吕泗渔场两种石首鱼科鱼类营养生态学特征:来自稳定同位素的证据
2021-08-17高世科黄金玲于雯雯
高世科, 黄金玲, 于雯雯, 张 虎, 张 硕*
(1.上海海洋大学海洋科学学院,上海 201306; 2.上海海洋大学海洋文化与法律学院,上海 201306;3.江苏省海洋水产研究所,江苏 南通 226007)
吕泗渔场位于东、黄海交界处, 因其鱼类和饵料资源丰富而成为中国重要的渔场之一。近年来, 对吕泗渔场的研究主要集中在水文、沉积物及生物群落等宏观层面上[1-3], 利用稳定同位素技术的吕泗渔场近岸海域夏季主要生物营养级也有报道[4], 而基于稳定同位素技术的鱼类个体营养生态学的研究则相对较少, 尤其是一些经济价值较高的洄游性鱼类。
多数海洋鱼类在发育过程中存在食性转变的现象, 并且由于个体规格的差异导致营养级变动, 这种现象甚至比种间差异更显著[5-6]。小黄鱼(Larimichthyspolyactis)、棘头梅童鱼(Collichthyslucidus)是吕泗渔场两种比较重要的中小型经济鱼类, 当前, 这两种鱼类面临环境恶化、过度捕捞、非法渔具使用等严峻考验, 导致种群数量日渐衰退、鱼类质量下降[7]。研究表明, 食性相同的这两种鱼类营养级存在差异, 海州湾差值为0.40[8]、东黄海为0.76~0.80[9]、长江口为0.46~0.56[10], 其原因除了与海域饵料环境差异有关, 还与体长规格密不可分[11]。
以传统的胃含物分析法为基础, 开展对鱼类食性及营养级的分析已有许多报道, 但该方法工作量大, 存在较大片面性和局限性, 且只能反映个体短期内的食性特征[12]。目前, 碳(δ13C)、氮(δ15N)稳定同位素技术能够更精确地阐明生物体内的能量流动和走向。该技术不仅被用于定性分析海域生态系统的食物网结构, 其中的同位素量化指标还常被用来研究生物群落营养结构及食性特征[13]。因此, 建立鱼类个体规格与碳、氮稳定同位素比值间的关系, 对了解鱼类的生活史具有重要意义。
本研究旨在运用碳、氮稳定同位素技术, 对比、分析吕泗渔场两种经济石首鱼科的碳氮稳定同位素特征及其随个体发育的变化规律, 结合IsoSource模型, 初步阐明其食性特征及潜在的食性转变现象, 为深入研究吕泗渔场的生态系统营养动力学研究提供科学的理论基础资料。
1 材料与方法
1.1 样品采集
本研究于2018年9月在吕泗渔场近岸海域的18个不同站位(32°23.140′—32°59.504′N, 121°17.509′—122°11.161′E, 图1)进行单船底拖网调查。调查船为270 kW的单拖渔船, 平均拖速3.64 kn, 每站拖网时间25 min左右。使用标准Ⅰ型浮游生物网采集浮游动物, 使用Ⅲ型浮游生物网采集浮游植物, 分别使用底层有翼单囊底拖网[规格为125.32 m×59.1 m(36.0 m)]采集游泳生物, 同时使用采水器采集2 L水样用于测定悬浮颗粒有机物(Particle Organic Matter, POM), 以及抓斗式采泥器采集底质用于测定底质有机物(Sedimental Organic Matter, SOM), 所有样品经过冰鲜后, 运回实验室冰冻保存[14]。
图1 调查站位
1.2 样品预处理
本研究共采集57条小黄鱼和60条棘头梅童鱼作为样品(表1)。样品运回实验室, 将两种鱼类的体长组分成10个等级, 以10 mm为一个分界(表1), 而后测定基础生物学指标(体长、体重), 最后收集适量背部白肌肉在-44.6 ℃冷冻干燥后研磨, 待测[11]。
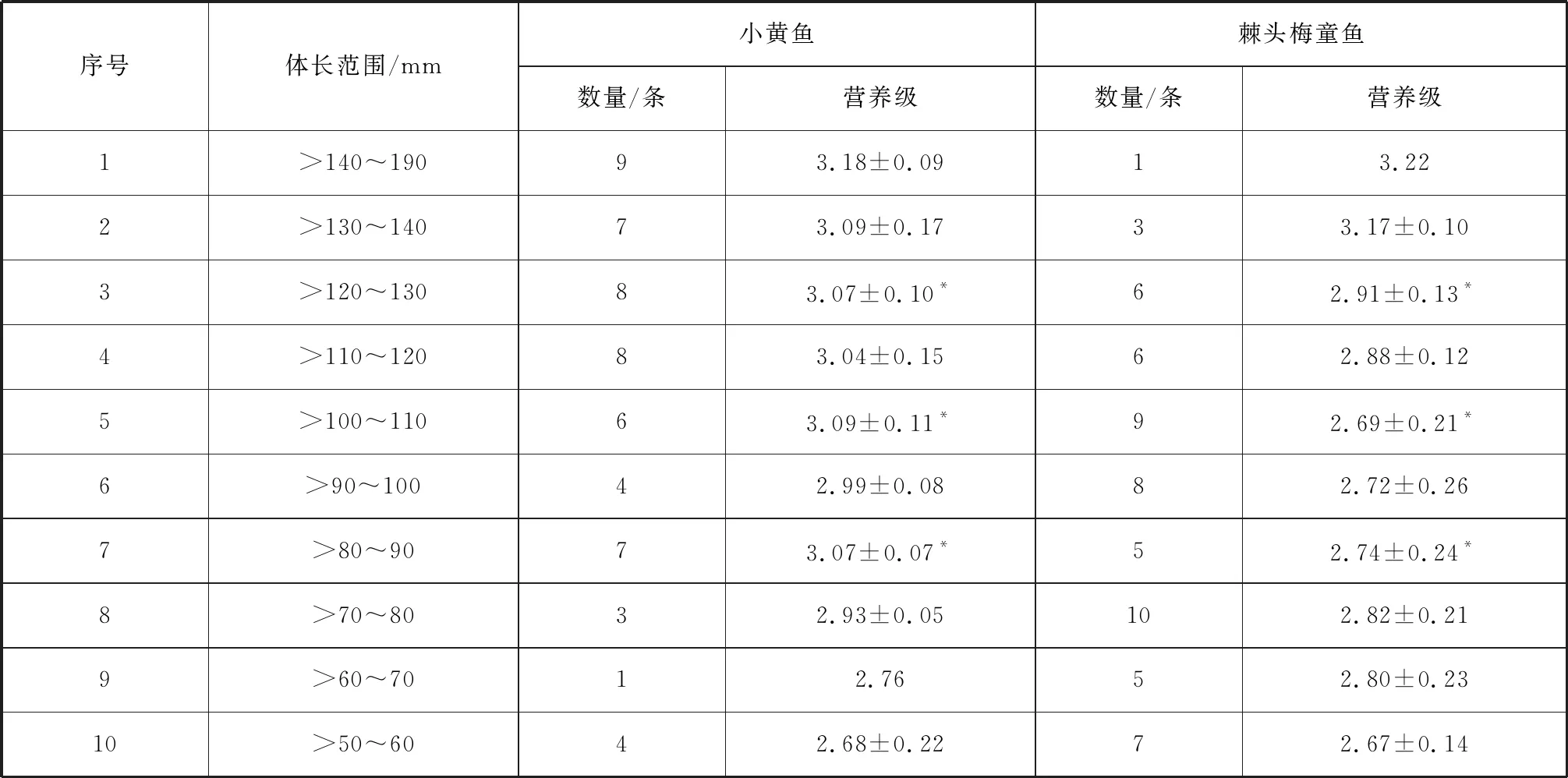
表1 不同体长小黄鱼和棘头梅童鱼的营养级
将浮游动物样品挑出杂质并清养1~2 h空胃后选出优势种, 将浮游植物样品挑出杂质并先后滤过100 μm和20 μm筛绢, 用蒸馏水静置后, 取上清液抽滤在经过500 ℃灼烧5 h的Whatman GF/F 玻璃纤维滤膜上。
水体POM用80目孔径筛绢过滤, 再抽滤至经过500 ℃预灼烧5 h的Whatman GF/F玻璃纤维滤膜上。
底质SOM在60 ℃恒温干燥24 h, 研磨后用10%盐酸酸化(去除碳酸盐干扰)[15], 同等条件下再次烘干、研磨, 经过63 μm的筛绢分筛待测。
1.3 样品分析
碳、氮稳定同位素分析在自然资源部第三海洋研究所同位素分析实验室进行, 使用德国进口的Vario Isotope Cube-Isoprime元素分析仪, 用蛋白质和乙酰苯胺来校准准确度和精密度, 以标准物质PeeDee石灰岩中的碳和大气氮作为参考标准, 用下列公式来表示碳、氮稳定同位素的自然丰度:
(1)
式(1)中:δ表示稳定同位素丰度,X为13C或者15N;λ为13C/12C或者15N/14N的比值。
营养级的确定采用Qu等(2016)重新编译、矫正过的公式[16], 如下:
(2)
式(2)中:TL表示所计算生物的营养级;δ15Nsample为系统消费者氮稳定同位素比值;δ15Nbaseline为该系统基准生物的氮稳定同位素比值;Δδ15N为相邻营养级间的富集度(Trophic Enrichment Factor), 本研究选取POM作为基线生物, 同时沿用Post(2002)研究稳定同位素计算营养级的3.4‰作为营养级富集度[17]。
1.4 IsoSource模型分析
根据质量守恒原理, 利用Phillips等(2003)[18]不断完善编译的IsoSource模型来定性分析和计算各个碳源对消费者的贡献率,原理如下:
δjM=AδjA+BδjB+CδjC
(3)
δkM=AδkA+BδkB+CδkC
(4)
(5)
式(3)至(5)中:jM、kM为消费者M的两种同位素j、k的比值;A、B、C为食物源;为食物对消费者的贡献率。在海域食物网中, 浮游植物、POM、SOM常被作为营养基础和潜在碳源来反映各类生物的潜在食源贡献比例, 因此被本研究选用。
1.5 数据处理及分析
站点图使用ArcGIS 10.3[ESRI (USA)]绘制;基于δ13C和δ15N值和标准化的欧氏距离, 使用PRIMER V5软件包进行组间连接等级聚类分析(Hierarchical Cluster Analysis);沿用Origin2018和Excel 2019进行图表的绘制;其余数据处理与分析均通过Excel 2019和SPSS 25.0(Inc., Chicago,IL, USA)来完成;Pearson相关分析和线性回归是用来确定鱼类不同体长组与δ13C、δ15N值之间的关系。使用方差分析(ANOVA)来检测两种鱼类间的同位素特征。
2 结果与讨论
2.1 两种鱼类的δ13C、 δ15N值特征
如图2所示, 两种鱼类的δ13C、δ15N值范围均较广, 小黄鱼的δ13C值范围为~18.90‰~15.40‰, 均值为(~16.50±0.66)‰,δ15N值范围为8.33‰~11.77‰, 均值为10.52±0.59‰;棘头梅童鱼的δ13C值范围为~18.67‰~12.81‰, 均值为(~16.61±0.98)‰,δ15N值范围为8.07‰~11.42‰, 均值为(9.69±0.79)‰。对所有样品进行单因素方差分析的结果表明, 不同体长组间小黄鱼和棘头梅童鱼的稳定碳同位素差异不显著(p>0.05), 而稳定氮同位素差异显著(p<0.05)。
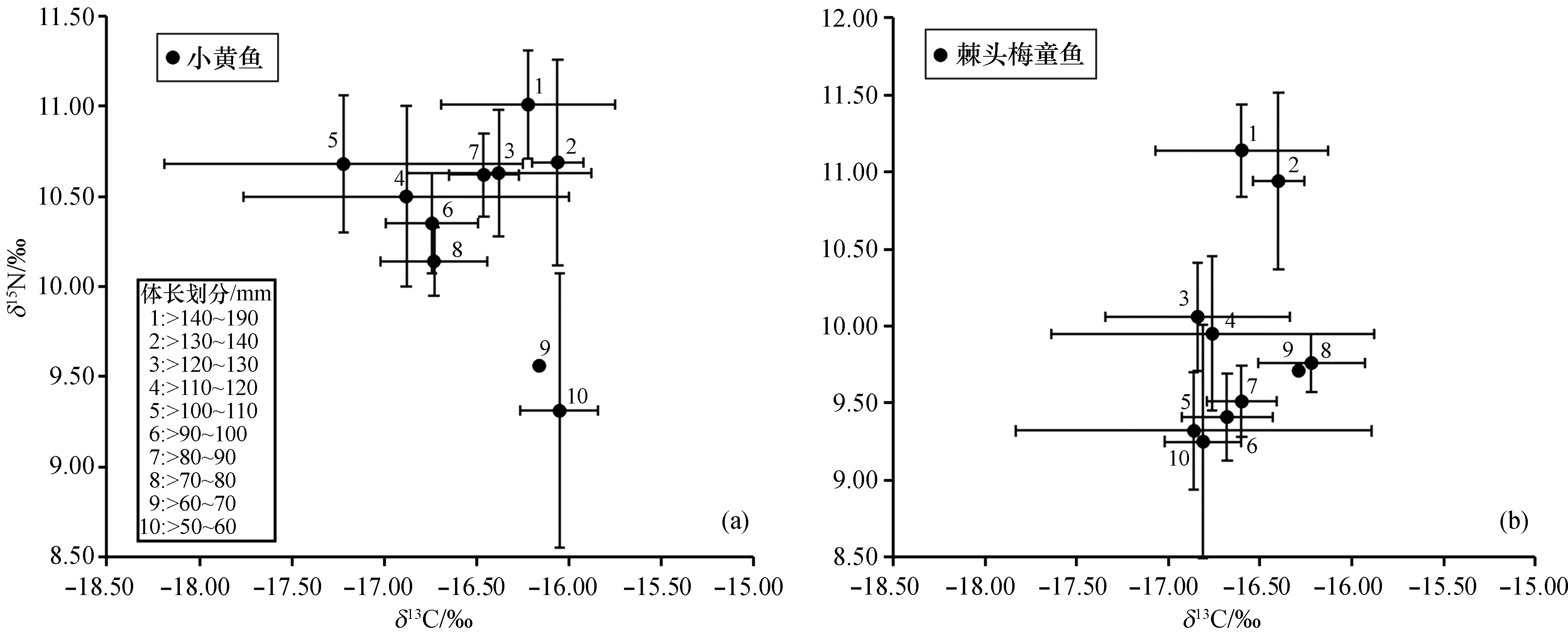
图2 小黄鱼和棘头梅童鱼的δ13C、δ15N值特征
基于δ13C和δ15N值和标准化的欧氏距离, 对两种鱼类不同体长组进行聚类分析, 结果如图3所示。小黄鱼的体长组可分成两大组, 包括>140~190 mm、>130~140 mm、>120~130 mm、>80~90 mm一组和剩余的体长组;棘头梅童鱼的体长组可分为>140~190 mm、>130~140 mm一组和剩余体长组。聚类分析的结果比较符合两种鱼类δ13C-δ15N比值分布的结果(图2)。
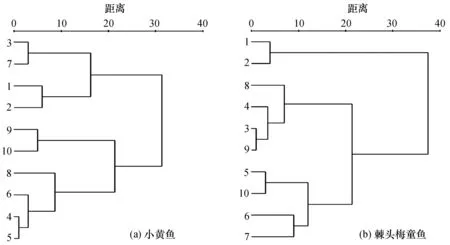
图3 两种鱼类不同体长组δ13C、δ15N值的聚类分析
两种鱼类的δ15N值随体长的增加而增大, 小黄鱼的δ15N值差值为3.44‰, 棘头梅童鱼的δ15N值差值为3.35‰。两者的差值均与Post(2002)研究消费者和食物间的氮同位素分馏时确定的δ15N富集平均值3.40‰[17]十分接近。这一结果一方面反映出随着个体增大, 生物体的δ15N值也出现增加趋势;另一方面, 不论从鱼类的个体发育还是系统发育层面上来看, 本研究的这一结论都加以验证了15N重同位素在鱼类体内富集现象的普遍性。
本研究中两种鱼类的δ15N值均是随体长增加而增大, 使用Pearson指数分析得到两组数据间的相关性较弱(0.2
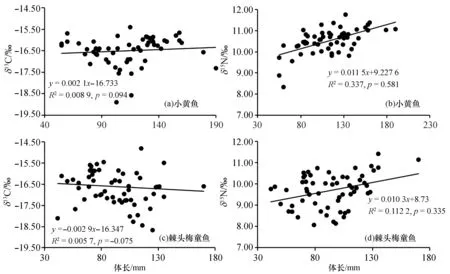
图4 小黄鱼和棘头梅童鱼不同体长组和δ13C、δ15N值之间的线性关系
2.2 两种鱼类的营养级特征
从表1可以看出, 小黄鱼的营养级范围为2.68~3.18, 平均值为2.99±0.15;棘头梅童鱼的营养级范围为2.67~3.22, 平均值为2.86±0.18。利用无重复双因素方差分析得知, 除体长为>60~70 mm、>130~140 mm和>140~190 mm的棘头梅童鱼的营养级大于小黄鱼之外, 其余体长组小黄鱼的营养级要显著高于棘头梅童鱼(p<0.05)。
使用不同的营养级公式会导致计算结果产生差异, 本研究使用POM[δ15N:(3.58±0.42)‰]作为基线和Post(2002)[17]的3.40‰作为营养级富集度计算两种鱼类的营养级。图5显示了两种鱼类不同体长组营养级增加的趋势图, 小黄鱼(2.99)比棘头梅童鱼(2.86)平均高出0.14个营养级(p<0.05)。研究表明, 小黄鱼和棘头梅童鱼早期幼鱼的食性主要以浮游生物为主, 成体后小黄鱼食性逐渐转向为以小型鱼类和虾类为主[21], 而棘头梅童鱼的栖息水层比小黄鱼高, 食性范围也比小黄鱼广, 但由于棘头梅童鱼的齿尖细, 鳃耙细长且数目多, 属于浮游生物食性鱼类[23], 因此从食性方面来看, 也能够解释本研究的结论。
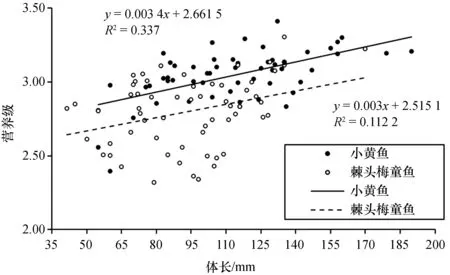
图5 不同体长小黄鱼和棘头梅童鱼营养级变化趋势图
与长江口相同体长组的小黄鱼相比, 本研究小黄鱼的营养级差异不显著(p<0.05,因长江口海域只提供δ15N值, 故沿用本研究的条件计算营养级), 但各体长组棘头梅童鱼的营养级与东海北部相比差异较显著(p>0.05), 较东海北部海域平均低了0.4个营养级[23]。产生这种差异的因素很多, 不仅与海域的饵料基础和环境因素有关, 还和研究方法相关[19]。有研究表明, 胃含物分析只能反映鱼类短时间内的摄食情况, 且分析时容易忽略一些小型、易消化的饵料种类, 可能导致营养级出现偏差[25]。
2.3 潜在碳源的δ13C、 δ15N值特征
潜在碳源的δ13C值在浮游植物的-24.27‰到SOM的-20.24‰之间变化, 均值为-22.64‰, 跨度为4.03‰;潜在碳源的δ15N值从POM的3.58‰变化到浮游植物的4.30‰, 均值为3.82‰, 跨度为0.72‰(表2)。

表2 潜在碳源的δ13C和δ15N值
利用单因素方差分析来检验不同食源之间碳稳定同位素比值的显著性差异, 结果显示, 各食源碳同位素比值之间均呈显著差异(p<0.05)。
2.4 IsoSource模型结果
以上述三大潜在碳源为基础贡献, 对两种鱼类的潜在食源贡献进行分析。结果(图6)表明, 浮游植物对小黄鱼的碳源贡献范围在15.60%(>60~70 mm)~62.20%(>100~110 mm), 均值为43.38%, 对棘头梅童鱼的碳源贡献范围为23.10%(>70~80 mm)~80.30%(>140~190 mm), 均值为36.38%。
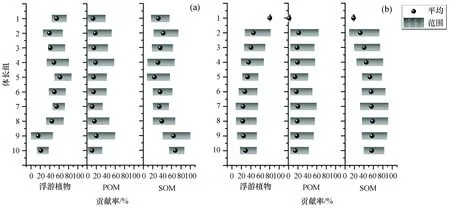
图6 三大潜在碳源对不同体长小黄鱼和棘头梅童鱼的贡献率
POM变化不明显, 对小黄鱼的碳源贡献范围为10.90%(>50~60 mm)~20.00%(>60~70 mm), 均值为15.02%, 对棘头梅童鱼的碳源贡献范围为1.80%(>140~190 mm)~21.70%(>130~140 mm), 均值为16.32%。
SOM对小黄鱼的碳源贡献范围为23.70%(>100~110 mm)~68.00%(>50~60 mm), 均值为40.66%, 对棘头梅童鱼的碳源贡献范围为17.90%(>140~190 mm)~58.30%(>70~80 mm), 均值为43.70%。因此判定小黄鱼和棘头梅童鱼的潜在碳源主要是浮游植物和SOM。
从潜在碳源的角度能够更清楚判断和说明鱼类的食性转变规律。在本研究中, 随着两种鱼类体长的增加, 浮游植物对两种鱼类的碳源贡献比例逐渐增大, 相反的, SOM的碳源贡献比例逐渐减小。尽管如此, 浮游植物和SOM对小黄鱼的碳源贡献比例在随体长增加的过程中出现波动较大的现象。研究表明, 小黄鱼主要以小型鱼类、甲壳类、头足类、矢状类和多毛类为食[21], 这与小黄鱼杂食性的食性特征分不开。郭斌等(2010)的研究表明, 当体长小于80 mm时, 小黄鱼主要以桡足类、糠虾类和少数虾类为食, 当体长大于80 mm时, 小黄鱼的食性开始转向小型鱼类和虾类[26]。本研究的结果表明, 当小黄鱼体长小于80 mm时, 浮游植物的平均碳源贡献比例为27.17%;当小黄鱼体长大于80 mm时, 浮游植物的平均碳源贡献比例为50.33%, 揭示了小黄鱼食性的潜在转变现象。而棘头梅童鱼潜在食性转变的体长临界值比小黄鱼高, 为110 mm。这一结论反映了棘头梅童鱼在生活史中转变食性的行为可能比小黄鱼晚。
POM的贡献比例最低, 魏虎进等(2014)在研究象山港海洋牧场食物网结构时发现, 相对于浮游植物和SOM, POM的相对碳源贡献率最小[27], 佐证了本研究的结论。此外, 中国各大海域POM对水生动物的碳源贡献比例均未发生太大变化[27-28], 本研究也不例外, 因此可以推测POM对两种鱼类幼体到成体生长发育过程的影响并不明显。
3 结论
综上所述, 与通过比较δ15N值及营养级的传统方法相比,结合潜在碳源对两种鱼类各体长组的影响,更能有力验证两种鱼类潜在食性的转变现象。本研究结果表明, 小黄鱼和棘头梅童鱼的潜在碳源主要是浮游植物和SOM。小黄鱼食性转变的体长临界点为80 mm, 而棘头梅童鱼为110 mm, 反映了棘头梅童鱼在生活史中转变食性行为的发生时间可能比小黄鱼晚。
