大亚湾西部海域中国毛虾种群动态变化及其对环境要素的响应
2021-08-17安丽娜欧丹云李伟文
安丽娜,王 磊,黄 浩,欧丹云,李伟文
(自然资源部第三海洋研究所,福建 厦门 361005)
大亚湾是一个半封闭性深水海湾[1],属于不规则半日潮,平均潮差为0.49 m,最大潮差为2.50 m,夏季粤东沿岸上升流的低温高盐水会侵入至湾内,并形成明显层化结构,因而大亚湾海水的环境具有明显的季节性变化[2]。近年来,冬季在大亚湾局部海区,尤其是西部近岸区域常发生毛虾暴发事件,暴发种类主要是中国毛虾(Aceteschinensis),而目前关于该区域中国毛虾种群动态的研究较少。
毛虾为樱虾科,毛虾属(Acetes)的统称。毛虾包括17个种类[3],我国北部沿海常见种主要是中国毛虾[4],而在华南亚热带海区可见日本毛虾(Acetesjaponicus)和红毛虾(Aceteserythraeus)等种类[5]。对于辽东湾的中国毛虾来说,达到产卵的初始水温在18 ℃左右,产卵盛期一般在水温达到20 ℃后出现[6]。中国毛虾的幼体发育经历4期无节幼体、3期溞状幼体、2期糠虾幼体和4期仔虾阶段[6]。近年来的研究对于中国毛虾幼体发育等生物学特征有了一定了解,但其分布与温度、盐度等环境因子关系方面的研究却并不多见。
本研究根据在大亚湾西部海域对中国毛虾的调查资料,分析了中国毛虾的种群动态变化及其对主要环境因子的响应,为该区域中国毛虾种群动态特征研究提供参考,以期为大亚湾西部近岸区域中国毛虾暴发事件提供防控思路。
1 材料与方法
1.1 采样区域
2016年10—12月,对大亚湾西部海域的表层和底层浮游植物光合色素进行了取样;2016年12月至2017年2月和2017年12月至2018年3月的每月上、中、下旬在大亚湾西部海域布设5个起始测站(D1至D5)以及5条毛虾拖网测线(L1至L5),开展了温度、盐度、营养盐及毛虾种群密度和体长等参数的取样观测,站位及测线布设如图1所示。本研究共开展了17个航次的调查,其中由于现场拖网取样获取的样品中主要以小型水母和箭虫等浮游动物为主而毛虾数量较少,因而共计有10个航次有效获取了毛虾样品。

图1 大亚湾西部海域观测航次测站布设
为便于数据比较分析,并考虑到拖网作业起始点的距离等因素,根据拖网起始点的离岸距离、南北距离和水深等要素,将起始点分为3组,分别为组1(D1)、组2(D2和D3)和组3(D4和D5),基本环境参数分析依据3组来进行。
1.2 采样方法
现场样品水样取自Niskin采水器。以47 mm直径的Whatman®GF/F滤膜 (孔径0.7 μm) 过滤水样。过滤完毕样品于干冰低温保存,回到实验室转移至-80 ℃保存,待进行测定。
毛虾拖网网具采用截面为方形的梅氏伊萨克框形网,网口面积为2.25 m2(1.5 m×1.5 m),网长13 m。网衣用网目为2 mm的尼龙筛绢制成,固定在金属网框上。拖网时间为15 min。网口系挂流量计用于计算过滤水量。
1.3 样品分析方法
温度 (T)、盐度 (S)由哈希sensION+EC5便携式多参数仪器于现场进行同步采样和测定。营养盐分析方法采用《海洋监测规范》[7]。浮游植物光合色素样品采用高效液相色谱法[8-10]进行分析。
1.4 数据统计分析方法
本研究中数据基本运算和整理是在Microsoft Office Excel下完成的。数据统计分析采用PASW®Statistics 17.0软件进行,海区数值散点图等采用ArcGIS Pro 2.4.0进行分析和绘图,散点图、柱形图等采用OriginLab OriginPro进行绘制。浮游植物类群反演由特征光合色素经CHEMTAX软件运算获得。
2 结果与讨论
2.1 温度和盐度的时空分布特征
通过比较各站位的表层温度可以看出(图2、3),在1—2月表层海水温度较低时,D1站温度明显低于D2、D3站,最大温度差可达到2.5~3.0 ℃;但在海水温度较高的12月,站位间表层温度差并不明显。D4、D5站的表层温度和盐度基本介于D1、D2、D3站之间,在12月甚至会略高于D2、D3站。从表层盐度分布(图4)可以看出,在1—2月盐度普遍高于其他时间。
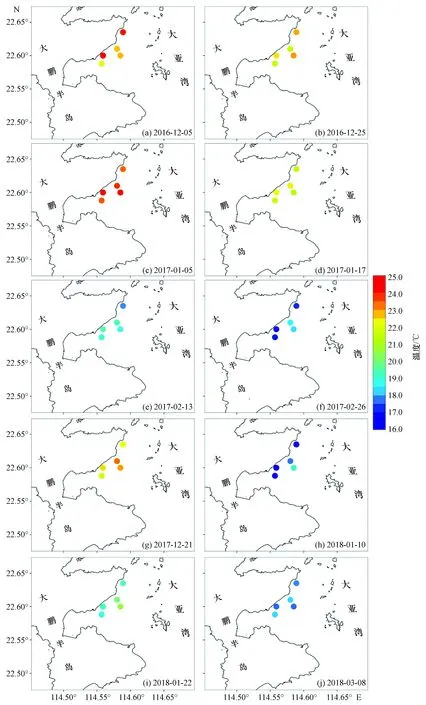
图2 大亚湾西部海域温度分布

图3 大亚湾西部海域温度变化
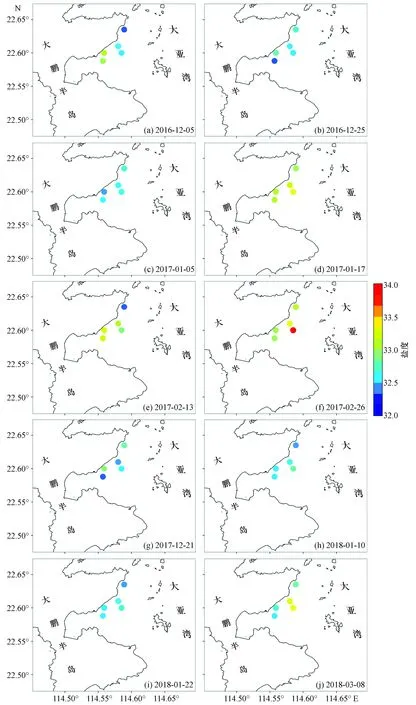
图4 大亚湾西部海域盐度分布
2.2 营养盐的分布特征
通过比较各站的表层营养盐(图5)可以看出,各类营养盐在3组中的浓度各有高低,并未呈现一定的规律。但总体上来看硅酸盐和磷酸盐在组2站位略低于其他站位。这可能是由于海水中含有余氯,导致硝酸盐形成了其他形态的化合物,而磷酸盐和硅酸盐由于在组1和组3站位处可利用的硝酸盐不足,而产生营养盐剩余。
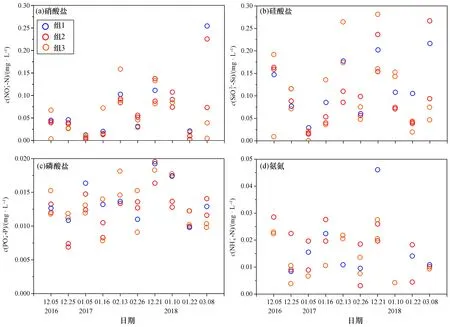
图5 大亚湾西部海域营养盐分布
2.3 浮游植物群落结构特征
对大亚湾西部海域的表层和底层浮游植物光合色素进行测定,并通过CHEMTAX软件对该区域浮游植物群落结构组成进行了反演(图6)。表层浮游植物总叶绿素a(TChl a)浓度平均值在11月最低,为(5.40±1.14) μg/dm3;10月略高于11月,为(6.38±2.23) μg/dm3;12月最高,为(9.38±2.33) μg/dm3。表层浮游植物群落组成在10月和12月均以硅藻为绝对优势类群,分别占73.56%和75.60%;而11月份浮游植物群落组成有较大不同,聚球藻(Synechococcus)占据绝对优势,对浮游植物TChl a浓度贡献量达到66.76%,而11月硅藻贡献量仅为12.48%。青绿藻(Prasinophytes)是仅次于上述两类浮游植物的第三大类群,对TChl a浓度贡献量在各月份之间差别不大,在11.93%~14.36%之间。其余类群生物量均较低。
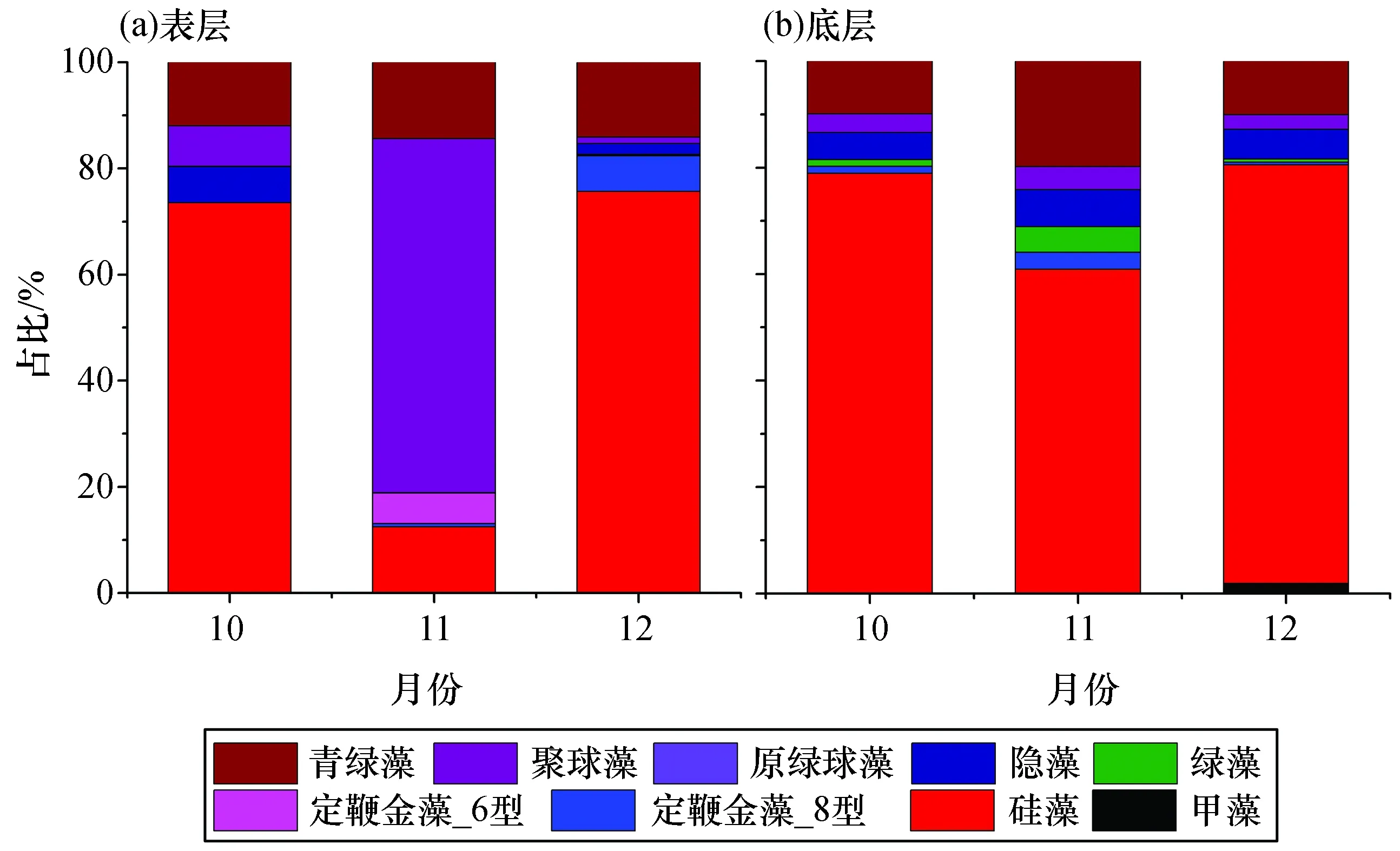
图6 2016年大亚湾西部海域浮游植物群落组成
底层浮游植物TChl a浓度平均值在10月和11月均较低,分别为(1.98±0.26)、(1.66±0.37) μg/dm3。12月TChl a浓度明显增高,为(7.53±1.85) μg/dm3。底层浮游植物群落组成在3个月份均以硅藻为绝对优势类群,分别占78.95%、60.95%和78.60%。青绿藻次之,分别占9.89%、19.79%和10.07%。其余类群生物量均较低。
由上述浮游植物群落结构组成的规律可以看出,10—12月,表层浮游植物优势类群存在着硅藻-聚球藻-硅藻的群落演替。
2.4 中国毛虾生物量分布和种群动态
2.4.1 中国毛虾个体密度分布和变化 中国毛虾个体密度最高值出现在2017年1月5日的L2测线和L3测线,分别为(38.87±2.32) 个/m3和(38.77±5.07) 个/m3,另外在2017年2月26日的L2测线中国毛虾个体密度也较高,达到(37.43±3.55) 个/m3。L1测线中国毛虾个体密度最高值出现在2017年1月17日,为(29.23±2.43) 个/m3。中国毛虾个体密度的最低值出现在2018年3月8日的L3测线,为(0.63±0.17) 个/m3(图7)。L1测线中国毛虾个体密度高于L2和L3测线的采样日期有2016年12月5日、12月25日,2017年1月17日,2018年1月10日和3月8日,其余多数采样日期均为L2、L3测线高于L1测线。
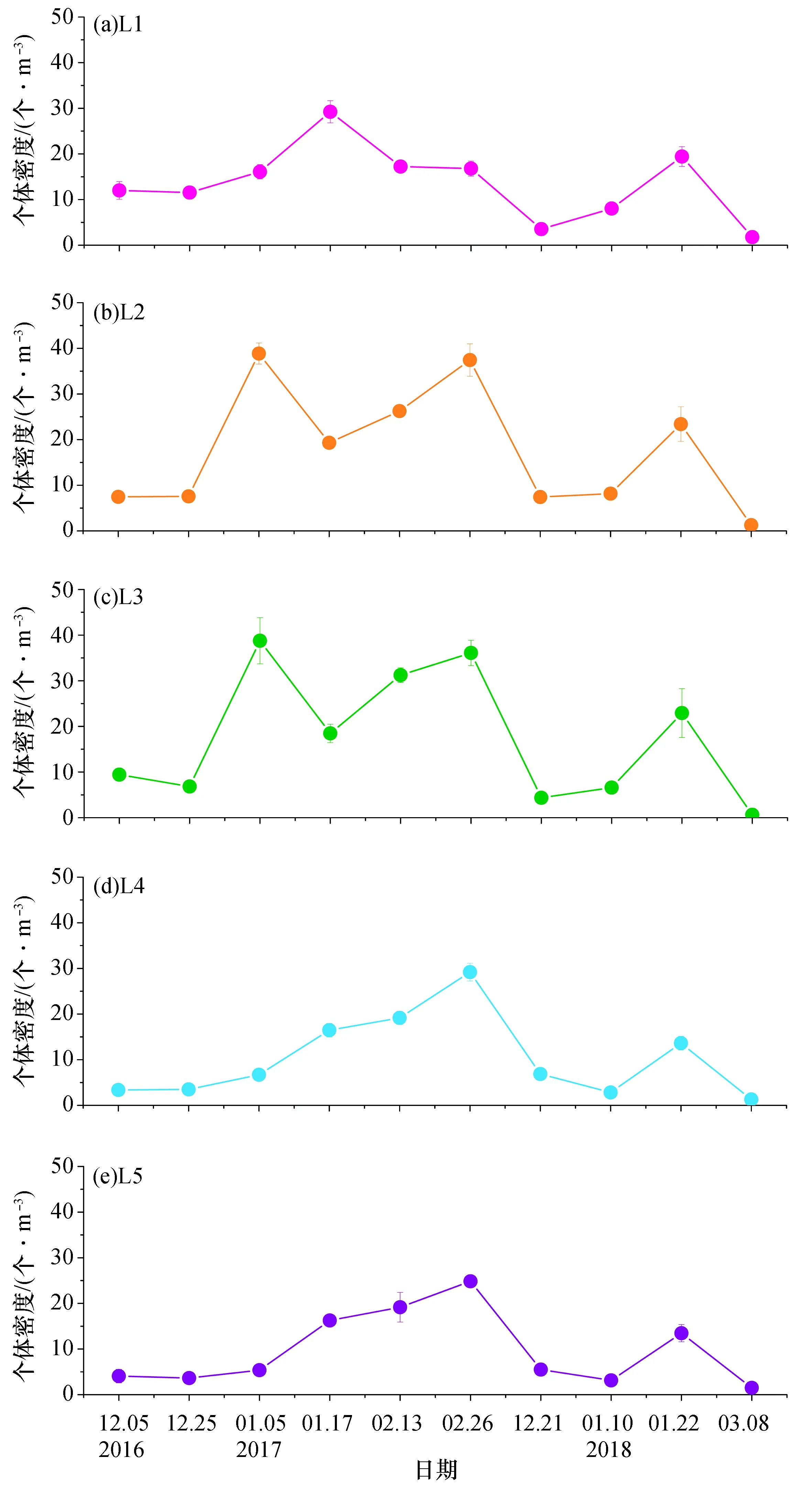
图7 大亚湾西部海域各测线中国毛虾个体密度分布
通过比较各测线中国毛虾个体密度(图8)可以看出,L1的总个体密度平均值[(13.57±7.66) 个/m3]与L2和L3[(17.63±12.98) 个/m3]无明显差异,但均略高于L4和L5[(10.00±8.13) 个/m3]。所采中国毛虾个体密度基本都是在1—2月达到最高值,并且中国毛虾个体密度与温度并无明显正相关关系(p>0.05),说明不存在温度聚集效应。
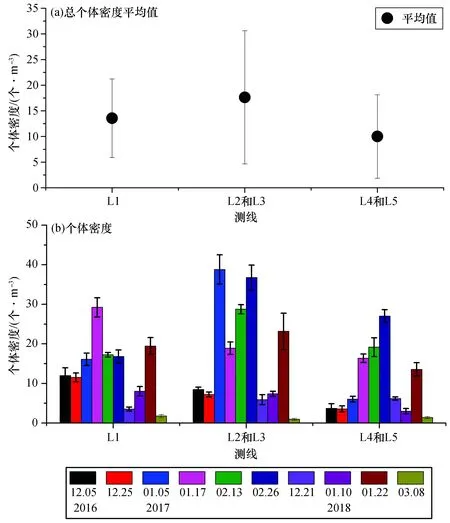
图8 大亚湾西部海域各测线中国毛虾个体密度
2.4.2 中国毛虾种群动态变化 通过测量中国毛虾体长,人为地将其划分为大型个体(体长>2.5 cm)、中型个体(体长为1.5~2.5 cm)和小型个体(体长<1.5 cm),意在分析中国毛虾种群的结构。各采样时间点中国毛虾体长平均值及占比如图9所示。在所有样品单元的分析中大型个体体长平均值为(2.78±0.12) cm,中型个体体长平均值为(2.12±0.05) cm,小型个体体长平均值为(1.24±0.13) cm。大型个体个体密度平均值为(12.13±2.54) 个/m3,中型个体个体密度平均值为(28.74±12.15) 个/m3,小型个体个体密度平均值为(26.44±9.86) 个/m3。因而可以看出中型和小型个体个体密度平均值明显高于大型个体。
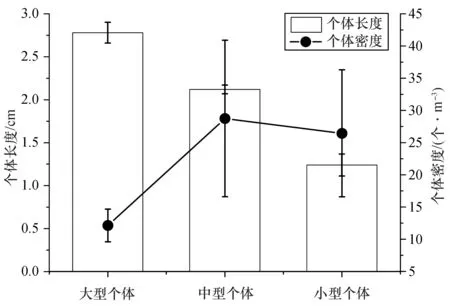
图9 大亚湾西部海域中国毛虾个体长度及个体密度平均值
如图10所示,大型个体在2017年1月5日的观测中所占比例最高,达到65.75%,而此时中国毛虾总个体密度也相对较高(51.97 个/m3)。类似的,在2017年2月26日的观测中,大型个体占比为47.6%,此时中国毛虾总个体密度约为65.39 个/m3。总体上来看,从12月至2月存在着小型-中型-大型占优势的转化,这种转化一般会在一个月内完成,因而1—2月之间往往会出现大型个体的大量产生。
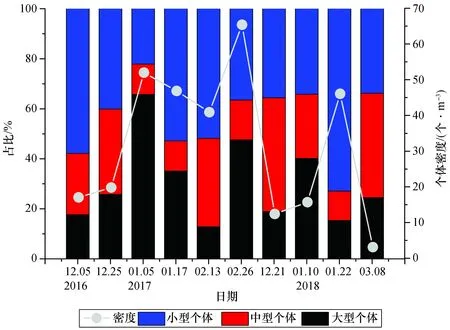
图10 大亚湾西部海域不同体长中国毛虾占比及总个体密度
影响毛虾种群动态变化的因素有很多,浮游动物等对毛虾的捕食作用,海水相关的理化因素(温度、盐度、营养盐等),海流的消长,气候变化等均会对毛虾生命过程中不同阶段产生影响[11]。曾现英等(2016)对渤海湾、莱州湾的中国毛虾调查发现,中国毛虾分布具有广温广盐性特征,其季节分布主要受温度影响,盐度对其影响只在最适温度条件下才有明显表现[12]。曾雷等(2019)利用声学探测对大亚湾临近海域的中国毛虾昼夜迁移分布研究也表明,水温是调节中国毛虾资源变动的重要因素之一[13]。前人的研究由于缺乏相关的理化因子以及与毛虾相关的生物因子,因而不能很好的说明其特征和规律。本研究在大亚湾西部海域冬季和春季所采的中国毛虾,个体密度基本都是在1—2月份达到最高值,中国毛虾个体密度与温度并无明显正相关关系(p>0.05),说明不存在着温度聚集效应。对于个体大小来说,从12月至次年2月中国毛虾存在着小型-中型-大型占优势的转化,即当温度下降时,大型个体更易成为主要优势,这可能与寒潮降温引起的藻类大量繁殖为毛虾提供了大量饵料有关,从而使其能够较为快速地生长成为较大个体。
表层浮游植物优势类群在10、11、12月存在着硅藻-聚球藻-硅藻的群落演替,中国毛虾种群的动态变化有可能因为较高的浮游植物生物量,尤其是硅藻生物量,为中国毛虾提供丰富的饵料,从而使其体现出较高的个体密度。因而影响大亚湾西部海域中国毛虾种群动态变化的主要因素是浮游植物生物量和群落结构,其为中国毛虾提供了饵料,在叶绿素a生物量较高,尤其是硅藻生物量较高时,中国毛虾个体密度较高,个体相对较大。由此初步判断影响冬季大亚湾西部海域中国毛虾种群变动的主要因素不是温度,而是浮游植物生物量和群落结构为中国毛虾提供了饵料。
3 结论
本研究依据在大亚湾西部海域进行的中国毛虾调查,分析了中国毛虾的种群动态变化及其对主要环境因子的响应。获得了如下结论:
(1)中国毛虾个体密度基本都是在1—2月达到最高值。中国毛虾个体密度与温度并无明显正相关关系,说明不存在着温度聚集效应。可能是因为具有较高的浮游植物生物量,尤其是硅藻生物量,从而使中国毛虾能够获取丰富的食物,从而体现出较高的个体密度。
(2)中国毛虾个体大小组成从12月至次年2月存在着小型-中型-大型占优势的转化。当温度下降时,大型个体更易成为主要优势,这一方面可能是中国毛虾个体自身的生长使种群中大型个体增多,而较小型的中国毛虾个体抵御低温能力较差有关;另一方面也与寒潮降温引起的藻类大量繁殖为中国毛虾提供了大量饵料有关,从而使其能够较为快速地生长成为较大个体。
