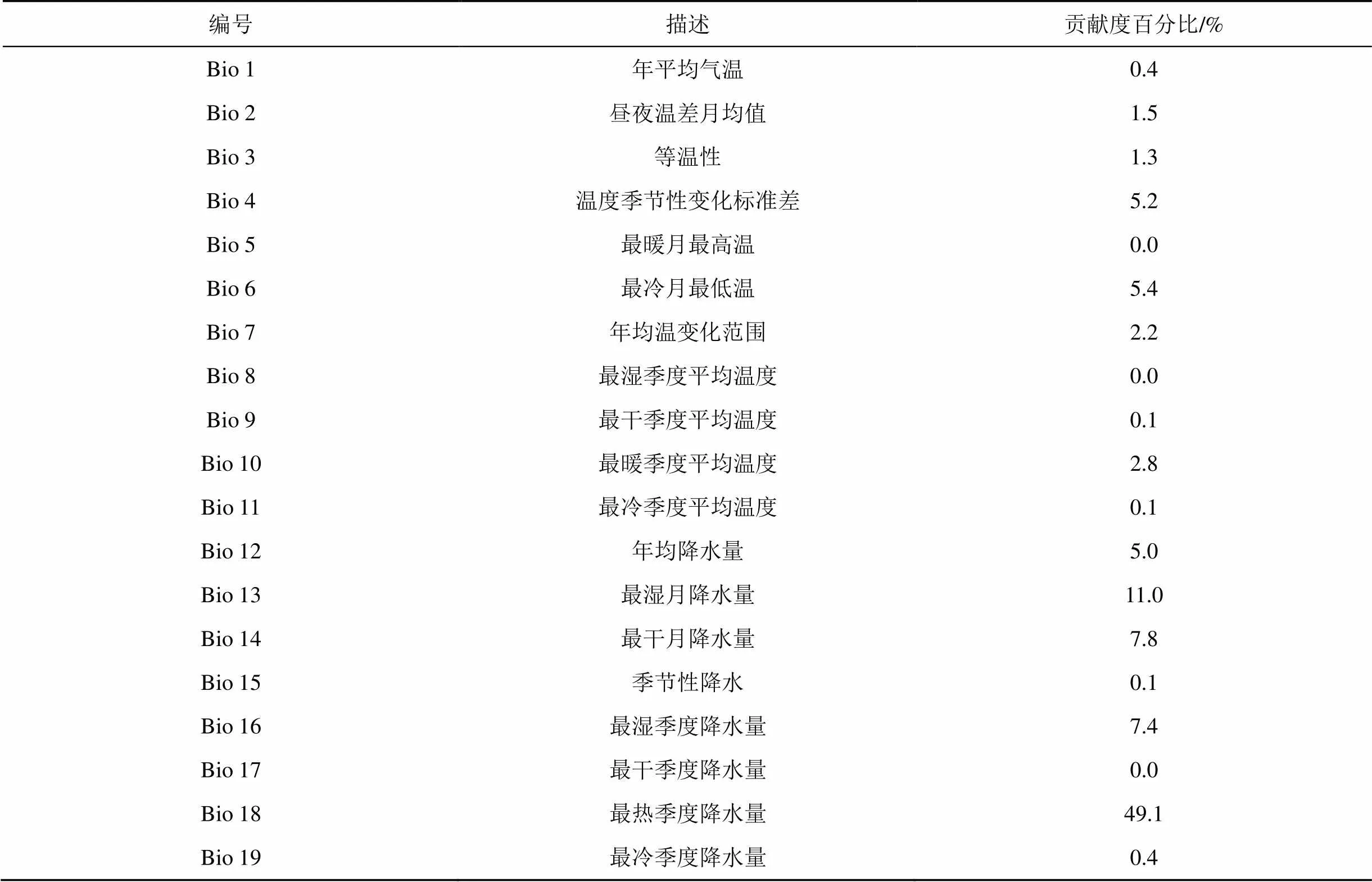
表1 环境因子描述及贡献率
1.4 预测结果处理
将MaxEnt模型预测结果ASCⅡ格式数据导入ArcGIS 10.2软件中转化为栅格数据, 得到南美蟛蜞菊的存在概率栅格图, 存在概率值在0—1之间, 值越高代表物种存在的可能性越大。利用ArcGIS 10.2软件中的重分类(Reclassify)工具中自然间断点分级法[22]可将南美蟛蜞菊在中国的分布区划分为5个等级: 非适生区(<0.08)、低适生区(0.08<<0.25)、中适生区(0.25<<0.39)、高适生区(0.39<<0.50)和最佳适生区(0.50<<1.00)。采用ArcGIS 10.2软件绘制当前和未来气候背景下南美蟛蜞菊的适生区分布图, 并通过统计像元数计算各省各级适生区的分布面积。
2 结果与分析
2.1 MaxEnt预测效果评价
模型预测结果显示训练集和测试集AUC值分别为0.968、0.971(图1), 说明MaxEnt模型对南美蟛蜞菊适生区预测结果非常好, 可信度较高。
2.2 影响南美蟛蜞菊分布的主要气候因子
刀切法对环境因子的贡献率分析结果见表1, 最暖季度降水量(bio 18)贡献率最高49.1%, 其次是最湿月降水量(bio 13)11.0%、最干月降水量(bio 14)7.8%、最湿季度降水量(bio 16)7.4%, 表明降水量是影响南美蟛蜞菊分布的主导因子。最冷月最低温(bio 6)贡献率为5.4%, 而最暖月最高温(bio 5)贡献率为0, 说明低温较高温对南美蟛蜞菊分布的影响更大。
主导气候因子对南美蟛蜞菊的影响见图2, 随着降水量的增加, 南美蟛蜞菊的分布概率迅速增加, 当最热季降水量达到330 mm左右, 最湿月降水量达到240 mm左右, 最干月降水量达到20 mm左右时, 南美蟛蜞菊的适生概率趋于稳定。最冷月最低温在8.7℃以上时南美蟛蜞菊的适生概率达到最大。

图1 MaxEnt模型ROC曲线预测结果
Figure 1 Prediction validation with ROC curve in the MaxEnt model
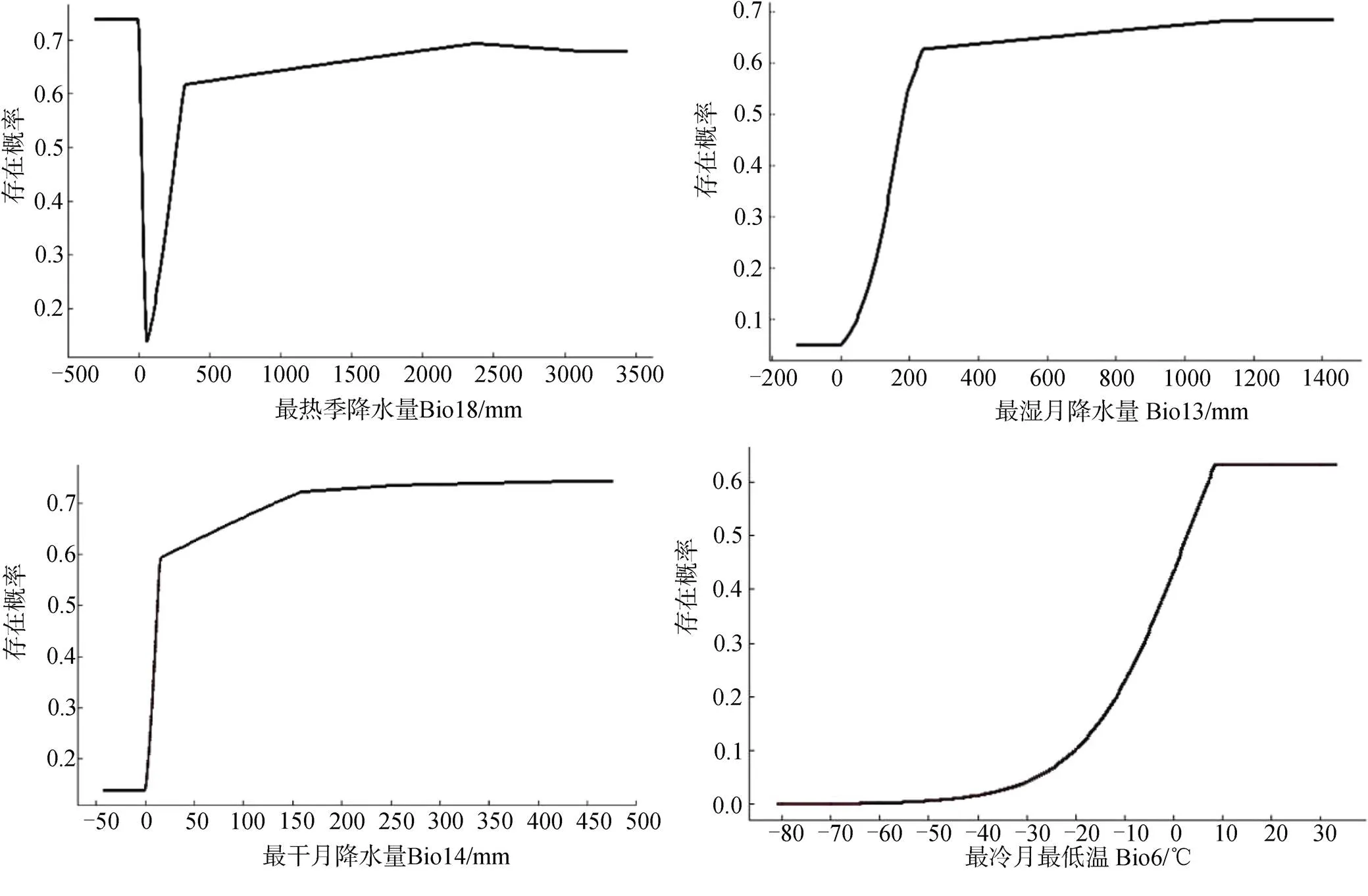
图2 主导环境因子对南美蟛蜞菊适生概率的影响曲线
Figure 2 Response curves of environmental factors affecting the fitness probability of
2.3 当前南美蟛蜞菊适生区预测
根据模型预测, 当前南美蟛蜞菊在我国的总适生区面积为21.85万km2(表2), 分布在22个省区, 占我国国土面积的22.77%。其中, 湖南、湖北、四川、贵州、安徽、河南、重庆、山东、西藏、陕西等10个省份目前没有记录, 但也被预测为适生区。最佳适生区面积为6.60万km2, 主要集中在海南、香港、澳门、广东、广西、福建、台湾、江西等8个省区, 超过该省面积65%以上(图3)。另外, 在云南、湖南、浙江、贵州和四川也有局部地区预测为最佳适生区。
2.4 未来气候条件下南美蟛蜞菊适生区预测
模型预测在RCP 2.6和RCP 4.5情景下, 2050s和2070s南美蟛蜞菊适生区总面积略有减少, 变幅在0.6—1.0万km2; 在RCP 8.5情景下, 2050s和2070s总适生区面积分别增加了0.1万km2和0.3万km2(表2)。就不同等级的适生区面积变化来看, 最佳和低适生区面积有减小的趋势, 而高适生区面积普遍有所增加(表2, 图4)。与当前最佳适生区范围相比, 未来最佳适生区有向西南部缩减的趋势,其中贵州最佳适生区面积有显著增加。
3 讨论
3.1 MaxEnt模型预测南美蟛蜞菊适生区的准确性
MaxEnt模型是基于已知物种的地理分布点和环境因子数据对其潜在适生区进行预测, 已知物种分布点数据越多越准确, 对模型的构建越有利, 精确度也越高。本研究对包括海南省在内的5个省区南美蟛蜞菊的分布情况进行了实地调查, 结合各种数据库和文献收集, 共获得南美蟛蜞菊在全球及中国分布的记录703条, 最大限度的保证了模型预测的准确性。本研究利用MaxEnt模型对南美蟛蜞菊适生区进行预测, 其AUC值大于0.9, 显示预测结果极佳。
3.2 影响南美蟛蜞菊适生区分布的主要环境因子
在19个环境因子中贡献率较高的前4个环境因子均与降水量有关, 表明降水量对于南美蟛蜞菊的分布影响较大。这与熊中人等[23]对天山花楸()的研究结果一致, 影响天山花楸分布的主要气候因子为降水和气温, 其中降水因子更为重要。本研究结果还发现, 最冷月最低温(bio 6)的贡献率高于最暖月最高温(bio 5), 表明低温较高温对南美蟛蜞菊限制作用更大。这与孙蓓育等[24]的研究结果一致。气候变化是促进生物入侵决定物种分布的重要因素[25]。在未来20—100年, 中国地表气温升高明显, 冬、春季增暖大于夏、秋季, 同时降水量呈增加趋势[26]。受气候变暖及降水量增加的影响, 预测未来南美蟛蜞菊的分布格局也会随之发生变化。

表2 南美蟛蜞菊在中国潜在适生区面积预测(单位/万km2)

图3 当前气候背景下南美蟛蜞菊潜在适生区分布
Figure 3 Distribution of potential suitable area ofunder current climate scenario
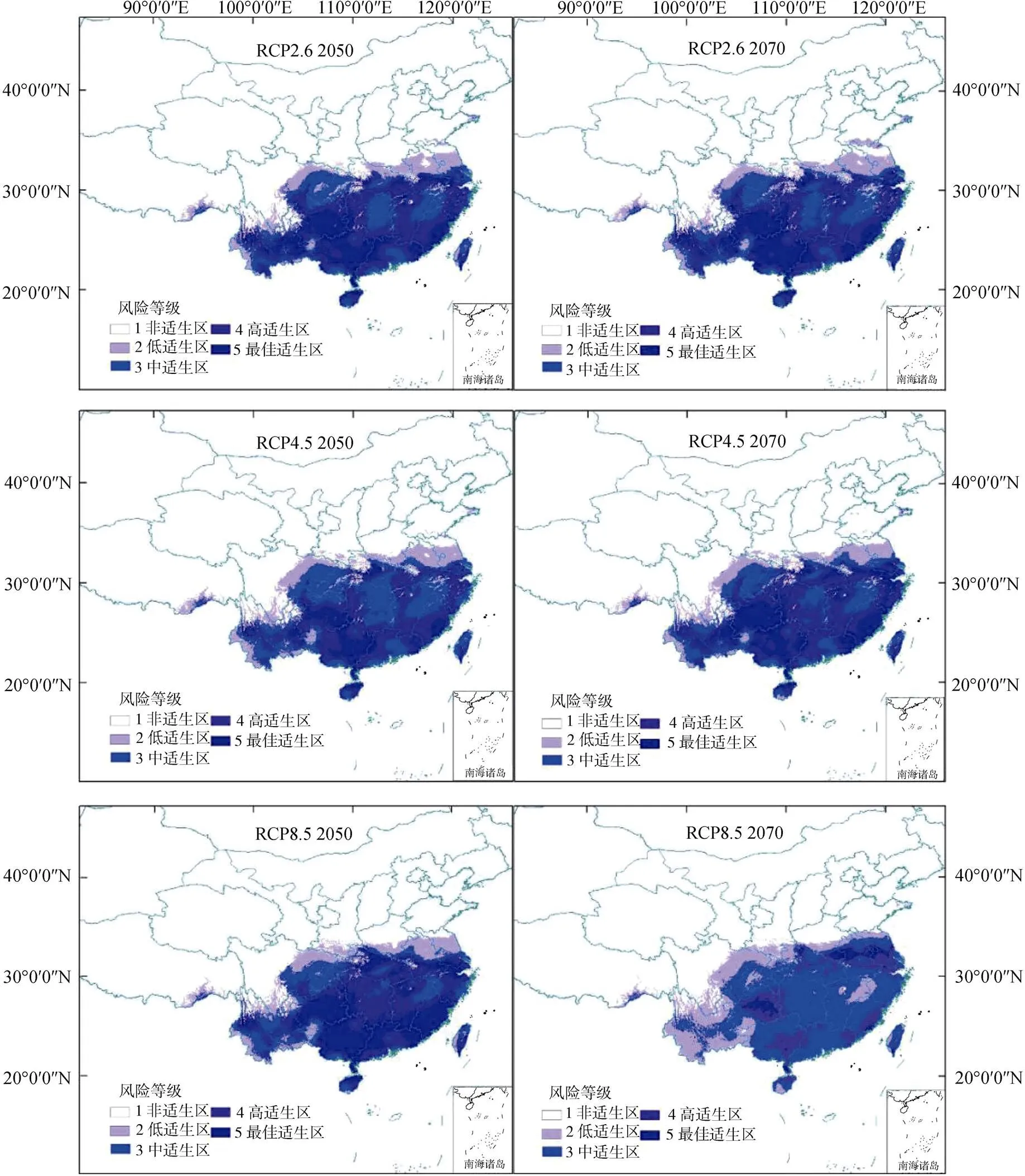
图4 2050、2070年代南美蟛蜞菊不同情景下预测适生区分布
Figure 4 Distribution of potential suitable area ofunder different climate scenarios in 2050s and 2070s
3.3 南美蟛蜞菊当前适生区分布
根据实地调查和数据库及文献记录, 当前南美蟛蜞菊在我国的分布主要集中在海南、广东、广西、福建、台湾、云南、浙江、江西等省份, 与MaxEnt模型预测当前最佳适生区基本吻合。与刘勇涛等[6]的预测结果相比, 本研究预测的当前南美蟛蜞菊在我国的适生区范围更大, 分布在22个省区, 总适生区面积21.85万km2。主要原因是模型预测时增加了海南省以外的其他省区的分布记录。这一结果提示我们, 虽然南美蟛蜞菊目前暂未被国家环保总局列入危害严重的外来入侵生物目录, 但其潜在适生区范围广、涉及省份多, 入侵风险极高。本研究中, 湖南、贵州和四川部分地区预测为当前最佳适生区, 这与Qin等[7]的预测结果类似, 而这些地区目前暂时未有南美蟛蜞菊的分布记录, 管理部门应当在这些地区限制引入、加强监测, 一旦有大面积扩散的趋势应及时予以清除。
3.4 未来气候下南美蟛蜞菊适生区的变化
由于外来入侵植物通常具有较高的环境适应性, 一般认为气候变暖会加速外来植物的入侵扩散, 并且有向高纬度地区迁移的趋势[27–28]。然而, 也有不少相反的案例。龚岩岩等[14]研究表明, 气候变化抑制了五爪金龙在中国的入侵扩散。本研究预测, 南美蟛蜞菊在未来适生区范围和面积变化不大, 仅在RCP 8.5情景下, 2050s和2070s总适生区面积分别增加了0.1万km2和0.3万km2。这一结果表明, 温室气体高排放情景下有利于南美蟛蜞菊的入侵扩散。因此, 控制温室气体排放对于减缓入侵植物扩散仍然十分必要。
未来南美蟛蜞菊适生区的变化主要体现在最佳适生区面积缩小, 而中、高适生区面积增加。其原因可能是气候变暖导致最佳适生区与中、高适生区气温差异变小, 气候条件趋于一致。综合模型对未来3种情景模式下南美蟛蜞菊适生区的预测情况来看, 最佳适生区有向西南部转移的趋势, 尤其是贵州省最佳适生区分布面积有显著增加, 这一地区应当作为南美蟛蜞菊的重点监测区域进行防控管理。
综合本研究的预测结果来看: 降水量是影响南美蟛蜞菊分布的主导因子; 南美蟛蜞菊当前在我国的适生区范围遍布22个省区, 入侵风险较高; 未来气候变化下, 南美蟛蜞菊总适生区面积变化不大, 但分布格局有向西南部转移的趋势。本研究按照大多数学者的方法选取了对物种分布影响最大的19个气候因子, 今后在建模过程中加入土壤、地形、人类活动密度等影响因素, 可进一步增加模型预测的准确性。
[1] THAMAN R R.: daisy invader of the pacific islands[M]. Suva: University of the South Pacific, 1999: 2–8.
[2] IUCN. 100 of the world’s worst invasive alien species[M]. Auckland: Invasive Species Specialist Group, 2001: 3–11.
[3] 李振宇, 解焱. 中国入侵种[M]. 北京: 中国林业出版社, 2002: 174–175.
[4] 吴彦琼, 胡玉佳, 陈江宁. 外来植物南美蟛蜞菊的繁殖特性[J]. 中山大学学报(自然科学版), 2005, 44(6): 93–96.
[5] SONG L Y, LI C H, PENG S L. Elevated CO2increases energy use efficiency of invasiveover its indigenous congener[J]. Biological Invasions, 2010, 12(5): 1221–1230.
[6] 刘勇涛, 戴志聪, 薛永来, 等. 外来入侵植物南美蟛蜞菊在中国的适生区预测[J]. 广东农业科学, 2013, 40(14): 174–178.
[7] QIN Z, ZHANG J E, DITOMMASO A, et al. Predicting invasions of(L.) Hitchc. with Maxent and GARP models[J]. Journal of Plant Research, 2015, 128(5): 763–775.
[8] 王娟, 倪健. 中国北方温带地区5种锦鸡儿植物的分布模拟[J]. 植物生态学报, 2009, 33(1): 12–24.
[9] JUNG J M, LEE W H, JUNG S. Insect distribution in response to climate change based on a model: review of function and use of CLIMEX[J]. Entomological Research, 2016, 46(4): 223–235.
[10] STOCKWELL D, PETERS D. The GARP modelling system: problems and solutions to automated spatial prediction[J]. International Journal of Geographical Information Systems, 1999, 13(2): 143–158.
[11] HIJMANS R J, GRAHAM C H. The ability of climate envelope models to predict the effect of climate change on species distributions[J]. Global Change Biology, 2006, 12(12): 2272–2281.
[12] PHILLIPS S J, SCHAPIRE R E. A maximum entropy approach to species distribution modeling[C]∥Proceedings of 21st International Conference on Machine Learning. ACM, 2004: 655–662.
[13] 段义忠, 鱼慧, 王海涛, 等. 孑遗濒危植物四合木()的地理分布与潜在适生区预测[J]. 植物科学学报, 2019, 37(3): 337–347.
[14] 龚岩岩, 贾晓霞, 朱倩倩, 等. 气候变化背景下五爪金龙在中国的潜在适生区预测[J]. 西北农林科技大学学报 (自然科学版), 2018, 46(6): 116–123.
[15] 柳晓燕, 李俊生, 赵彩云, 等. 基于MAXENT模型和ArcGIS预测豚草在中国的潜在适生区[J]. 植物保护学报, 2016, 43(6): 1041–1048.
[16] 王瑞, 万方浩. 入侵植物银毛龙葵在中国的适生区预测与早期监测预警[J]. 生态学杂志, 2016, 35(7): 1697–1703.
[17] 曹向锋, 钱国良, 胡白石, 等. 采用生态位模型预测黄顶菊在中国的潜在适生区[J]. 应用生态学报, 2010, 21(12): 3063–3069.
[18] 张殷波, 高晨虹, 秦浩. 山西翅果油树的适生区预测及其对气候变化的响应[J]. 应用生态学报, 2018, 29(4): 1156–1162.
[19] ELITH J, PHILLIPS S J, HASTIE T, et al. A statistical explanation of MaxEnt for ecologists[J]. Diversity and Distributions, 2011, 17(1): 43–57.
[20] 王运生. ROC曲线分析在评价入侵物种分布模型中的应用[J]. 生物多样性, 2007, 15(4): 365–372.
[21] ARAúJO M B, PEARSON R G, THUILLER W, et al. Validation of species–climate impact models under climate change[J]. Global Change Biology, 2005, 11(9): 1504–1513.
[22] 梁红艳, 姜效雷, 孔玉华, 等. 气候变暖背景下春兰和蕙兰的适生区分布预测[J]. 生态学报, 2018, 38(23): 8345– 8353.
[23] 熊中人, 张晓晨, 邹旭, 等. 中国天山花楸适生区预测及其对气候变化的响应[J]. 生态科学, 2019, 38(5): 44–51.
[24] 孙蓓育, 孙兰兰, 宋莉英, 等. 入侵植物三裂叶蟛蜞菊叶片叶绿素荧光对温度的响应[J]. 华南师范大学学报(自然科学版), 2009, 41(4): 100–105.
[25] STACHOWICZ J J, TERWIN J R, WHITLATCH R B, et al. Linking climate change and biological invasions: ocean warming facilitates nonindigenous species invasions[J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(24): 15497–15500.
[26] 丁一汇, 任国玉, 赵宗慈, 等. 中国气候变化的检测及预估[J]. 沙漠与绿洲气象, 2007, 1(1): 1–10.
[27] WALTHER G R, POST E, CONVEY P, et al. Ecological responses to recent climate change[J]. Nature, 2002, 416(6879): 389–395.
[28]ROOT T L, PRICE J T, HALL K R, et al. Fingerprints of global warming on wild animals and plants[J]. Nature, 2003, 421(6918): 57–60.
Prediction of potential distribution ofin China under the background of climate change
Xiao Yusha, Zheng Jiening, LI Hongchun, SONG Liying*
School of Life Science, Guangzhou University, Guangzhou 510006, China
The distribution model of maximum entropy (MaxEnt) was used to predict potential distribution ofin China under current, 2050 and 2070 climate conditions, and to explore the main environmental factors affecting its distribution. The areas under the receiver operating characteristic curve(AUC)values were above 0.9, which indicated the high prediction accuracy. Results from the mode showed that the top four with the highest contribution rate among the 19 environmental factors were the precipitation of the warmest quarter (bio 18), precipitation of the wettest month (bio 13), precipitation of the driest month (bio 14) and precipitation of the wettest quarter (bio 16), which suggested that precipitation was the dominant factor affecting the distribution of. The model predicted that the potential distribution area ofwas 218500 km2, accounting for 22.77% of the total land area in China, indicating its high invasive risk. The best suitable area ofwas 66000 km2, which was basically consistent with its current distribution area. With the climate change in future, the total area suitable forin China would remain about the same, while the best suitable area would tend to spread towards southwest China. The research results provided an important reference to the ecological risk assessment and control of this invasive species.
; MaxEnt; invasive plants; potential distribution
肖雨沙, 郑洁宁, 李红春, 等. 气候变化背景下南美蟛蜞菊在中国的潜在适生区预测[J]. 生态科学, 2021, 40(4): 75–82.
Xiao Yusha, Zheng Jiening, LI Hongchun, et al. Prediction of potential distribution ofin China under the background of climate change[J]. Ecological Science, 2021, 40(4): 75–82.
10.14108/j.cnki.1008-8873.2021.04.009
Q948.1
A
1008-8873(2021)04-075-08
2020-02-08;
2020-03-22
广东省自然科学基金(2018A030313478); 广州市科技计划(201707010257); 广州大学大学生创新训练项目(201811078098)
肖雨沙(1994—), 女, 贵州都匀人, 研究生, 主要从事植物入侵生态学研究, E-mail: 651484906@qq.com
宋莉英, 女, 博士, 副教授, 主要从事植物入侵生态学研究, E-mail: liying_song@126.com
